Une protéine liant les sucres et située à la surface du virus H5N1 de la grippe aviaire.
Depuis son apparition en 1996, le virus H5N1, responsable de la grippe aviaire (également connue sous le nom d’influenza aviaire), a causé la mort et l’abattage de millions d’oiseaux dans le monde entier, entraînant des pertes économiques considérables pour l’industrie de la volaille et suscitant des inquiétudes concernant des populations d’oiseaux sauvages à risque. Les récentes épidémies n’ont pas touché que les oiseaux, mais également les mammifères sauvages et domestiqués.
Un assortiment de sous-types
L’hémagglutinine est une protéine en forme de pointe qui, avec la neuraminidase, recouvre la surface des virus de la grippe. Il existe actuellement 18 sous-types connus d’hémagglutinine (H1 à H18) et 11 sous-types connus de neuraminidase (N1 à N11). Les différentes combinaisons de ces protéines déterminent le sous-type viral, par exemple H1N1 ou H3N2. Alors que les humains sont le plus souvent infectés par des virus portant à leur surface les hémagglutinines H1, H2 ou H3, les oiseaux peuvent être infectés par des virus exprimant presque tous les sous-types d’hémagglutinines (à l’exception de H17 et H18, qui ont été isolés chez les chauves-souris). Deux sous-types d’hémagglutinines seulement provoquent des maladies graves chez les oiseaux, les virus portant les hémagglutinines H5 et H7.
Une spécificité de sucre
Le virus de la grippe peut pénétrer dans les cellules et les infecter grâce à l’action de l’hémagglutinine, qui reconnaît et adhère à des molécules spécifiques à la surface des cellules-cibles (appelées récepteurs par la suite). La plupart des hémagglutinines interagissent avec les acides sialiques, une famille de dérivés d’un sucre à neuf carbones, situés généralement à l’extrémité des glycanes, ou chaînes de sucre, qui sont liées aux protéines et aux lipides de la surface des cellules. L’acide sialique peut être lié aux glycanes de différentes manières. Les deux types de liaisons les plus courantes sont les liaisons α-2,3 et α-2,61. Les virus de la grippe aviaire, comme les variants H5N1 ou H7N9, sont capables de se fixer aux glycanes porteur d’un acide sialique lié en α-2,3, qui se trouvent principalement dans les voies respiratoires et gastro-intestinales des oiseaux. L’hémagglutinine H5 aviaire est représentée sur la figure 1 (entrée de la banque de données sur les protéines (PDB) 1JSN) 2. Chez les humains, la majorité des cellules des voies respiratoires supérieures (nez, gorge et trachée) possèdent des glycanes porteurs d’acides sialiques liés en α-2,6. Les virus de la grippe humaine, comme le variant H1N1, sont capables de se fixer à ces récepteurs, ce qui les rend plus facilement transmissibles d’une personne à l’autre. L’hémagglutinine H1 est également représentée sur la figure 1 (entrée PDB 1RVT) 3.
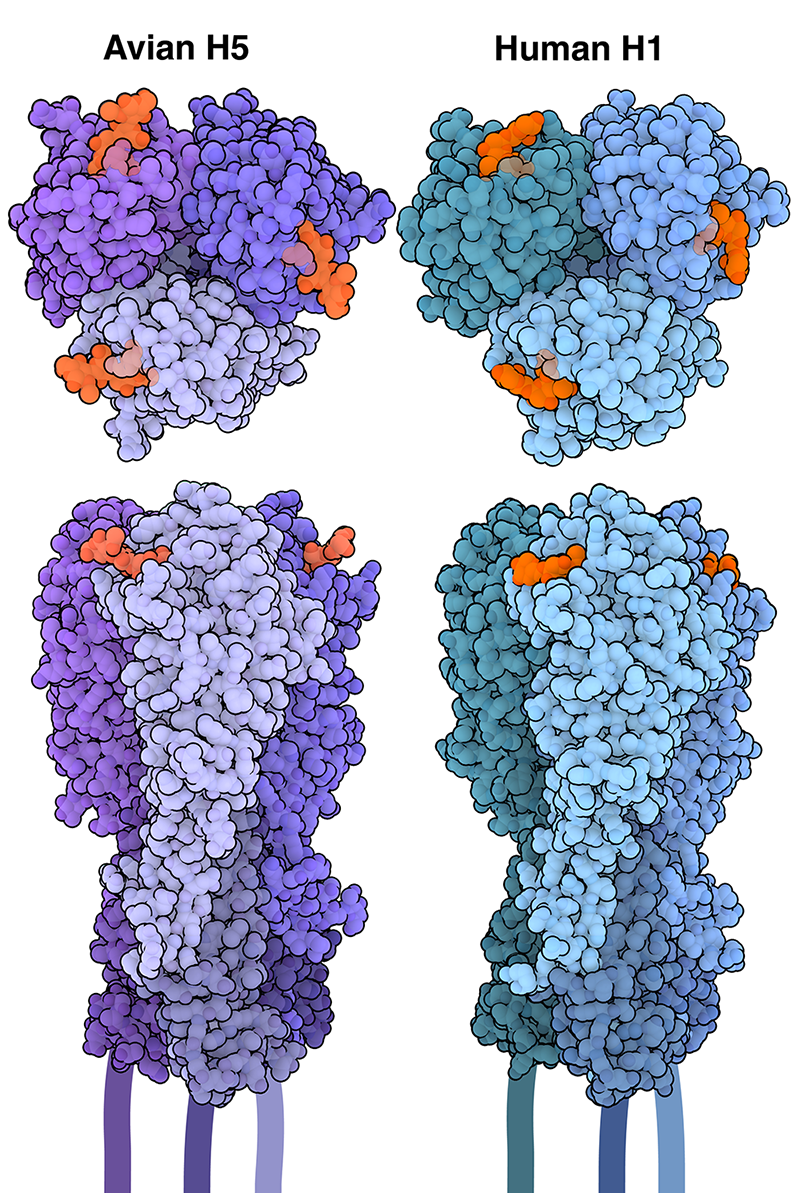
Les trimères d’hémagglutinine liés à des analogues de récepteurs d’acides sialiques (en orange) sont représentés en vue de dessus et de côté. L’hémagglutinine H5 de la grippe aviaire est représentée en violet et l’hémagglutinine H1 de la grippe humaine est représentée en bleu.
