Les processus de décomposition de la matière organique occupent une place centrale dans le fonctionnement des écosystèmes. Ils dépendent de nombreux acteurs physico-chimiques et biologiques, et sont impliqués dans plusieurs services écosystémiques. La dynamique de la décomposition et le devenir des produits de la décomposition dépendent fortement des conditions environnementales. Cette dynamique change donc avec les perturbations climatiques et environnementales en cours.
Tous les organismes vivants maintiennent leur intégrité corporelle et leur fonctionnement métabolique en consommant de l’énergie et de la matière prélevées dans leur environnement, réduisant ainsi leur entropie. À leur mort, les molécules composant leur structure ne sont plus construites, remplacées ou réparées. À partir de là, la destruction de ces molécules et des cellules qu’elles constituent est thermodynamiquement favorisée : c’est le processus de décomposition.
Dans cet article nous traiterons principalement de la décomposition de la matière organique en milieu terrestre, en lien avec le sol. La décomposition de la matière en milieu aquatique est également un processus d’une importance majeure pour les écosystèmes et à l’échelle de la planète. La décomposition en eau douce (lacs, rivières) présente de nombreuses similitudes avec la décomposition en milieu terrestre (acteurs, dynamique et contrôles similaires).
Décomposition, décomposeurs, détritivores
La décomposition correspond à l’ensemble des processus physiques, chimiques et biologiques menant à la fragmentation, l’altération et la minéralisation des particules et des molécules de la matière organique morte. Les organismes qui réalisent cette décomposition, et qui dépendent donc trophiquement de la matière organique morte, sont appelés décomposeurs (sens large). Ceux-ci réunissent les détritivores et les décomposeurs microbiens (décomposeurs sens strict). Les détritivores sont des animaux consommant de la matière organique morte et/ou des microorganismes associés à la matière organique morte. Ces éléments sont ingérés, puis digérés dans le tube digestif. Les décomposeurs microbiens sont les microorganismes (bactérie ou champignons) qui consomment la matière organique morte en absorbant des molécules libres parfois issues de la déstructuration des polymères in situ, notamment par des exoenzymes.
Dans la suite du texte, le mot « décomposeur », employé seul, désigne les décomposeurs au sens large, tandis que l’expression « décomposeurs microbiens » est utilisée lorsque seuls les micro-organismes sont impliqués dans le processus décrit.
Les acteurs de la décomposition
Dans les écosystèmes terrestres, la grande majorité de la matière organique primaire (produite par les organismes photosynthétiques) n’est pas consommée directement. En effet, le réseau herbivores-carnivores ne consomme qu’une minorité de cette matière organique vivante. La majorité, jusqu’à 90 %, rejoint le sol après sa sénescence, soit sous forme de feuilles mortes ou branches constituant la litière, soit sous forme de racines mortes ou d’exsudats racinaires, déjà présents dans le sol 1. Ces derniers représentent près de la moitié de la matière organique morte dans les écosystèmes terrestres, mais la décomposition de cette matière souterraine est beaucoup moins étudiée 2. Les processus de décomposition de cette matière organique commencent alors sous l’impulsion de plusieurs acteurs.
Les processus physico-chimiques
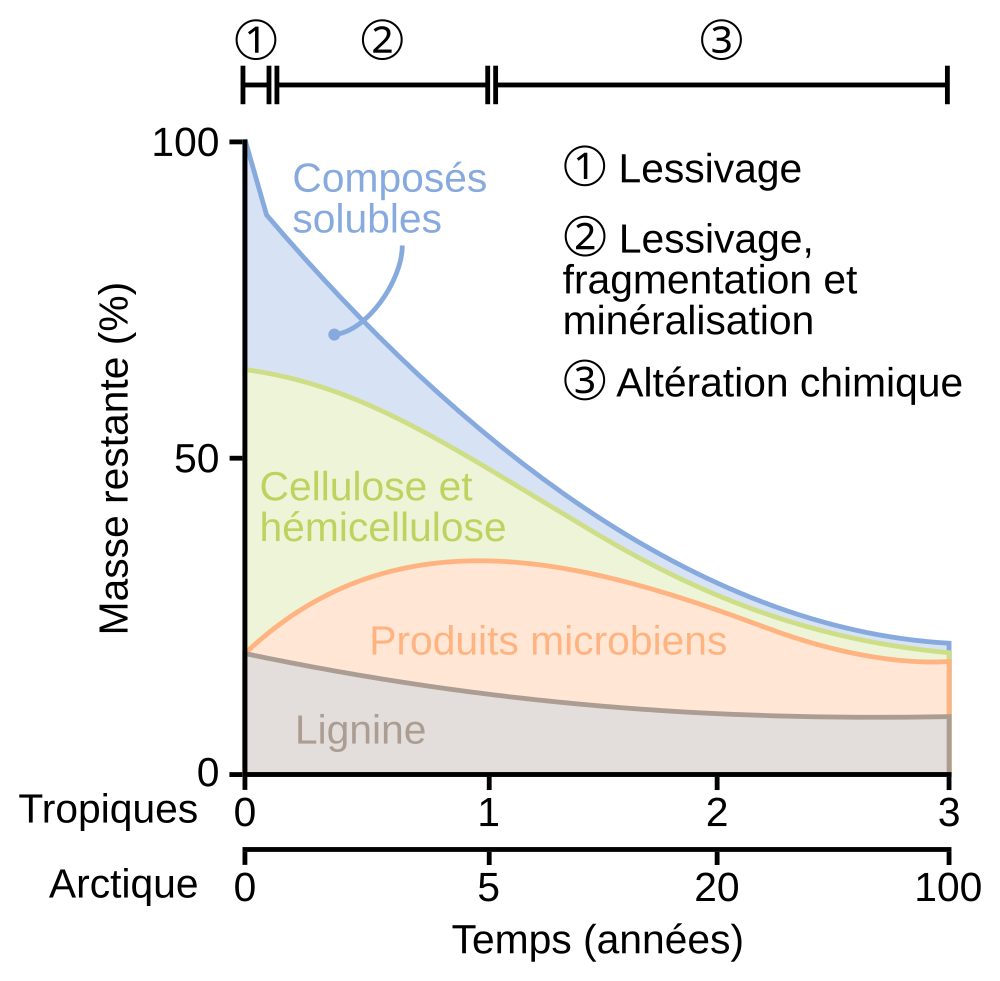
Trois phases ont été identifiées par les auteurs : une phase de lessivage, une phase de fragmentation et minéralisation par les décomposeurs, et une phase d’altération chimique des molécules dans le sol. Les composés solubles disparaissent très rapidement, notamment pendant la phase de lessivage, contrairement à la lignine qui se dégrade peu. Des repères temporels indicatifs sont donnés en contexte tropical (décomposition rapide) et arctique (décomposition lente). Figure d'après de Chapin et coll., 2002 1.
L’eau traverse la matière organique et emporte avec elle les molécules hydrophiles conduisant à leur lessivage (Figure 1). Ce lessivage par l’eau peut mener à la perte de jusqu’à un tiers de la masse des feuilles mortes 1. Par ailleurs, le cycle de gel-dégel provoque l’éclatement des cellules, car l’eau prend plus de volume à l’état solide qu’à l’état liquide. Il en va de même pour les cycles d’humectation et de dessèchement, qui exercent également des stress physiques sur les structures cellulaires. De plus, les rayonnements ultraviolets (UV) de la lumière agissent sur certaines molécules, comme la lignine, et les dégradent partiellement. Ces processus contribuent à fragmenter, déstructurer et altérer la matière organique aussi bien à l’échelle de l’organe (ex : feuille morte) qu’à l’échelle moléculaire.
Les microorganismes
La matière organique morte ne possède plus de système de défense actif contre les autres organismes et est donc facilement colonisable par les microorganismes qui s’en nourrissent. Ce sont principalement les bactéries et les champignons qui participent à la dégradation enzymatique des cellules et des matrices extracellulaires.
Ces organismes sont en effet capables de libérer des enzymes dans le milieu extracellulaire, notamment des oxydases, capables de décomposer les molécules de la matière organique morte. En particulier, seuls certains champignons basidiomycètes, du groupe des Agaricomycètes (par exemple le polypore du pin, Heterobasidion annosum), sont capables de produire des peroxydases et des radicaux libres suffisamment puissants pour décomposer la lignine 12, molécule particulièrement résistante des matrices extracellulaires du bois. Ces champignons sont à l’origine de la pourriture blanche, où la structure extracellulaire du bois est complètement perdue, résultant en une dégradation du bois en fibres blanchâtres. À l’inverse, les champignons à l’origine de la pourriture dite brune sont uniquement capables de dégrader la cellulose et l’hémicellulose, conservant ainsi une partie de la structure extracellulaire du bois, et résultant en une dégradation du bois en blocs bruns (Figure 2) 34.

Pourriture brune (en haut, décomposition de la cellulose uniquement, structure en blocs bruns) et pourriture blanche (en bas, décomposition de la cellulose et de la lignine, structure fibreuse blanchâtre) sur du bois provenant vraisemblablement du même arbre. Octobre 2023, parc naturel régional du Pilat, au sud de Saint-Étienne (1000 m d’altitude).
En déstructurant et en assimilant des molécules de la matière organique morte, les microorganismes décomposeurs participent à modifier les propriétés de la litière, tant d’un point de vue physique, en diminuant la résistance des éléments qui composent la litière, que chimique, car la minéralisation du carbone organique en CO2 diminue le rapport carbone/nutriments (par exemple, le rapport C/N, c’est-à-dire carbone/azote). L’ensemble des transformations de la litière par des processus physico-chimiques et microbiologiques, qui ont pour effet de la rendre plus appétente et assimilable par les détritivores, est appelé conditionnement de la litière.
Les détritivores
Les détritivores sont des animaux participant à la décomposition que ce soit par la fragmentation, l’ingestion, l’assimilation et la minéralisation de tout ou partie de la matière organique morte, ou par la consommation des décomposeurs microbiens. Par leurs différentes actions, ils accélèrent la vitesse de décomposition de 30 à 50 % (Figure 3) 12. Certains détritivores (ex : collemboles, acariens oribates) sont spécialisés dans la consommation des biofilms et mycéliums des décomposeurs (Figure 5). Ils sont qualifiés de racleurs ou de brouteurs de biofilms et mycéliums. D’autres, tels que les cloportes, les termites, les gastéropodes, les diplopodes (myriapodes) et les vers de terre, fragmentent la litière et ingèrent des fragments de litière et de microorganismes décomposeurs. Ils sont qualifiés de fragmenteurs.
L’ingestion de matière organique morte conduit à la minéralisation des molécules (sous forme de CO2 pour le carbone, d’ammoniaque NH3 pour l’azote…) ou de transformation des molécules dans les fèces. En effet, un élément central de leur rôle dans le processus de décomposition est la production de boulettes fécales. Ce sont des structures organo-minérales compactes, issues de la fragmentation de la litière en particules par l’appareil buccal et le reste du système digestif des détritivores. Ces boulettes présentent plusieurs caractéristiques qui favorisent leur décomposition microbienne : rapport C/N faible, forte teneur en eau 3, richesse en carbone et azote dissous 456, et peu de tanins 78. Elles peuvent aussi, dans d’autres cas, ralentir ce processus en augmentant la proportion de composés récalcitrants, notamment la lignine 9. Le devenir de la matière ingérée dépend donc de la dégradation physique et chimique propre à chaque détritivore (Encart 2).
Certains détritivores agissent aussi par bioturbation, en déplaçant la matière organique dans le sol. À ce titre, les vers de terre exercent un effet particulièrement important sur la litière, en fragmentant, ingérant, et enfouissant de grandes quantités de matière organique. La fragmentation de la litière peut favoriser la décomposition microbienne en augmentant les surfaces d’attaque disponibles aux bactéries et aux champignons.

À l’équateur (latitude 0), la faune est responsable d’un peu moins de 50 % de la décomposition. À haute latitude (proche des pôles), plus de 80 % de la décomposition est réalisée par les microorganismes. La vitesse de décomposition ainsi que l’importance relative des microorganismes et des détritivores dans la décomposition peuvent être étudiées grâce à la méthodologie des sacs de litière. Traduit et adapté de Zeng et coll., 2024 1.
La dégradation chimique chez les détritivores
En plus de la dégradation physique évoquée dans la partie 1.3, les détritivores contribuent à la décomposition par une dégradation chimique de la matière organique. Cette dégradation chimique est dépendante des enzymes synthétisées par le détritivore lui-même, et par ses endosymbiotes microbiens intestinaux 1. En effet, certains taxons comme les vers de terre, les isopodes, les diplopodes et les termites coproduisent des cellulases extracellulaires avec leurs endosymbiotes intestinaux 234. De plus, des études récentes ont montré que certains taxons comme les isopodes et gastéropodes sont capables de produire leurs propres cellulases et hémicellulases, indépendamment de leurs endosymbiotes 5.
Place dans les sols
Cycle du carbone
Le dioxyde de carbone (CO2) est un composé minéral présent à 0,04 % dans l’atmosphère terrestre. Les émissions anthropiques de ce gaz sont responsables de la majeure partie du réchauffement climatique. Ce gaz est prélevé par les végétaux via la photosynthèse et constitue la matière première de la production de biomasse, la matière organique. À sa sénescence, la décomposition de la matière organique, via le métabolisme des décomposeurs, entraîne un retour du CO2 vers l’atmosphère via la respiration. Cependant, le carbone n’est pas entièrement minéralisé, et une fraction finit donc par se fossiliser (Figure 4).

Les cercles jaunes illustrent les acteurs directs (microorganismes et détritivores) et indirects (prédateurs) de la décomposition. Les flèches jaunes entre les cercles représentent la consommation de la litière et de la matière organique morte souterraine par les microorganismes et les détritivores, la consommation des microorganismes par les détritivores, ainsi que celle des détritivores par les prédateurs. Les flèches grises représentent les processus de catabolisme liés à la minéralisation du carbone via la respiration, et à la minéralisation des nutriments. La flèche blanche représente le processus d’anabolisme via la photosynthèse. Les flèches en pointillés représentent le chemin de la matière inorganique et organique (MO).
La quantité de carbone organique stockée dans les sols varie selon leur nature géologique, les conditions climatiques locales et les pratiques de gestion des terres. Par exemple, les sols des régions froides de l’hémisphère Nord renferment la plus grande quantité de carbone stocké, en raison des basses températures qui freinent l’activité biologique et ralentissent ainsi la décomposition de la matière organique 1. Or, le réchauffement climatique perturbe cet équilibre en accélérant la décomposition, entraînant la minéralisation et donc l’émission de ce carbone dans l’atmosphère, sous forme de CO2 ou de CH4.

Cycle des nutriments
Les processus de décomposition participent au cycle des nutriments (Figure 4). Les végétaux prélèvent la majorité des nutriments sous forme minérale, dans le sol (ammonium NH4+ et nitrate NO3– pour l’azote ; phosphate PO43– pour le phosphore, cation K+ pour le potassium…). Certains organismes dits autotrophes à l’azote peuvent prélever le diazote N2 de l’atmosphère et le convertir en ions ammonium (bactéries en symbiose ou non avec des plantes 1). Ces éléments participent à la production de matière organique (ex : protéines et acides nucléiques pour l’azote, acides nucléiques et phospholipides pour le phosphore…), et au fonctionnement cellulaire (ex : transduction du signal pour le phosphore) 2. Lors de la décomposition, ces éléments sont directement ou indirectement (via la consommation par les décomposeurs) reminéralisés et rejetés dans l’environnement (dans l’eau ou le sol) 3.
Une synthèse de 2005 d’Emma Sayer 4, vulgarisée en français sur le site Zoom Nature, a illustré l’impact de la décomposition sur la richesse en nutriments du sol. Le ratissage de litière, une pratique paysanne particulièrement utilisée au XIXe siècle, consiste à récupérer les feuilles mortes au sol (la litière) dans une forêt et à l’utiliser pour le bétail. En comparant des forêts où cette pratique du ratissage de litière a eu lieu pendant plusieurs années avec des forêts semblables où cette pratique n’a pas eu lieu, plusieurs études ont montré que le sol des forêts avec ratissage de litière contenait moins de nutriment (N, P, K, Ca, Mg). Avec l’export des feuilles mortes, les nutriments contenus dans celles-ci étaient également exportés, et donc perdus pour la forêt. Le sol était d’autant plus appauvri en nutriments que cette pratique de ratissage de litière avait eu lieu pendant de nombreuses années. Cette baisse de nutriments peut même conduire à un ralentissement de la croissance des arbres, et donc à des arbres plus petits dans les forêts où le ratissage de litière a été pratiqué durant plusieurs décennies.
Le même processus explique l’appauvrissement en nutriments des sols agricoles : l’exportation de tout ou partie de la matière végétale produite appauvrit d’année en année le sol en nutriments. Seul un apport en engrais naturels ou chimiques, ou une rotation des cultures (en particulier avec des fabacées fixatrices d’azotes) permet de réenrichir le sol 5.
Flux d’énergie et de matière au sein du réseau trophique brun
En plus de constituer un abri (sous forme de microhabitats), la matière organique en décomposition constitue une ressource trophique pour un grand nombre d’espèces microbiennes et animales. Ces organismes sont eux-mêmes consommés par d’autres organismes, constituant ainsi un réseau trophique basé sur la matière organique morte, parfois appelé réseau « brun », par opposition au réseau trophique « vert » basé sur la consommation d’organismes photosynthétiques vivants. Il est possible de reconstituer un réseau trophique, c’est-à-dire d’identifier les liens entre organismes consommateurs et consommés, en se basant sur plusieurs méthodes (Figure 5) :
-
L’observation directe au champ ou au laboratoire : Les observations in situ (dans la nature) ou au laboratoire de comportements de consommation permettent d’identifier le régime alimentaire de différents organismes. Il est également possible d’observer dans les contenus stomacaux ou dans les fèces les restes de la ressource et de les identifier (restes d’animaux ou restes de végétaux). Cependant, il peut être difficile d’identifier précisément l’espèce consommée.
-
L’observation d’indices morphologiques, comme les pièces buccales, peuvent permettre de déterminer si l’organisme consomme de la sève, des mycorhizes, ou des animaux par exemple.
-
Les mesures d’abondance d’espèces : L’abondance des espèces au sein d’un réseau trophique covarie dans le temps et dans l’espace. Mesurer l’abondance des espèces dans un écosystème en faisant des relevés de terrain peut être utile pour la construction de réseaux trophiques en reconstituant des dynamiques consommateurs-ressources (type prédateurs-proies).
-
La mesure en teneur d’isotope stable : Les consommateurs (de niveau trophique élevé) ont tendance à s’enrichir en isotope lourd 15N de l’azote par rapport à la ressource qu’ils consomment (de niveau trophique inférieur). Des prédateurs ont donc tendance à avoir une composition en isotope lourd 15N de l’azote plus élevé que les herbivores 6. En pratique, l’enrichissement en azote lourd d’un échantillon est quantifié par son δ15N, dans lequel le rapport isotopique 15N/14N de l’échantillon est comparé à une référence, le 15N/14N de l’atmosphère. $$\delta^{15}\mathrm{N} = \left( \frac{ \left(\frac{^{15}\mathrm{N}}{^{14}\mathrm{N}}\right)_{\text{échantillon}} }{ \left(\frac{^{15}\mathrm{N}}{^{14}\mathrm{N}}\right)_{\text{atmosphère}} } - 1 \right) \times 1000$$
-
La production ou assimilation de certains composés biochimiques : Les enzymes digestives produites peuvent également être informatives. Par exemple, la production de chitinase peut indiquer une capacité à digérer des champignons et/ou des arthropodes 7. La composition en acides gras peut également être un indicateur de la ressource trophique assimilée. Par exemple, l’acide gras 18:2ω6,9 est caractéristique des champignons. Le retrouver chez un animal indique que cet animal consomme des champignons 8.
-
Test ELISA : Cette méthode se base sur le marquage des proies par des anticorps monoclonaux et leurs détections dans les contenus stomacaux des consommateurs à l’aide de la technologie ELISA (méthode immuno-enzymatique) afin d’identifier les proies ingérées 9.
-
Le séquençage ADN du contenu stomacal : L’association du séquençage à haut débit (NGS) et du métabarcoding ADN permet d’analyser le contenu digestif sans connaissance préalable des espèces consommées et de reconstituer des réseaux trophiques complexes.
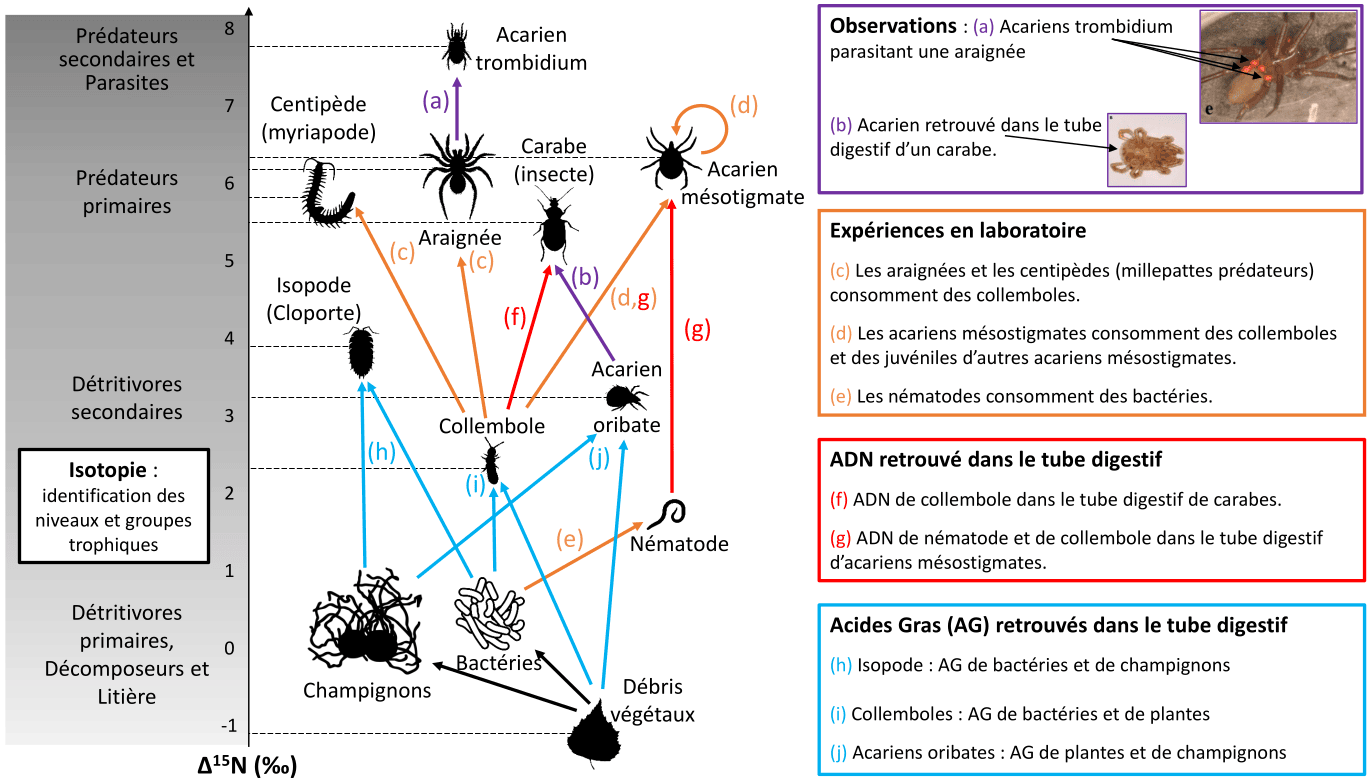
L’isotopie, en particulier le $\delta^{15}\mathrm{N} = \left( \frac{ \left(\frac{^{15}\mathrm{N}}{^{14}\mathrm{N}}\right)_{\text{échantillon}} }{ \left(\frac{^{15}\mathrm{N}}{^{14}\mathrm{N}}\right)_{\text{atmosphère}} } - 1 \right) \times 1000$, permet d’identifier le niveau trophique des organismes (traits horizontaux en pointillés noirs). D’autres méthodes permettent d’identifier des liens trophiques (en violet, orange, rouge, et bleu) entre différents acteurs du réseau. Certains acteurs peuvent consommer des congénères (flèche circulaire (d) sur l’acarien prédateur). Certains groupes taxonomiques (ex : acariens) peuvent avoir des rôles trophiques très différents (parasites, prédateurs, détritivores). Les liens présentés ici sont binaires (il y a une interaction, ou pas), mais une interaction trophique peut avoir différentes intensités (intensité forte : le consommateur dépend exclusivement d’une seule ressource ; intensité faible : le consommateur consomme occasionnellement voire rarement cette ressource, ou en petites quantités). Enfin, une absence de lien ne signifie pas qu’il n’y a pas d’interaction ; il se peut simplement qu’elle n’ait pas été observée. C’est l’accumulation de différents indices avec différentes méthodes (ex : flèche (d, g)) et différents niveaux de preuves (l’isotopie ne suffit pas à relier deux acteurs du réseau), issus de différentes études, qui permet de reconstituer des réseaux de plus en plus robustes et fiables. Les illustrations noires proviennent de phylopic.org. Références : flèche (a) 1, flèche (b) 2, flèche (c) 3, flèche (d) 4, flèche (e) 5, flèche (f) 6, flèche (g) 7, flèches (h, i, j) 8, flèches noires 910.
Cependant, la reconstruction du réseau brun est rendue difficile pour plusieurs raisons. Premièrement, du fait de la structure même du réseau détritique : tous les organismes, une fois morts, ou leurs déchets (fèces), retournent à la base du réseau brun, ce qui complexifie notamment les réseaux d’enrichissement en isotope lourd de l’azote (15N) 1. De plus, de nombreux détritivores ont un régime alimentaire généraliste. Par exemple, les isopodes peuvent consommer des fragments végétaux, des biofilms microbiens, des mycorhizes, des algues, des fèces (dont leurs propres fèces ; on dit qu’ils sont coprophages), et même des congénères morts ou vivants (cannibalisme) 2. Certains détritivores peuvent également ingérer des débris végétaux et des mycorhizes, mais n’assimiler que les mycorhizes : les débris végétaux ne sont pas assimilés et sont alors rejetés sous forme de fèces. Il ne faut donc pas les considérer comme une ressource trophique. Les acteurs du réseau brun ont des tailles et des métabolismes très différents (bactéries, arthropodes, nématodes…) et différentes méthodes sont donc nécessaires pour étudier ces différents organismes (Figure 5). Enfin, le réseau brun peut être en interaction avec le réseau trophique « vert », basé sur la consommation de matière vivante des producteurs primaires, car certains prédateurs de détritivores consomment également des herbivores (ex : araignées consommant des collemboles ou des pucerons).
Paramètres contrôlant la décomposition
Paramètres abiotiques
Les paramètres abiotiques contrôlent la vitesse et le devenir de la décomposition. La température accélère et modifie l’équilibre des réactions chimiques. Elle accélère généralement les réactions biochimiques et l’activité biologique jusqu’à un effet seuil 3. L’humidité joue également un rôle important : dans les endroits secs (déserts chauds et déserts froids), le lessivage des molécules et l’activité biologique sont fortement réduits. La quantité de dioxygène est également très importante pour l’activité biologique : dans les tourbières, le sol est saturé en eau stagnante pauvre en dioxygène (on parle d’un milieu anoxique). Il en résulte une décomposition réduite. De plus, la luminosité et en particulier les UV, sont responsables de la photodégradation plus intense en milieu ouvert (prairie) qu’à l’ombre des forêts. Enfin, le pH du sol est un facteur important de régulation de la décomposition (Figure 6), en modifiant à la fois les réactions chimiques et l’activité biologique. Les sols acides ont généralement une décomposition lente 4.

Cette figure représente un continuum entre différentes caractéristiques des humus mull, moder et mor et le lien à différents paramètres du sol. L’humus correspond à l’ensemble de matières organiques incorporées au sol, issues des processus de décomposition et de transformation de la matière organique morte. Un humus de type mull est caractérisé par une décomposition rapide de la matière organique, une intense activité biologique et un brassage en profondeur. Un humus de type mor est caractérisé par une décomposition lente, avec une matière organique s’accumulant en surface, et une activité biologique faible. Un humus de type mull est traditionnellement associé à un sol carbonaté plutôt que siliceux, avec de l’argile plutôt que du sable, des cations basiques (pH neutre ou alcalin) plutôt que des protons (sol acide), beaucoup de nutriments disponibles, de l’azote inorganique (nitrates NO3− et ammonium NH4+), des vers de terres fouisseurs et des macroinvertébrés, des arbres associés à des mycorhizes arbusculaires (MA) plutôt qu’à des champignons ectomycorhiziens (CEM), un sol dominé par les bactéries plutôt que par les champignons, avec une matière organique labile (facilement décomposable ; ex : feuillus) plutôt que récalcitrante (ex : aiguilles de pin), une décomposition microbienne dynamique plutôt qu’une décomposition dominée par la dégradation de lignine en phénols, une matière organique associée aux minéraux (MOAM, < 50 μm) plutôt qu’une matière organique particulaire (MOP, entre 50 μm et 2 mm), et un sol caractérisé par des agrégats (indicateurs d’un sol fertile) plutôt que par des complexes entre matière organique et aluminium ou fer. Figure traduite de Prescott & Vesterdal, 2021 1, conçue par Julia Amerongen-Maddison.
L’influence de ces paramètres abiotiques se retrouve à l’échelle planétaire, notamment avec la latitude 1. Les zones à haute latitude, de climat froid, présentent généralement une décomposition lente, à l’image des forêts boréales qui contribuent à stocker du carbone dans le sol. À l’inverse, la décomposition est généralement rapide dans les forêts tropicales, et participe à recycler rapidement la matière (ions minéraux, carbone…).

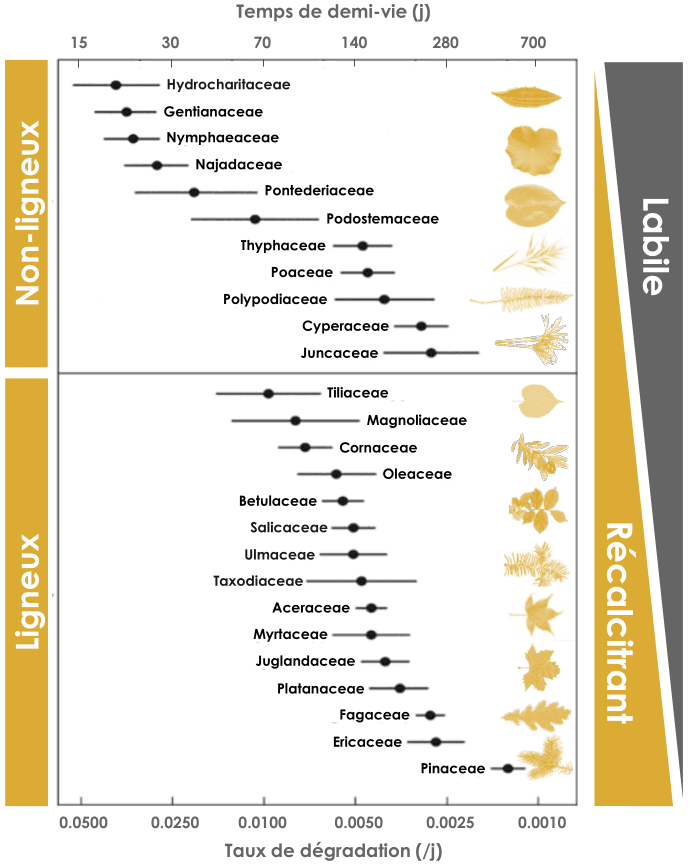
La masse des feuilles au cours de la décomposition suit une exponentielle décroissante et les vitesses de décomposition peuvent donc être indiquées en taux de décomposition (/j) ou en temps de demi-vie. Ces taux de décomposition ont été déterminés en milieu aquatique (rivières, lacs…). Les vitesses de décomposition sont plus lentes d’environ un ordre de grandeur en milieu terrestre 1. On observe des différences de vitesse de décomposition entre plantes ligneuses et non-ligneuses, suivant un gradient de matériaux labiles (dégradation facile et rapide) et récalcitrants (dégradation lente). Ces différences sont liées à des différences de composition physique (dureté) et chimique (proportion d’éléments solubles, d’azote et de phosphore, de lignine, etc.). Figure adaptée de Webster & Benfield 1986 2.
Nature de la litière
La nature même de la matière organique est un facteur important de contrôle de la décomposition. Les matières animales peuvent se décomposer très rapidement à l’inverse des matières végétales. Au sein des matières végétales, les feuilles de plantes non ligneuses se décomposent généralement plus rapidement que les feuilles de plantes ligneuses. Les feuilles (aiguilles) de conifères sont particulièrement lentes à se décomposer (Figure 7). Ces différences de vitesse de décomposition tiennent à la nature de la matière. En particulier, une litière riche en éléments réfractaires telles que les lignines et celluloses, ce qui est le cas pour les aiguilles de pin, se décompose plus lentement, car ces molécules sont plus résistantes à la dégradation et confèrent à la litière une résistance physique qui limite sa consommation par les détritivores 1. Une litière riche en nutriments (azote, phosphore) se décompose généralement plus rapidement. Enfin, certains éléments tels que les phénols et tanins réduisent la digestibilité ou l’appétence de la matière par les détritivores, réduisant ainsi la vitesse de décomposition et modifiant son devenir.
Interactions biotiques
Les interactions biotiques au sein du réseau trophique brun influencent l’abondance, la diversité, et l’activité des acteurs de la décomposition et participent donc à la régulation de la décomposition. Les microorganismes décomposeurs modifient la litière, en la conditionnant (réduction de la résistance physique, augmentation de la teneur relative en nutriments, et donc diminution des rapports C/N et C/P), ce qui la rend plus appétente et davantage consommée par les détritivores. Différentes espèces de détritivores peuvent présenter des appétences différentes selon le degré de transformation de la litière par les microorganismes 1. Cette variabilité d’appétence suggère une possible complémentarité temporelle entre les détritivores. Cette complémentarité peut également être observée entre gastéropodes et diplopodes : en raclant la litière, les gastéropodes fragilisent la litière et facilitent son accès, rendant la matière organique plus aisément consommable par les diplopodes 2. De plus, en fragmentant la litière, les macrodétritivores augmentent les surfaces d’attaque disponibles par les microorganismes. Les détritivores augmentent généralement la vitesse de décomposition (Figure 3). Les règles et les conséquences des mécanismes de facilitation et de compétition entre décomposeurs sont encore très mal connues. De même, les prédateurs peuvent diminuer la vitesse de décomposition en consommant les détritivores ou en réduisant la consommation de litière par ces derniers. En effet, en présence d’un prédateur, certains détritivores passent davantage de temps à se cacher, et consomment moins de litière 3. Enfin, des organismes symbiotes ou parasites peuvent modifier les dynamiques des populations de décomposeurs et ainsi modifier la vitesse et le devenir de la décomposition.
Perturbations de la décomposition et des services écosystémiques
Le processus de décomposition est associé à différents services écosystémiques (fertilité des sols et rivières, stockage de carbone, support de biodiversité, élimination de la litière…). Nous proposons ici deux exemples de perturbation du réseau trophique brun par des espèces nouvellement introduites. Ces perturbations du réseau trophique ont mené à des perturbations de la décomposition et du fonctionnement général des écosystèmes concernés.
Des bousiers en Australie
À partir du XVIIIe siècle, les colons européens introduisirent des bovins en Australie. Au XIXe siècle, l’Australie fit face à un problème majeur : les bouses produites par des millions de bovins ne se décomposaient pas : elles polluaient les champs dans lesquels les animaux ne voulaient plus se nourrir, et séquestraient des nutriments qui ne n’étaient pas minéralisés, ce qui limitait la production végétale. Les bouses étaient tout de même exploitées par des mouches dont les populations explosèrent, répandant des maladies 4. Il existe des bousiers autochtones en Australie, mais ils sont habitués aux déjections solides des marsupiaux, et aucune espèce n’est adaptée à la bouse de bovin. À partir de 1965, des programmes d’introduction de dizaines d’espèces de bousiers européens et africains ont été lancés pour résoudre le problème 5. Le principal projet d’introduction a duré jusqu’en 1985 et a globalement été un succès, même si des programmes d’introduction sont toujours actifs aujourd’hui.
Un chiffre montre l’importance des bousiers dans le fonctionnement des écosystèmes : le service écosystémique de décomposition fourni par ces insectes a été estimé, pour le seul territoire de la Grande-Bretagne, à 367 millions de livres par an 6.
Les lombrics invasifs en Amérique du Nord
La présence d’espèces exotiques envahissantes à un endroit donné peut fortement perturber le réseau brun local et les processus de décomposition associés. Les forêts d’Amérique du Nord ne possèdent pas de vers de terre natifs. Or, des vers européens et asiatiques sont invasifs dans ces milieux. Du fait de leur impact sur le sol, les vers de terre perturbent fortement les sols nord-américains et leur fonctionnement. Ils accélèrent drastiquement la décomposition, perturbent la disponibilité en nutriments, modifient la structure du sol par la bioturbation et la production de turricules. Ils peuvent également perturber les communautés locales de microorganismes, d’arthropodes, et mêmes les communautés de plantes 7. Pour l’instant, l’impact de ces changements sur le cycle du carbone est inconnu. L’enjeu est grand car les sols des forêts nord-américaines stockent beaucoup de carbone.