Après leur traitement au niveau de la cochlée, les informations auditives sont transmises du nerf vestibulocochléaire au cortex auditif, en passant par plusieurs relais. La tonotopie, c’est-à-dire la correspondance entre les fréquences des sons perçus et l’organisation spatiale des structures nerveuses qui traitent ces sons, est conservée tout au long du chemin. Différents traitements des informations sonores ont lieu, permettant par exemple de localiser l’origine des sons.
Percevoir son environnement est une tâche indispensable à la survie des organismes vivants et passe par la reconnaissance de différents types de stimulus : ondes électromagnétiques, molécules présentes dans l’environnement, ondes mécaniques… Chez les animaux, chacun des sens (vision, olfaction, audition…) permettant l’analyse de ces stimulus repose sur la détection d’un signal physique ou chimique par un organe récepteur spécialisé et sur la transduction de ce signal en un signal physiologique sous forme de potentiels d’action. Les organes récepteurs des différentes modalités sensorielles envoient des projections nerveuses vers les noyaux du tronc cérébral. Ces noyaux projettent ensuite vers le thalamus, structure cérébrale qui centralise les entrées vers le cortex. Les noyaux thalamiques des différentes modalités sensorielles projettent chacun vers une région dédiée du néocortex, appelées cortex sensoriels primaires. Les régions corticales attenantes qui reçoivent des projections du cortex sensoriel primaire et du thalamus sont appelées cortex sensoriels secondaires. Nous nous focalisons ici sur le système auditif. Le système auditif est le plus rapide des systèmes sensoriels et permet de détecter des sons d’alerte ou de suivre les riches variations temporelles des vocalisations, de la parole ou de la musique. Chez l'être humain, il est capable d’analyser des modifications temporelles inférieures à la milliseconde et de faire parvenir cette information jusqu’au cortex auditif en moins de 25 ms.
Le son, stimulus du système auditif
Les sons sont des ondes mécaniques qui peuvent se propager dans l’air et dans l’eau. Ils sont captés par nos oreilles et transmis au cerveau qui les décode. Le son est caractérisé par deux facteurs : sa fréquence qui se mesure en hertz (plus un son est grave, plus sa fréquence est basse ; plus il est aigu, plus sa fréquence est haute) et son intensité qui se mesure en décibel. Chez l’humain, les fréquences audibles se situent entre environ 20 Hz (en dessous, il s’agit d’infrasons) et 20 kHz (au-dessus, il s’agit d’ultrasons). La gamme de fréquences audibles varie selon les espèces. Un son pur n’est composé que d’une seule fréquence, mais de tels sons n’existent pas dans notre environnement où nous rencontrons très majoritairement des sons complexes, c’est-à-dire des mélanges de sons purs dont les différentes fréquences peuvent être séparées lors d’une analyse spectrale.
L’oreille, système auditif périphérique
L’intégration d’un son débute par la transduction d’un signal physique en signal physiologique utilisable par le système nerveux central. Le système auditif périphérique, l’oreille, a pour rôle de capter, d’amplifier et de transduire le signal acoustique. Pour plus de détails sur l’organisation et le fonctionnement de l’oreille, voir les articles Audition humaine et Déficience auditive induite par le bruit : comment peut-elle être prévenue et traitée ?, dont nous résumons dans l’encadré suivant les principales informations.
Le traitement des sons par l’oreille
Le signal acoustique capté par l’oreille externe provoque la vibration du tympan, qui entraîne la mise en mouvement des osselets : malléus (ou marteau), incus (ou enclume) et stapès (ou étrier). Ce dernier, relié à la cochlée via la fenêtre ronde, transmet l’onde sonore dans la périlymphe ce qui met en mouvement la membrane basilaire de la cochlée. Les cellules ciliées internes de l’organe de Corti vont alors être activées et se dépolariser, entraînant une libération de glutamate dans les synapses formées par les terminaisons des fibres du nerf auditif (ou nerf vestibulocochléaire, nerf VIII). Cette libération de neurotransmetteurs induit l’émission de potentiels d’action par les axones des neurones bipolaires du ganglion spiral qui forment le nerf auditif, afin de propager le message à travers les voies auditives jusqu’au cortex auditif. C’est la transduction mécano-électrique.
La voie auditive primaire véhicule l’information vers le cortex auditif primaire alors que les voies auditives non primaires véhiculent l’information vers des structures cérébrales associatives.
Anatomie de la voie auditive primaire
La voie auditive primaire commence au niveau de la cochlée pour aboutir au cortex auditif primaire. Elle véhicule l’information codée par la cochlée jusqu’au cortex auditif, chaque relais effectuant un travail de décodage et d’analyse du signal sonore pour transmettre l’information au relais supérieur.

Le nerf auditif (ou nerf vestibulocochléaire, nerf VIII) relie la cochlée au noyau cochléaire ipsilatéral, c’est-à-dire situé du même côté (gauche ou droit) du corps. L’information est ensuite envoyée au complexe olivaire supérieur puis au noyau du lemnisque latéral, puis au colliculus inférieur avant d'être relayée dans le corps géniculé médial (anciennement appelé corps genouillé médian) du thalamus, et d'aboutir enfin dans le cortex auditif primaire. Celui-ci est représenté sur une coupe frontale tandis que les autres éléments sont présentés en coupes transversales.
De la cochlée au thalamus : les relais sous-thalamiques
Les corps cellulaires des neurones du nerf auditif, ou nerf VIII, sont situés dans le ganglion spiral (environ 35 000 neurones chez l’humain 1). Le faisceau d’axones de ces neurones est composé de fibres de type 1 (90-95%), myélinisées, en provenance des cellules ciliées internes tandis que les fibres de type 2 (5-10%), non myélinisées, proviennent des cellules ciliées externes. Le axones du nerf auditif de chacune des deux cochlées projettent sur le noyau cochléaire ipsilatéral2 (Figures 1 et 2). Au sein de cette structure, il existe sept types morphologiques de neurones dans le noyau ventral et deux types dans le noyau dorsal, présentant également chacun des caractéristiques électrophysiologiques différentes. Les noyaux cochléaires possèdent de nombreux circuits internes avec des rôles activateurs ou inhibiteurs entre les neurones, contribuant à l’analyse du signal auditif.
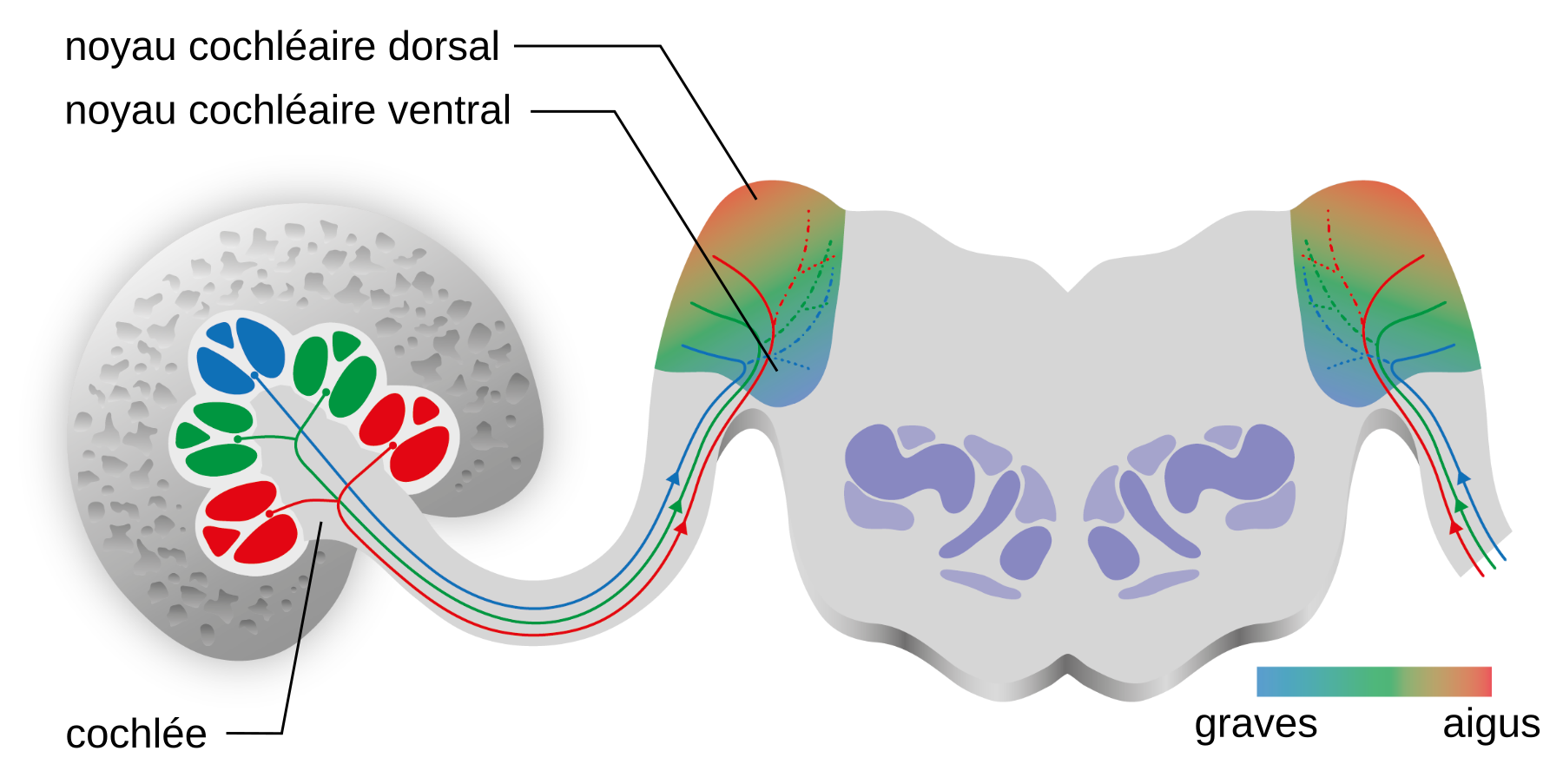
Les neurones de type I relient les cellules ciliées internes de la cochlée et le noyau cochléaire, formant le nerf auditif. Les dégradés de couleurs, du bleu au rouge, marquent la tonotopie cochléaire et sa projection dans le noyau cochléaire. Le noyau cochléaire présente ainsi une organisation tonotopique selon un axe dorso-ventral.
Le second relais du système auditif est le complexe olivaire supérieur (Figures 1 et 3). Les neurones du noyau cochléaire projettent sur l’olive supérieure de manière majoritairement controlatérale (Figure 3). Ce complexe se compose de trois noyaux principaux : le noyau olivaire supérieur latéral, le noyau olivaire supérieur médian et le noyau médian du corps trapézoïde.
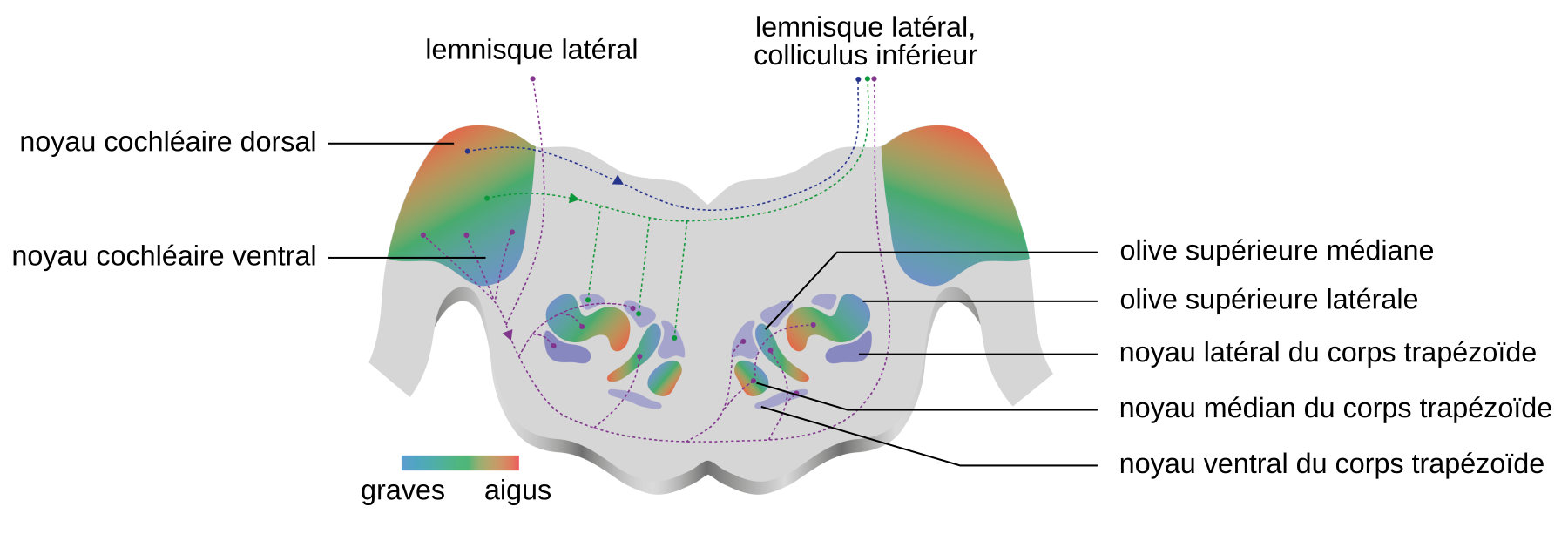
La partie ventrale des noyaux cochléaires émet des projections vers le noyau médian du corps trapézoïde controlatéral, le noyau ventral du corps trapézoïde controlatéral, le noyau latéral du corps trapézoïde ipsilatéral, les deux olives supérieures médianes, les deux olives supérieures latérales, les deux lemnisques latéraux et le colliculus inférieur controlatéral. Le noyau cochléaire dorsal émet des projections vers le lemnisque latéral et le colliculus inférieur controlatéraux. Les dégradés de couleurs, du bleu au rouge, traduisent l’organisation tonotopique des structures.
NB : Sur ce schéma, les noyaux cochléaires et les olives supérieures sont figurés dans le même plan par commodité de représentation. En réalité, ces structures n'occupent pas exactement la même place le long de l'axe antéropostérieur (voir Figure 1).
Le troisième relais de la voie auditive principale est le lemnisque latéral (Figures 1 et 4) qui reçoit les projections axonales des neurones de l’olive supérieure. Il est composé d’un noyau ventral et d’un noyau dorsal. Le noyau dorsal du lemnisque latéral reçoit les projections excitatrices et inhibitrices des noyaux olivaire supérieur médian et latéral alors que le noyau ventral du lemnisque latéral est innervée par le noyau cochléaire controlatéral et le noyau médian du corps trapézoïde ipsilatéral.
Les projections du lemnisque latéral ciblent ensuite principalement le colliculus inférieur (Figures 1 et 4) au niveau du mésencéphale. Cette structure est divisée en trois régions : le noyau central, le cortex dorsal et le cortex latéral. La totalité des afférences auditives ascendantes forme un relais dans un de ces noyaux, ce qui fait que le colliculus inférieur occupe une place stratégique. Le noyau central reçoit les projections du lemnisque latéral ventral et dorsal, ainsi que du complexe olivaire. Le cortex dorsal reçoit les projections du lemnisque latéral dorsal. Quant au cortex latéral, il reçoit les projections du lemnisque latéral dorsal et du noyau central du colliculus inférieur.
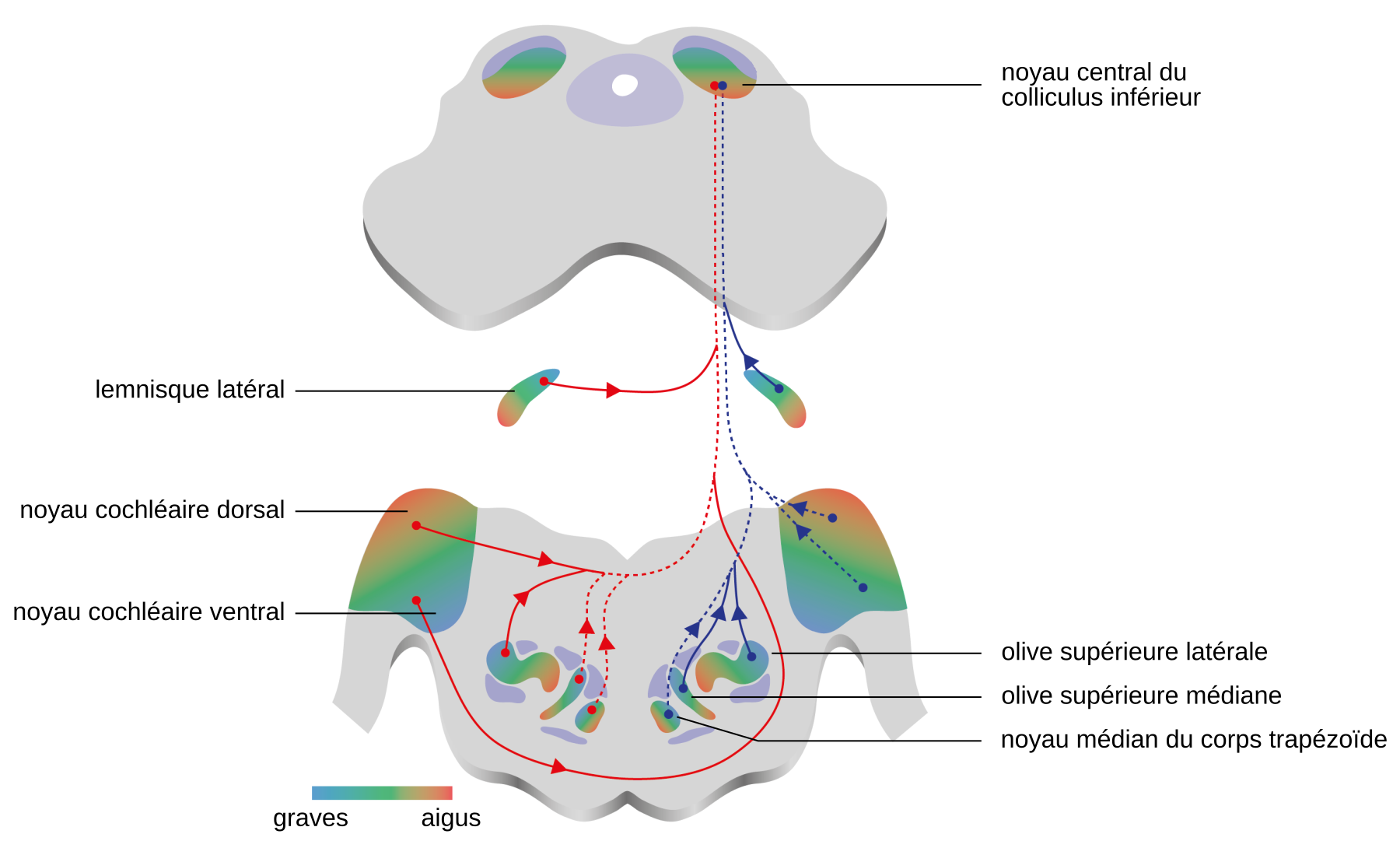
Le noyau central du colliculus inférieur reçoit des afférences directes de faisceaux d’origine controlatérale, issus des noyaux cochléaires ventral et dorsal, des olives supérieures latérales et médiane, du corps trapézoïde et du lemnisque latéral (traits rouges). Il reçoit également les projections de ces mêmes structures mais du côté ipsilatéral (traits bleus). Les traits pleins marquent les ascendances majeures, les pointillés les mineures. Les dégradés de couleurs, du bleu au rouge, marquent les tonotopies connues des structures.
Le thalamus auditif : le corps géniculé médial
Le dernier relais sous-cortical de la voie auditive primaire est le corps géniculé médial (Figures 1 et 5) qui reçoit les projections ipsilatérales du colliculus inférieur. C’est le relais thalamique entre le tronc cérébral et le cortex auditif. Il est composé de plusieurs noyaux, qui se distinguent par leur densité, la morphologie de leurs cellules, ainsi que par les projections qu’ils reçoivent et envoient : la partie ventrale, la partie dorsale et la partie médiane ou magnocellulaire 1. La partie ventrale du corps géniculé médial reçoit les connexions en provenance du noyau central du colliculus inférieur et fait donc partie de la voie auditive primaire.
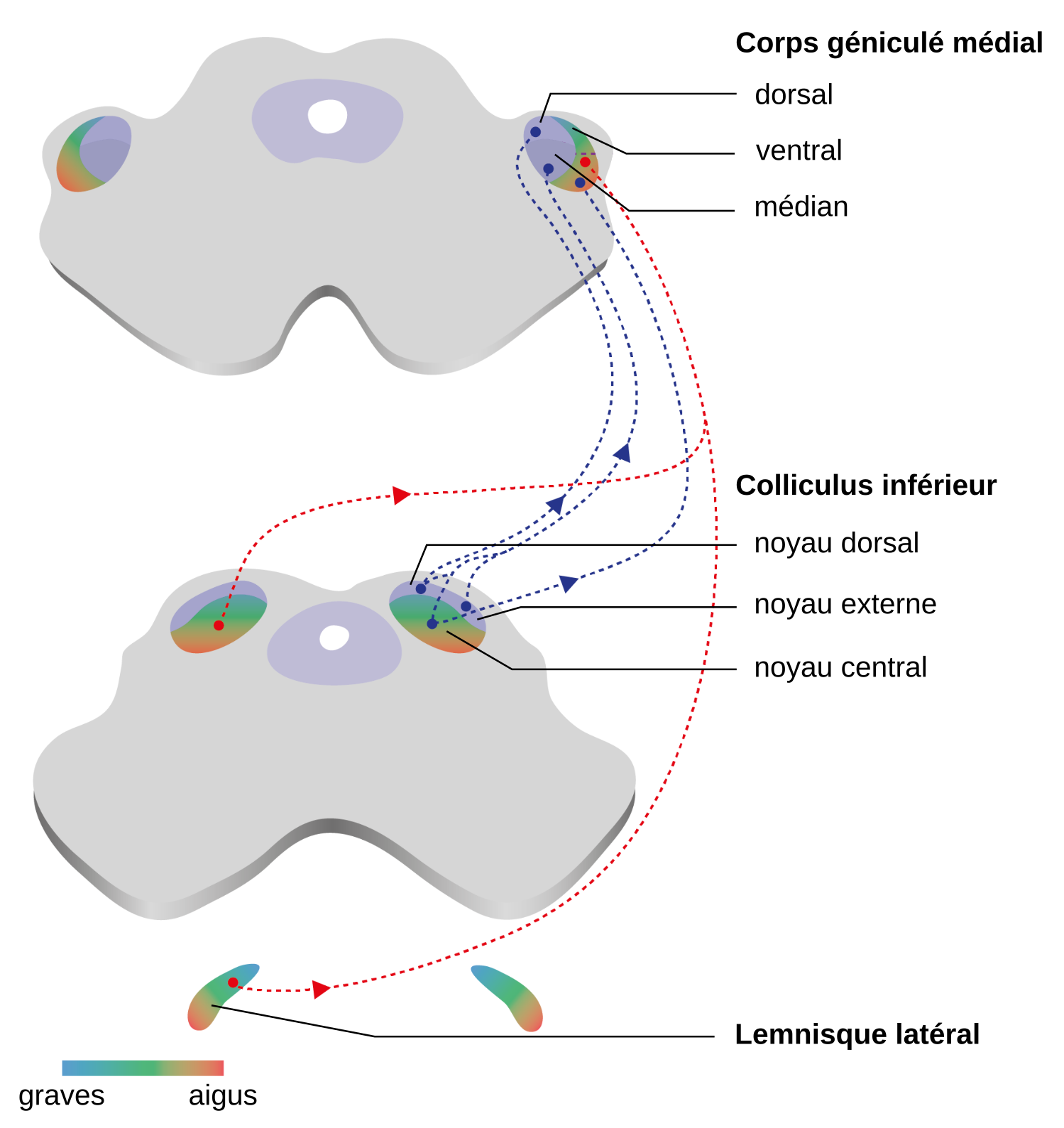
Le corps géniculé médial (anciennement appelé corps genouillé médian) est subdivisé en trois parties, ventrale, médiane et dorsale. Les principales afférences proviennent du lemnisque latéral et du colliculus inférieur controlatéral (traits rouges) mais aussi des noyaux central, externe et dorsal du colliculus inférieur (traits bleus).
Cortex auditif
Le cortex auditif constitue le dernier relais de la voie ascendante du système auditif central. Il est localisé au niveau du lobe temporal et est subdivisé en plusieurs aires corticales en fonction de critères anatomiques et physiologiques (Figure 6). Chez l’ensemble des mammifères, le cortex auditif se compose d’une ou plusieurs aire(s) tonotopique(s) entourée(s) de plusieurs aires non tonotopiques. Néanmoins, selon les espèces, le nombre d’aires identifiées est variable : trois ou cinq chez le rat 12, treize chez le chat 3 ou encore douze ou plus chez le macaque 4. En général, l’aire auditive primaire (A1) correspond à la région centrale du cortex auditif qui reçoit les projections directes de la voie auditive ascendante en provenance du corps géniculé médial.
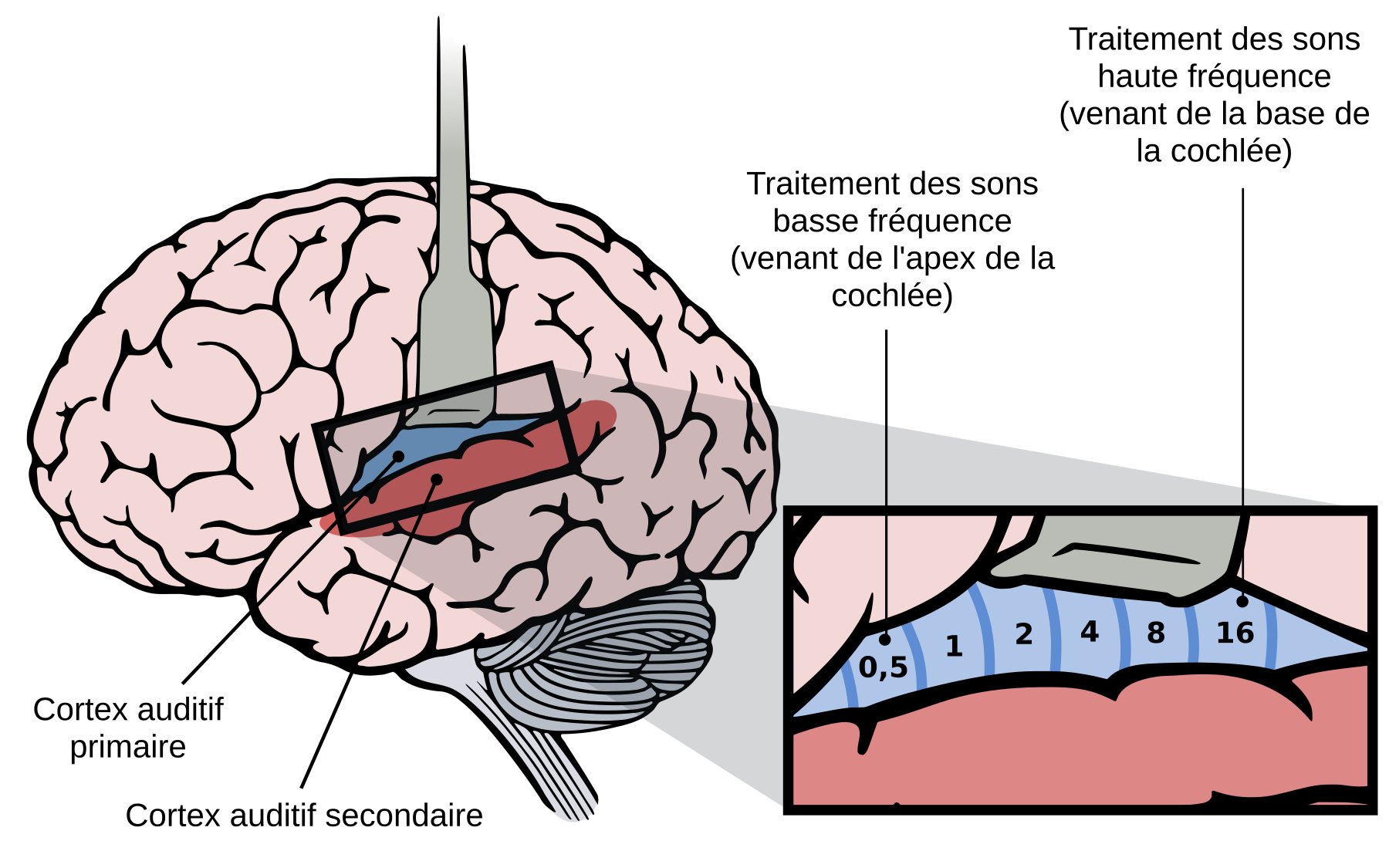
L’aire auditive primaire (en bleu) est située dans le tiers postérieur du gyrus temporal supérieur. Les basses fréquences sont traitées dans la partie antérieure du cortex auditif primaire alors que les hautes fréquences sont traitées dans sa partie postérieure. L’aire auditive secondaire est représentée en rouge.

Comme tous les cortex sensoriels, le cortex auditif est organisé en six couches composées de types cellulaires différents (Figure 7). Deux types cellulaires principaux se trouvent au sein du cortex auditif : les neurones pyramidaux (70-80%, excitateurs glutamatergiques) et les interneurones (20-30%, excitateurs glutamatergiques ou inhibiteurs GABAergiques). Les interneurones inhibiteurs sont d’une extrême diversité et classés selon des critères morphologiques, moléculaires et électrophysiologiques 1.
Les neurones de la partie ventrale du corps géniculé médial projettent sur le cortex auditif primaire au niveau de la couche IV et du bas de la couche III. De façon schématique, chaque neurone du corps géniculé médial qui projette vers le cortex auditif génère un arbre de fibres qui se ramifie horizontalement entraînant de nombreux contacts sur les neurones de la couche IV. Ce système permet l’amplification du signal de la voie auditive. Les neurones pyramidaux de la couche IV projettent alors vers les neurones des couches II et III qui distribuent ensuite l’information aux neurones de la couche V qui projettent eux-mêmes vers les neurones de la couche VI. Une partie des projections des neurones de la couche VI projette de façon rétrograde sur le corps géniculé latéral afin de réguler la voir auditive.
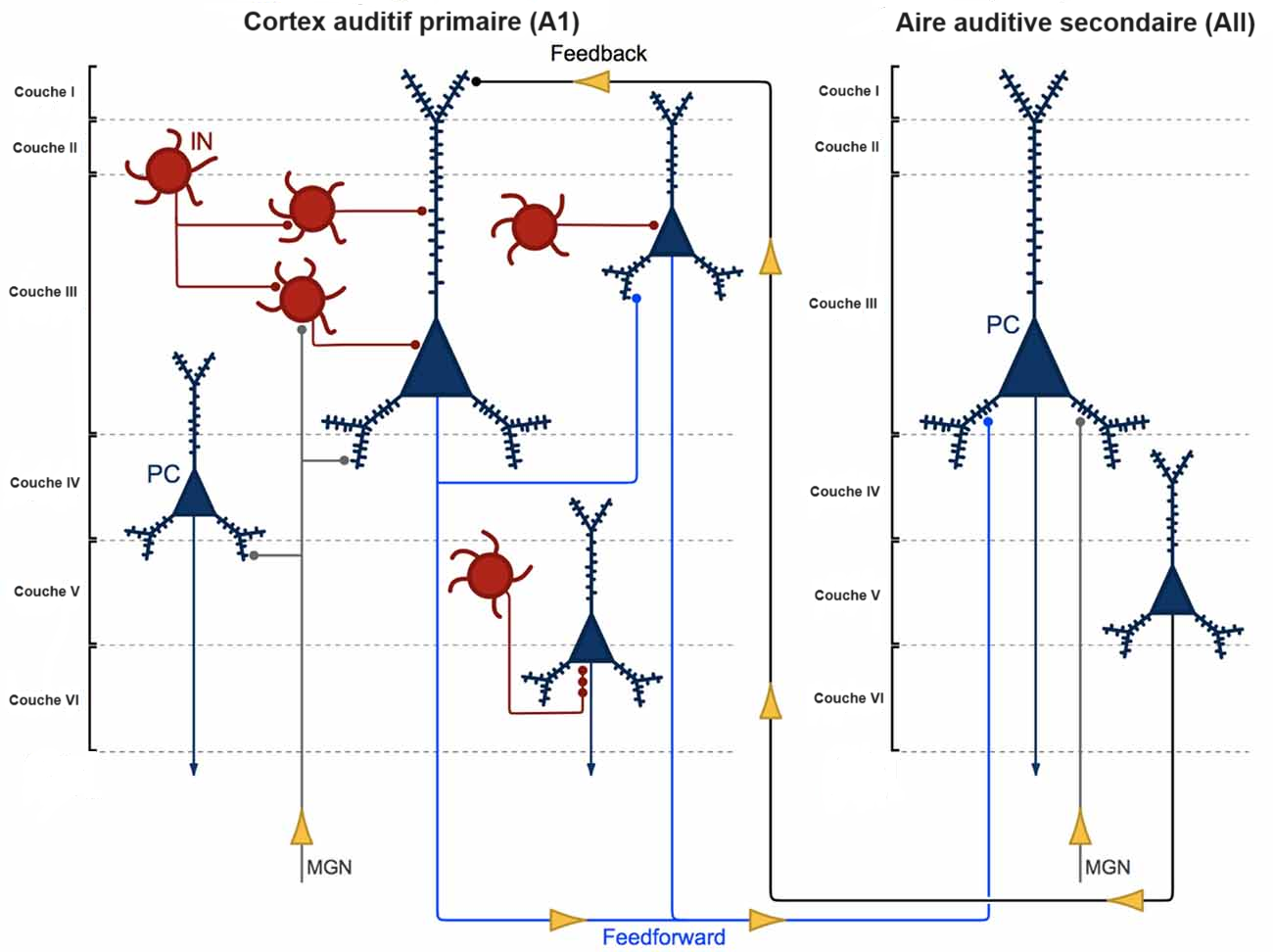
Les projections thalamiques du noyau genouillé médian (MGN) arrivent sur les cellules pyramidales (PC, en bleu) et les interneurones (IN, en rouge). Les cellules pyramidales de la couche III du cortex auditif primaire envoient des projections à d’autres celles pyramidales de la couche III du même cortex et du cortex auditif secondaire. Celles de la couche V du cortex auditif secondaire envoient à leur tour des projections aux neurones de la couche I du cortex auditif primaire. Les interneurones de la couche II, III et V régulent l’activité des cellules pyramidales localement.
Fonctions de la voie auditive primaire
La cochlée permet la décomposition d’un signal acoustique en fonction de sa fréquence, de son intensité et de son décours temporel (évolution du signal au cours du temps). Ces informations sont ensuite conservées dans les différents relais du système auditif.
Codage de l’intensité
L’intensité du son est codée par deux éléments : la fréquence de décharge des neurones et le nombre de neurones activés. En effet, pour des stimulations d’intensité croissante, la fréquence des potentiels d'action augmente. Par ailleurs, plus un stimulus est intense, plus le nombre de fibres recrutées augmente. En effet, il existe trois populations de fibres auditives : des fibres à bas seuil ayant une activité basale élevée, des fibres à seuil et activité moyennes, et enfin des fibres à haut seuil présentant une faible activité spontanée. Une stimulation sonore de faible intensité active les premières fibres mais sature rapidement. Des stimulations d’intensité plus fortes recrutent ensuite les secondes et troisièmes fibres. Ceci entraîne des décharges plus fortes au niveau du nerf auditif et tout le long de la voie auditive primaire.

Tonotopie et codage de la fréquence
Les stimulus auditifs sont décomposés en bande de fréquence par la cochlée. Du fait des propriétés biomécaniques de la membrane basilaire – rigide à la base et souple à l’apex – les ondes sonores de hautes fréquences provoquent une déflexion maximale de cette membrane à la base alors que les basses fréquences provoquent une déflexion maximale à l’apex. Cette organisation spatiale correspondant à l’organisation en fréquence du signal acoustique est nommée tonotopie. Celle-ci est conservée tout au long de la voie auditive primaire (Figures 2 à 6).
La tonotopie de la cochlée est conservée au niveau du noyau cochléaire : la partie dorsale est reliée à la base de la cochlée (hautes fréquences) tandis que la partie ventrale reçoit les informations en provenance de l’apex de la cochlée (basses fréquences). Les trois noyaux de l’olive supérieure présentent également une distribution tonotopique : les basses fréquences dans les aires dorsales ou latérales et les hautes fréquences dans les aires plus ventrales et internes. Les deux noyaux du lemnisque latéral sont eux aussi organisés de manière tonotopique 12. Dans le colliculus inférieur, seul le noyau central est composé d’un grand nombre de neurones organisés de façon tonotopique. La partie ventrale du corps géniculé médial présente également une organisation tonotopique 3.
Comme dans les autres étages du système auditif, les neurones du cortex auditif primaire déchargent en rapport avec la fréquence du son et ils présentent des fréquences caractéristiques couvrant le spectre des fréquences audibles. La représentation tonotopique des basses fréquences est latérale et rostrale alors que la représentation des hautes fréquences est médiane et caudale (Figure 6). Le cortex auditif est formé de rangées de neurones transversales dans lesquelles tous les neurones ont à peu près les mêmes fréquences caractéristiques.
Il est intéressant de noter que dans le corps géniculé médial, les neurones de la partie dorsale répondent davantage aux sons complexes, comme des vocalises, suggérant un rôle dans la discrimination des motifs sonores.
Localisation spatiale des sons
Notre capacité d’orientation dans notre environnement dépend en partie de la faculté à localiser des sons dans l’espace, aussi bien sur le plan horizontal que vertical.
Sur le plan horizontal, la localisation des sons est effectuée par comparaison entre les informations sonores qui parviennent aux deux oreilles. Si la source sonore ne se situe pas directement face à lui, le son mettra plus de temps à parvenir à l’une des deux oreilles. Les sons n’atteignent donc pas les deux oreilles de façon synchrone et le décalage temporel entre les sons qui atteignent les deux oreilles permet d’apprécier la localisation de la source sonore dans l’espace. Le système auditif est ainsi capable de discriminer des différences de délai de l’ordre de quelques dizaines de microsecondes (µs), délai extrêmement court en comparaison avec la durée d’un potentiel d’action (~ 1 ms). Comment le système auditif peut-il effectuer une discrimination aussi fine ? Le complexe olivaire supérieur est considéré comme le siège de la détection de la localisation des sources sonores 4, grâce notamment à la détection du délai interaural. Ce délai correspond au fait qu’un son mettra plus de temps à parvenir à l’une des oreilles par rapport à l’autre.
Pour les sons graves (< 1 400 Hz), c’est la différence interaurale de temps qui est utilisé. C’est principalement l’olive supérieure médiane qui permet d’analyser la différence de phase entre deux stimuli sonores grâce à des neurones détecteurs de coïncidence. Ces neurones sont bipolaires et reçoivent les informations excitatrices venant des deux noyaux cochléaires, gauche et droit. Un mécanisme propose que la différence de longueur des voies ipsi et controlatérales induit une activation différentielle des neurones de l’olive supérieure médiane, c’est-à-dire à des endroits spécifiques dans la structure dépendant de la localisation de la source sonore selon le principe des lignes de délai (Figure 8). Les neurones de l’olive supérieure médiane constituent donc une carte de la position des sons.

L’analyse de la différence de phase s’effectue par une augmentation progressive du temps de conduction le long des voies véhiculant l’information. Une information sonore provenant de l’oreille droite atteint les neurones de l’olive supérieure médiane dans le sens E-A alors que les informations sonores provenant de l’oreille gauche atteint les mêmes neurones dans le sens A-E. Si les deux signaux sont synchrones, c’est-à-dire pour un son émis dans l’axe de la tête), c’est le neurone C qui répondra. Si le son atteint d’abord l’oreille droite, c’est-à-dire pour un son provenant du côté droit de la tête, ce sera plutôt au niveau du neurone A ou B que se produira la coïncidence. Et lorsqu’un son atteint d’abord l’oreille gauche, c’est-à-dire pour un son provenant du côté gauche de la tête, ce sera plutôt au niveau du neurone D ou E que se produira la coïncidence.
Pour les sons plus aigus (> 1 400 Hz), la localisation dans l’espace repose sur la différence d’intensité sonore perçue par les deux oreilles. Cette fois-ci c’est principalement l’olive supérieure latérale et le noyau médian du corps trapézoïde qui permettent de détecter des différences interaurales d’intensité. Lorsqu’un son arrive au niveau de l’oreille gauche, il active les neurones de l’olive supérieure latérale du côté gauche et via l’intermédiaire du noyau médian du corps trapézoïde, inhibe les neurones de l’olive supérieure latérale du côté droit. Et inversement lorsqu’un son arrive au niveau de l’oreille droite. Ainsi, cette interaction excitation-inhibition se traduit par une plus forte excitation de l’olive supérieur latérale située du même côté que la source sonore. Cette différence d’activation liée à l’intensité sonore permet ainsi de coder la localisation des sons aigus sur un plan horizontal.
Étant donné que les deux oreilles se trouvent sur le même plan horizontal, les mécanismes binauraux précédemment décrits ne s’appliquent pas pour la localisation verticale d’une source sonore. La détection de l’élévation d’une source sonore est permise par la forme du pavillon de l’oreille, qui transmet davantage les composantes de hautes fréquences d’un son quand la source est située en hauteur que lorsqu’elle est au même niveau que l’oreille.
Voie auditive non primaire
Le rôle de cette voie, qui regroupe plusieurs modalités sensorielles, est de permettre une sélection du type d’informations à traiter en priorité. Cette voie est constituée de petites fibres qui, partant des noyaux cochléaires, rejoignent le système réticulaire ascendant commun à toutes les modalités sensorielles. Après plusieurs relais dans la formation réticulée, puis dans le thalamus, cette voie auditive non primaire aboutit au cortex polysensoriel. C’est dans la formation réticulée du tronc cérébral et du mésencéphale que les informations auditives sont intégrées aux autres informations sensorielles pour participer au tri de la modalité prioritaire à un instant donné. De la même façon que les autres systèmes sensoriels non spécifiques, elle participe à différentes fonctions complexes telles que la motivation, l’éveil ou encore les émotions.
Intégration comportementale de l’information auditive
La connexion entre la voie auditive et les autres aires cérébrales joue un rôle dans l’intégration du signal sonore aux autres informations issues de l’environnement. Les neurones du thalamus auditif projettent sur plusieurs aires corticales incluant les aires auditives secondaires, le cortex insulaire, le cortex temporal, les aires suprassylviennes et les aires ectosylviennes. C’est dans cette structure que se fait un important travail d’intégration avec notamment la préparation d’une éventuelle réponse motrice vocale. La partie médiane du thalamus auditif est une structure multisensorielle recevant des afférences auditives et non auditives. Certains de ses neurones projettent sur l’amygdale suggérant une implication dans la signification émotionnelle des sons.
Régulation des voies auditives
La neuromodulation est le processus par lequel plusieurs neurotransmetteurs (dopamine, noradrénaline, adrénaline, sérotonine, acétylcholine, GABA…) régulent plusieurs populations de neurones. Ils sont libérés dans l’environnement neuronal afin d’influencer le fonctionnement des neurones et des réseaux neuronaux. Le rôle des neuromodulateurs a fait l’objet de nombreuses recherches depuis le milieu du siècle dernier. Le mode de libération et la localisation ubiquitaire des récepteurs des neuromodulateurs font de ces neurotransmetteurs des modulateurs à grand échelle des processus cérébraux, y compris des systèmes sensoriels. Il n’existe néanmoins que peu d’études sur l’influence des neuromodulateurs sur la sélectivité des réponses des neurones des cortex sensoriels.
Le locus cœruleus, source principale de noradrénaline du système nerveux central, projette vers un nombre important de régions du système nerveux central, incluant des régions impliquées dans l'audition : le noyau cochléaire, le colliculus inférieur, le corps géniculé médial et le cortex auditif 123. Il existe également une innervation cholinergique du cortex auditif, comme pour tous les cortex sensoriels en général. Le noyau cochléaire et la cochlée reçoivent une innervation cholinergique depuis le complexe olivaire supérieur 4. La modulation des cortex sensoriels par la noradrénaline et l’acétylcholine est souvent liée à des contextes d’apprentissages ou plus généralement de perception de stimuli sensoriels. Ces deux neuromodulateurs modifient l’activité spontanée des neurones corticaux (l’acétylcholine l’augmente alors que la noradrénaline la diminue), les réponses évoquées des neurones corticaux, la sélectivité des réponses corticales (la noradrénaline augmente le seuil de réponse des neurones à l’ensemble des fréquences, ce qui réduit la gamme de fréquences à laquelle le neurone répond, alors que l’acétylcholine facilite les réponses neuronales préférentiellement aux sons purs et diminue les seuils de réponse).
Au sein du cortex, les aires primaires auditives projettent vers un grand nombre de noyaux auditifs sous-corticaux ipsilatéraux afin de moduler leur activité, comme la sensibilité des neurones du colliculus inférieur aux indices de localisation des sources sonores 5. En plus de cette innervation directe, il existe des boucles de régulation locales d’un noyau sur le précédent, formant ainsi une chaîne descendante du cortex auditif primaire jusqu’aux cellules ciliées de la cochlée. Un faisceau de fibres projette depuis le cortex auditif primaire vers le colliculus inférieur, puis deux groupes de fibres projettent vers l’olive supérieure et le noyau cochléaire respectivement. Un ensemble de fibres innerve les cellules ciliées externes de la cochlée depuis l’olive supérieur, et un ensemble de fibres en provenance du noyau cochléaire innerve les cellules ciliées internes. Ce système permet une modulation rapide (trois synapses) de l’activité des récepteurs sensoriels.