Le franchissement de la barrière d’espèces
Bien que plus efficace pour infecter les oiseaux, le virus H5N1 peut également infecter l’espèce humaine et d’autres mammifères1. Ces infections surviennent généralement à la suite d’un contact direct avec des oiseaux infectés dans des environnements très peuplés, tels que les élevages de volailles ou les marchés d’oiseaux vivants. Si la transmission entre humains du variant H5N1 est très rare et souvent inefficace, de nombreux cas de transmission entre mammifères ont récemment été signalés, notamment un foyer d’infection par un variant H5N1 dans plusieurs élevages de vaches laitières aux États-Unis. Des recherches ont montré que les glandes mammaires des vaches sont riches en glycanes porteurs d’acide sialique lié en α-2,3, ce qui permet au virus H5N1 de se répliquer et d’être libéré dans le lait, et donc potentiellement d’être transmis à d’autres vaches par l’intermédiaire d’équipements de traite contaminés 2.
La création d’une cellule recouverte de sucres
Comment les acides sialiques sont-ils ajoutés aux molécules situées à la surface des cellules ? Ceci est permis par des sialyltransférases, des enzymes qui transfèrent l’acide sialique des molécules donneuses, comme l’acide CMP-sialique, aux molécules cibles, comme les glycoprotéines et les glycolipides. Les sialyltransférases, qui se trouvent sur la membrane du Golgi, sont classées en plusieurs sous-familles en fonction du type de liaison glycosidique qu’elles forment : les α-2,3-sialyltransférases comme ST3Gal1, représentée sur la figure 2 (entrée PDB 2WNB) ajoutent l’acide sialique aux résidus de galactose via une liaison en α-2,3 3, tandis que les α-2,6-sialyltransférases comme ST6Gal1 (entrée PDB 6QVT) attachent l’acide sialique au galactose via une liaison en α-2,6 4. L’abondance de la sialylation à la surface des cellules, qui est impliquée dans divers processus comme la reconnaissance, la signalisation et la différenciation cellulaire, fait des acides sialiques des cibles idéales pour les virus 5.
Les sialyltransférases ST3Gal1 (représentée en vert clair) et ST6Gal1 (représentée en vert foncé) sont attachés à la membrane de l’appareil de Golgi (représentée en gris) par l’intermédiaire d’un lien flexible, schématisé sur cette figure. La sialyltransférase ST3Gal1 est représentée liée à une molécule de produit de la réaction (représenté en orange foncé) et à un sucre accepteur (représenté en orange clair). La partie catalytique est surlignée en vert clair. La sialyltransférase ST6Gal1 est représentée liée à un acide sialique lié en α-2,6 (orange clair).
Le saut d’obstacles
La recherche a montré que la modification d’un seul acide aminé de l’hémagglutinine H5 peut entraîner sa liaison préférentielle aux récepteurs humains et de mammifères en général (liés en α-2,6) plutôt qu’aux récepteurs aviaires (liés en α-2,3) 1. Plus précisément, le remplacement du résidu 226, qui est normalement une glutamine, par une leucine, crée un environnement plus hydrophobe qui favorise la liaison avec les récepteurs mammaliens 2. Pour explorer la façon dont les hémagglutinines H5 normales (entrées PDB 4K63 et 4K64) et mutées (entrées PDB 4K66 et 4K67) se fixent à des récepteurs d’origine aviaire ou humaine, vous pouvez utiliser par exemple le logiciel de visualisation de molécules disponible sur le site PDB-101.
L’hémagglutinine H5 normale fixant de l’acide sialique sur un glycane d’un récepteur d’origine aviaire (à gauche) et la forme mutée (à droite) fixant de l’acide sialique sur un récepteur d’origine mammalienne.
Pour aller plus loin
- Après avoir reconnu les récepteurs de la surface cellulaire et s’y être liée, l’hémagglutinine permet la fusion des membranes et l’infection. Pour en savoir plus, vous pouvez retrouver un article de la molécule du mois à ce sujet : Molecule of the Month : Hemagglutinin.
- Pour en savoir plus sur les autres composants de la grippe et sur la structure d’un virus, vous pouvez voir cette illustration de David S. Goodsell.
- Vous pouvez observer une représentation d’un vaccin contre la grippe, formulé à partir d’hémagglutinines virales, sur cette illustration de David S. Goodsell.
Ce texte correspond à la traduction par Cédric Bordi de l’article Molecule of the Month : H5 Hemagglutinin 1 écrit par Janet Iwasa et paru en février 2025 sur le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB). Les notes de bas de page correspondent à des ajouts du traducteur.
Crédits

Cet article est publié en partenariat avec le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB).