

Notre étude récemment publiée dans Nature présente une estimation du taux de mutations germinales chez 68 espèces de Vertébrés. En utilisant la méthode directe de comparaison de génomes entre parents et enfants, nous avons pu mettre en évidence un biais paternel dans la contribution aux mutations transmises aux descendants. Nous avons aussi pu déterminer certains des traits biologiques qui agissent sur l’évolution du taux de mutation. Cela nous a permis de conclure que le taux de mutation est un trait soumis à la sélection naturelle.
Les mutations germinales sont la source de l’évolution. Ces mutations germinales, en opposition aux mutations somatiques, touchent les gamètes et seront donc transmises aux descendants. Elles peuvent être à l’origine de maladies génétiques, mais aussi d’adaptations évolutives. Quantifier le taux auquel les mutations germinales sont introduites dans les génomes de différentes espèces est important pour comprendre l’apparition de nouvelles maladies génétiques, appréhender les événements démographiques importants pour la conservation, ou encore dater les phénomènes de spéciations. Plusieurs hypothèses ont été formulées pour tenter d’expliquer l’évolution de ces taux de mutations, mais le manque d’estimations précises et comparables reste un frein pour tester ces hypothèses 1.
Grâce au séquençage de nouvelle génération, une approche directe d’estimation de taux de mutations a émergé. Cette méthode repose sur une comparaison du génome de deux parents avec celui de leur enfant. En pratique, cela revient à chercher quelques aiguilles (les mutations germinales par dizaines) dans une énorme botte de foin (des génomes de plus de 3 milliards de bases). Ainsi, différentes méthodes ont été utilisées dans les études actuelles, qui se focalisent généralement sur une unique espèce. Il est donc difficile de comparer les taux estimés sans introduction d’un biais méthodologique 2.
Dans notre étude, nous voulions couvrir une grande partie de la phylogénie des Vertébrés, tout en étant limités par la disponibilité des génomes de références et par les informations sur les pédigrées des espèces 3. Nous avons collecté 151 trios (deux parents et un enfant) pour 68 espèces de Vertébrés : 36 Mammifères, 18 Oiseaux, 5 Lépidosauriens (groupe comprenant les lézards, serpents, iguanes…), une espèce de tortue, et 8 Actinoptérygiens (poissons à nageoires rayonnées). Pour 39 espèces, seul un unique trio a pu être analysé. Néanmoins l’analyse des espèces pour lesquelles nous disposions de plusieurs trios, ainsi que de précédentes études se focalisant sur une seule espèce avec de nombreux trios, montrent une faible variabilité intraspécifique du taux de mutations germinales. Nous avons estimé un taux de mutations en utilisant la même méthode d’analyse sur chaque espèce, ce qui nous a permis de faire une comparaison systématique des taux entre espèces. Nous avons pu explorer certains mécanismes moléculaires concernant l’apparition de ces mutations et tester les hypothèses sur l’évolution du taux de mutations.
Notre première observation a été une variation du taux de mutation par génération d’un facteur 40. Néanmoins, la plupart des espèces étudiées présentent des taux de mutation compris dans une gamme restreinte de valeurs, allant de 0,5 à 1,5.10−8 mutations par site et par génération. Une variation modérée compte tenu de la variation de traits biologiques entre espèces. Par exemple, le poids d’une orque et celui d’un combattant varient d’un facteur de 2,8 millions et il y a un facteur 93 entre le temps de génération d’un humain et celui d’un gecko. Le taux de mutation par génération serait donc contraint par des forces évolutives.
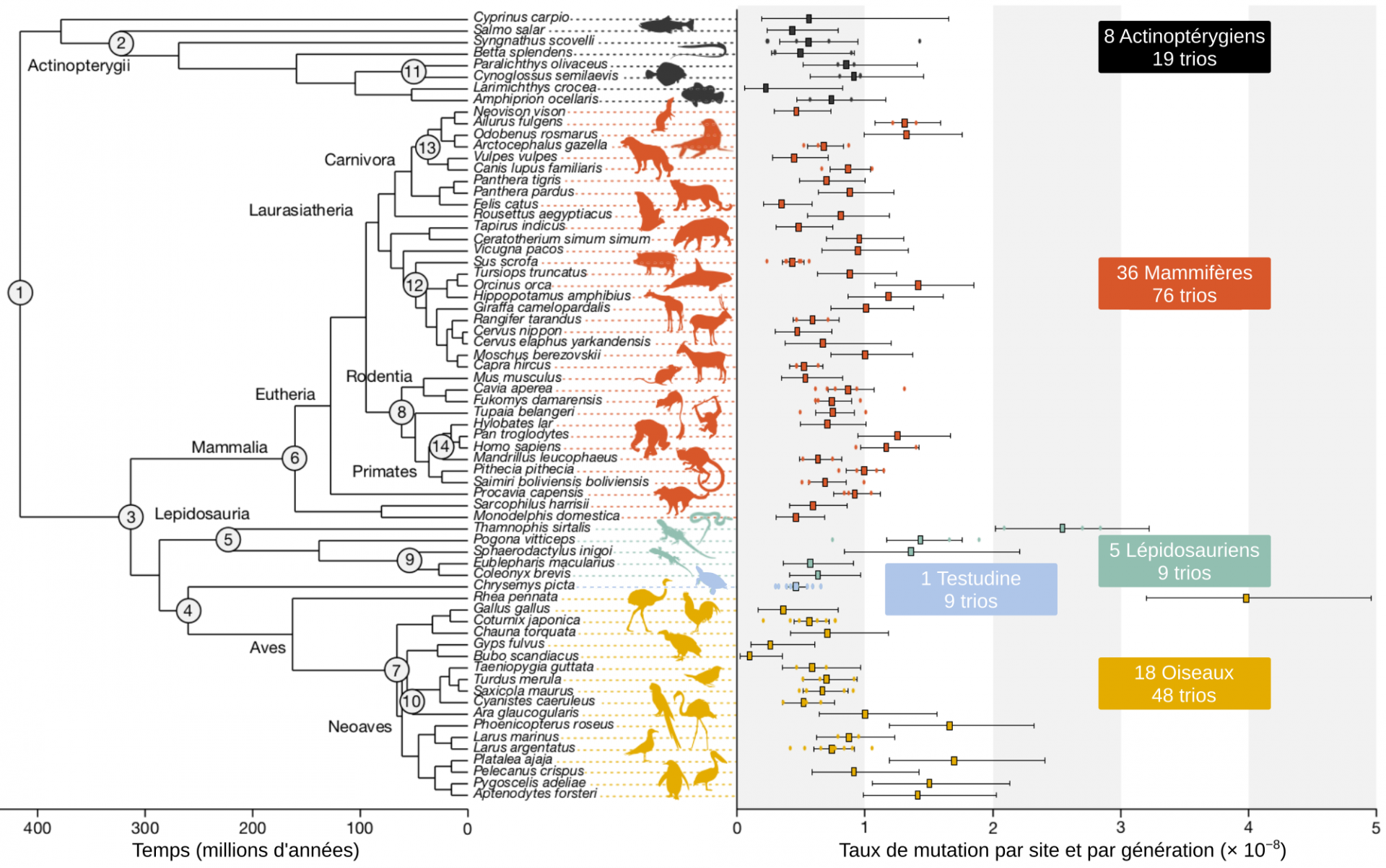
Pour chaque espèce étudiée, les cercles colorés correspondent aux taux de mutations calculés sur les différents trios analysés tandis que les carrés représentent la moyenne et les lignes noires horizontales aux intervalles de confiance binomiaux à 95%). Le taux de mutation montre une variation d’un facteur de 40 entre les 68 espèces. Les silhouettes d’animaux proviennent de PhyloPic, sauf pour les silhouettes de Syngnathus scovelli créée par Josefin Stiller, de Sarcophilus harrisii créée par S. Werning, et de Pan troglodytes créée par T. M. Keesey (vectorisation) et T. Hisgett (photographie); ces deux dernières sont sous la licence CC-BY 3.0.
Notre méthode nous a permis d’observer un biais paternel chez les Mammifères et les Oiseaux : les pères contribuent à une plus grande partie des mutations que les mères. Ce biais peut s’expliquer par la différence de gamétogenèse entre les deux sexes. Chez ces espèces, les mâles produisent des spermatozoïdes toute leur vie (avec réplication d’ADN) entrainant une accumulation de mutations. À l’inverse, chez les femelles, les ovocytes I sont formés durant le développement embryonnaire et il n’y a donc plus de réplication d’ADN après la naissance. Les Lépidosauriens et les Actinoptérygiens présentent moins de différence de gamétogenèse entre les sexes, puisque les mâles produisent des spermatozoïdes seulement en période de reproduction et les femelles produisent de nouveau œufs (avec réplication d’ADN) au cours de leur vie. Or, ces espèces ne présentent pas de biais paternel dans la contribution des mutations germinales.
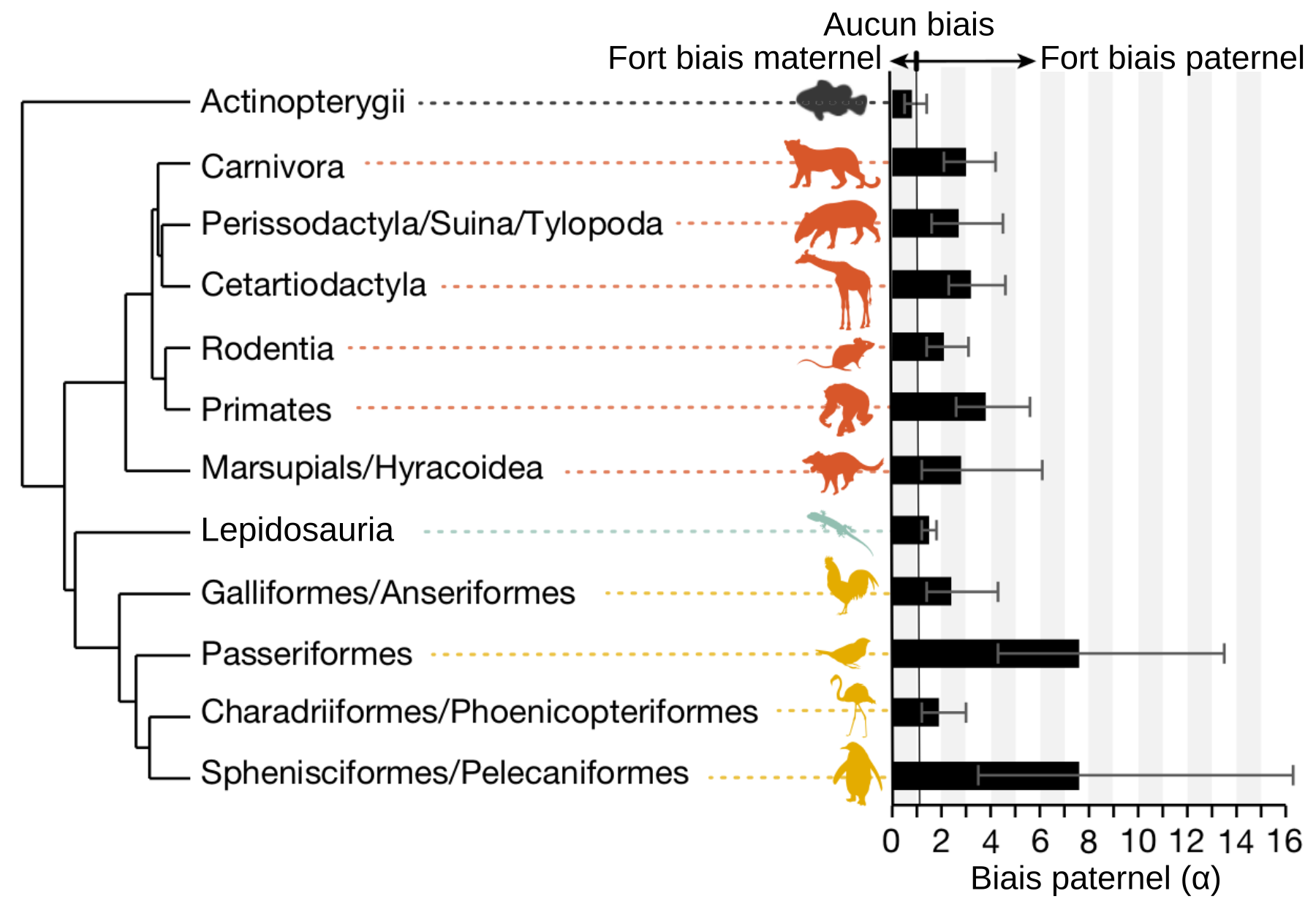
Le ratio mâle/femelle dans la contribution aux mutations transmises a été analysé par groupe d’espèces. Le biais le plus important s’observe chez deux lignées d’Oiseaux, alors que ce biais est négligeable chez les Actinoptérygiens (poissons à nageoires rayonnées) et les Lépidosauriens (lézards, serpents, iguanes…).
Le taux de mutations germinales chez l’espèce humaine
Chez l’espèce humaine, le taux de mutations germinales est de 1,2.10−8 par site et par génération. En considérant une longueur de génome diploïde voisine de 6 milliards de paires de bases (3 milliards de paires de bases par génome haploïde), cela représente 72 mutations par génération, soit 72 différences entre le génome de l’enfant et les deux moitiés de génome transmises par ses parents.
Dans la mesure où le biais paternel est de 3,8, c’est-à-dire que le père transmet 3,8 fois plus de mutations que la mère, cela signifie que sur les 72 mutations, environ 57 viennent du spermatozoïde et seulement 15 de l’ovocyte.
Enfin, nous avons mis en évidence des corrélations entre certains traits biologiques et les taux de mutation. Le temps de génération et l’âge de la maturité sexuelle sont positivement corrélés avec le taux de mutation par génération, alors que le nombre de descendants par génération ou la taille de population effective sont négativement corrélés au taux de mutation. Cette dernière observation va dans le sens d’une hypothèse formulée par Michael Lynch et qui relie la taille de la population aux effets de la sélection naturelle et de la dérive génétique sur la fréquence des mutations 1. Dans une grande population, la sélection naturelle contribuerait à diminuer la fréquence des mutations dans les cellules germinales, puisque la plupart des mutations ont des effets délétères sur la descendance. Alors que dans une petite population, l’effet de la sélection naturelle est négligeable face à la dérive génétique, et un plus haut taux de mutation est observé.
En conclusion, bien que les mutations germinales soit la source ultime de l’évolution, les taux auxquelles ces mutations vont être introduites dans les génomes de différentes espèces sont eux-mêmes soumis à la sélection naturelle, comme n’importe quel autre trait biologique. Nos résultats ont permis de tester des hypothèses sur l’évolution de ces taux de mutations ou sur la contribution des sexes dans les mutations sans validation jusqu’ici par manque de données. Une limite de cette étude reste l’échantillonnage, puisque nous avions seulement un trio (deux parents et un enfant) pour de nombreuses espèces. Obtenir de plus grands pédigrées permettra une meilleure compréhension des variations au sein d’une espèce ainsi que des mécanismes moléculaires sous-jacents à ces mutations. Cette étude est un pas en avant dans l’estimation des taux de mutations et ouvre de nouveaux horizons, notamment pour la calibration des « horloges moléculaires » et l’estimation de la divergence des espèces.





