

Le terme « champignon » ne désigne pas un groupe monophylétique mais plusieurs taxons distincts, caractérisés par un mode de vie filamenteux et une nutrition par absorbotrophie. Cet article présente les différents groupes d'êtres vivants désignés sous le nom de « champignon ».
Retrouvez également cet article dans un dossier consacré aux symbioses sur le site de formation du Muséum national d'histoire naturelle.
Le terme « champignon » désigne plusieurs organismes, ou portions d’organismes, selon le point de vue depuis lequel on les observe. Historiquement, les champignons sont les objets d’étude des mycologues ; pour les promeneurs, les champignons sont des produits forestiers que l’on ramasse après la pluie. Scientifiquement, on regroupe sous le nom de « champignons » des êtres vivants eucaryotes réunissant deux caractéristiques :
- ils ont un mode de vie filamenteux, c’est-à-dire que leur appareil végétatif est composé de filaments, appelés hyphes, l’ensemble des hyphes formant le mycélium ;
- ils se nourrissent par absorbotrophie : ils sécrètent dans leur environnement des enzymes digestives qui dégradent la matière organique en nutriments, puis ils absorbent ces nutriments. Les champignons sont non-photosynthétiques, donc hétérotrophes au carbone : ils dépendent d’autres êtres vivants pour leur nutrition carbonée.
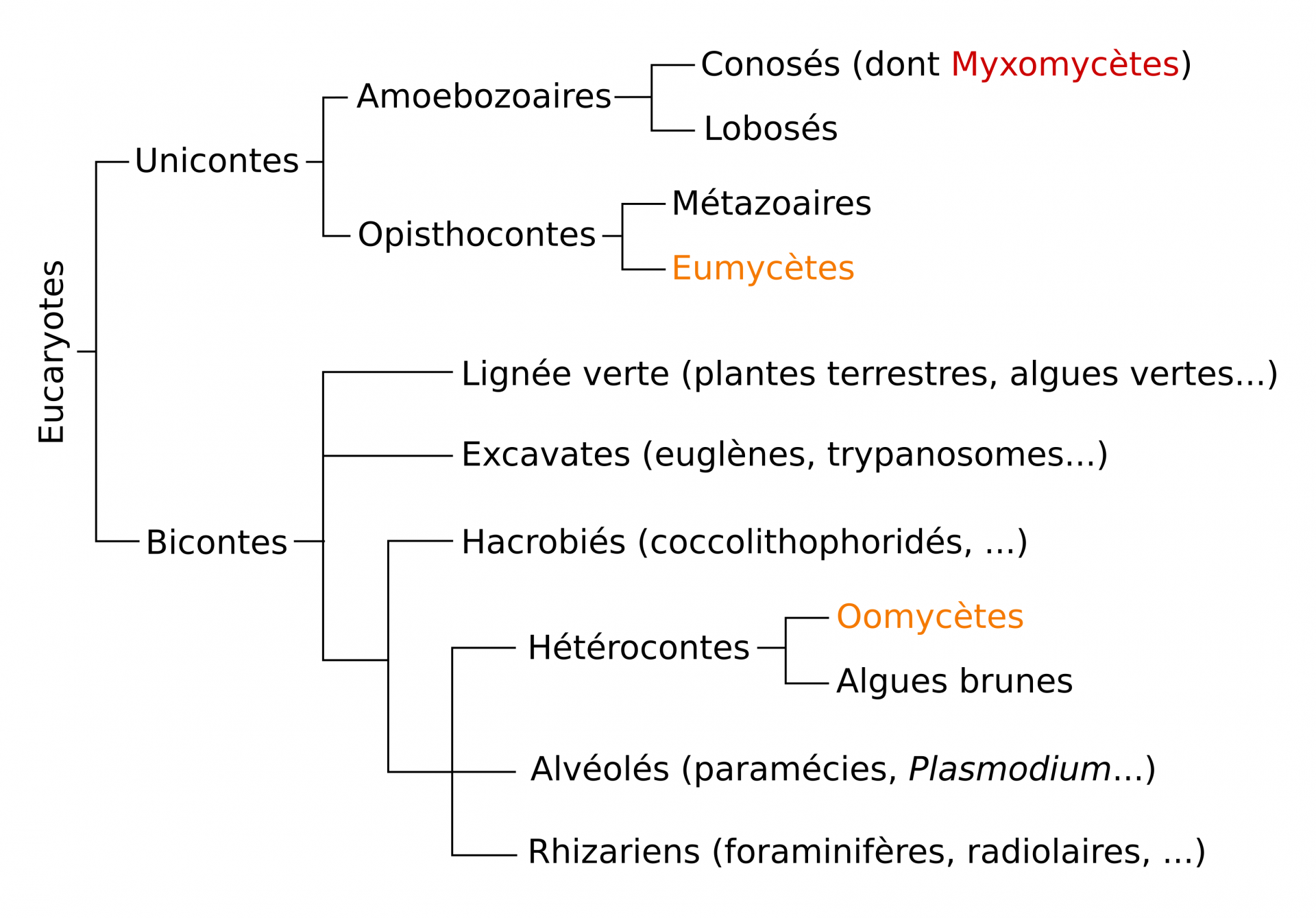
Les groupes en orange sont les groupes rassemblés sous le nom de « champignon » (sens large). D’après Lecointre et Le Guyader (2016).
Ces caractéristiques, qui peuvent être perdues secondairement dans certains taxons, permettent de regrouper des organismes appartenant à des groupes distincts dans la phylogénie des Eucaryotes, en orange dans la figure 1 : les Oomycètes et les Eumycètes (c’est donc un groupe polyphylétique). Cependant, l’utilisation du terme « champignon » pour désigner un groupe polyphylétique ne fait pas consensus. Dans leur ouvrage de référence, Classification phylogénétique du vivant, tome 1 (Belin, 2016), Lecointre et Le Guyader utilisent le terme « champignons » pour désigner le groupe monophylétique des Eumycètes et de leur taxon frère, les Microsporidies, en excluant les Oomycètes. On utilisera ici le terme de « champignon » (sens large) pour désigner toutes les formes de vie filamenteuse et absorbotrophes (Oomycètes et Eumycètes) ; on gardera le terme d’Eumycètes pour parler des « champignons vrais » (sens strict), c’est-à-dire les organismes filamenteux absorbotrophes à paroi chitineuse, qui incluent la plupart des espèces connues du grand public (Ascomycètes et Basidiomycètes). Les Myxomycètes, en rouge dans la figure 1, seront rapidement abordés puisqu’ils sont aussi traditionnellement étudiés par les mycologues, bien qu’ils n’aient pas grand-chose à voir avec des champignons.
Trois groupes historiquement étudiés par les mycologues
Les Myxomycètes, des « amibes collectives »
Les Myxomycètes, bien que parfois considérés comme des champignons, s’en distinguent par deux différences majeures : ils se nourrissent par phagocytose et non par absorbotrophie, et ils n’ont pas de mycélium. Ils appartiennent au groupe des Conosés, au sein des Amoebozoaires (figure 1). Le terme « amibe » désigne des organismes produisant des pseudopodes et changeant de forme lorsqu’ils se déplacent. Il est donc parfois utilisé pour qualifier les Myxomycètes, qui forment des masses gélatineuses capables de mouvement. Cependant, il s’agit d’un groupe polyphylétique : le terme« amibe » n’est donc plus utilisé en phylogénie, et la majorité des organismes anciennement appelés ainsi se trouvent aujourd’hui dans le groupe des Lobosés.
Au cours de leur cycle de vie, les Myxomycètes passent par des stades morphologiques variés : en conditions favorables, ils vivent sous la forme d’un plasmode, une très grande cellule entourée d’une paroi cellulosique qui contient de nombreux noyaux. Cette cellule plasmodiale forme une masse gélatineuse (Myxomycète vient du grec muxa, mucus, en référence à leur aspect gluant) qui peut se déplacer à la recherche de nourriture et se diviser en plusieurs plasmodes si les ressources sont suffisantes (on parle de multiplication végétative).

Les Oomycètes, des « champignons » au sens large

Les Oomycètes font partie des Bicontes, groupe monophylétique qui rassemble les Eucaryotes dont les cellules mobiles possèdent deux flagelles (du grec bi, deux, et kontos, flagelle), comme les plantes terrestres. Au sein des Bicontes, les Oomycètes appartiennent aux Hétérocontes (aussi appelés Straménopiles), et sont donc proches du groupe des algues brunes. Parmi les Oomycètes, on dénombre plusieurs pathogènes connus, comme Phytophtora infestans, responsable du mildiou de la pomme de terre, ou Plasmopora viticola, agent du mildiou de la vigne. D’autres espèces sont parasites d’animaux, ou dégradent la matière organique morte du sol (on les appelle saprophytes).
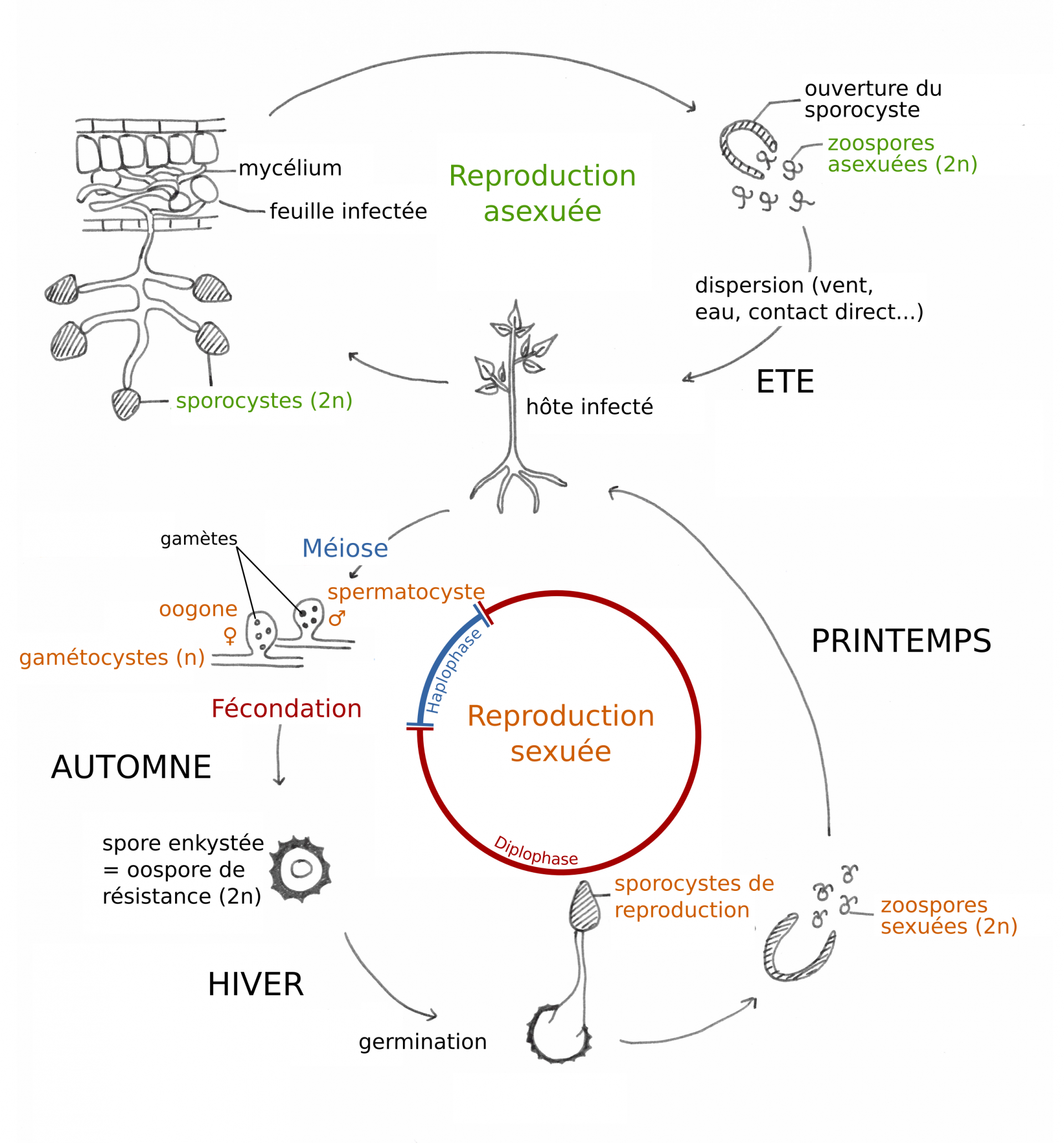
Les Oomycètes sont des organismes constitués d’hyphes non cloisonnés (aussi appelés siphons), à paroi cellulosique. Comme tous les « champignons » au sens large, les Oomycètes peuvent se reproduire de manière sexuée ou asexuée (c’est-à-dire avec ou sans brassage du matériel génétique, figure 4). Lorsque les conditions sont favorables, la reproduction asexuée est assurée par les zoospores, des spores rendues mobiles par leurs deux flagelles et produites par mitose au sein de structures spécialisées (les sporocystes). Ces zoospores asexuées assurent la dispersion de l’oomycète : emportées par le vent, par l’eau ou par contact avec un organisme malade, elles sont déposées sur des organismes sains qu’elles infectent en germant, et où elles continuent leur cycle de vie.
Lorsque les conditions se dégradent a lieu la reproduction sexuée, qui fait intervenir un hyphe mâle et un hyphe femelle. Ces hyphes possèdent des structures spécialisées, les gamétocystes, dans lesquelles sont formés les gamètes mâles et femelles par méiose. La fusion des hyphes mâle et femelle (plasmogamie) met en contact les noyaux haploïdes mâle et femelle qui fusionnent à leur tour (caryogamie) pour former des œufs diploïdes appelés oospores. Ces oospores sont enkystées dans une paroi de mélanine épaisse qui leur permet de passer la mauvaise saison. Lorsque les conditions sont de nouveau favorables, l’oospore germe, met en place un sporocyste et libère les zoospores qui redonnent une nouvelle génération après germination.

Les groupes en gras sont les phylums les plus importants en termes de nombre d’espèces et ceux dans lesquels on retrouve des espèces mycorhiziennes. Le point orange représente le plus récent ancêtre commun aux Mucoromycètes, Ascomycètes et Basidiomycètes ; il correspond à l’origine évolutive de la capacité à mettre en place des symbioses mycorhiziennes telles qu’on les connaît aujourd’hui, et à produire des sporocarpes. D’après Spatafora et coll. (2017).
Les Eumycètes, des champignons « vrais »
À la différence des Oomycètes, les Eumycètes (du grec eu, vrai, et mukes, champignon) font partie des Unicontes, groupe monophylétique qui rassemble les Eucaryotes dont les cellules mobiles possèdent un seul flagelle. La position postérieure de ce flagelle les place, comme les Animaux, au sein des Opistochontes (du grec optisthios, postérieur). Les Eumycètes sont donc évolutivement plus proches des animaux que des plantes, bien qu’ils aient été longtemps assimilés à des plantes à cause de leur apparente immobilité !
L’appareil végétatif des Eumycètes est composé de filaments à paroi chitineuse (et non cellulosique comme les deux groupes précédents), qui constituent la majeure partie de l’organisme. La partie visible, le « champignon » des cueilleurs, qui peut être comestible ou au contraire toxique, n’est que la structure reproductive de l’individu (le sporocarpe). La reproduction des Eumycètes a ceci de particulier qu’il n’y a pas de gamète mobile libéré dans le milieu. Au contraire, les noyaux haploïdes sont contenus dans les hyphes et c’est la fusion de ceux-ci qui permet la fécondation.
Les Eumycètes présentent une grande diversité de formes et de modes de vie. Ils sont classés en plusieurs groupes, dont les mieux connus et les plus importants en termes de rôle écologique sont les Gloméromycètes (inclus dans le groupe des Mucoromycètes), les Ascomycètes et les Basidiomycètes (figure 5).

A. Moisissures du genre Rhizopus (Mucoromycètes) se développant sur des tomates (photographie par Calimo, Wikimedia). B. Ascocarpe de truffe (Ascomycètes) ; photographie par Michel Royon, Wikimedia. C. Basidiocarpe d’amanite tue-mouche, Amanita muscaria (Basidiomycètes) ; photographie par JJ Harrison, Wikimedia.
Les trois principaux groupes d’Eumycètes
Les trois groupes d’Eumycètes les plus importants en nombre d’espèces et en rôles écologiques – Mucoromycètes, Ascomycètes et Basidiomycètes – sont caractérisés par l’apparition de relations végétaux-champignons originales : les lichens et les symbioses mycorhiziennes. Dans les deux cas, un champignon s’associe avec un organisme photosynthétique (plantes terrestres dans le cas des mycorhizes, algues et/ou Cyanobactéries dans le cas des lichens), créant ainsi un organe (la mycorhize) ou un organisme (le lichen) mixte au sein duquel les deux partenaires échangent protection et nutriments. C’est aussi dans ces trois groupes que l’on trouve des espèces capables de former des sporocarpes, structures reproductives dans lesquelles sont mises en place les spores. Ces deux caractéristiques, symbioses et sporocarpes, sont absentes chez les autres groupes d’Eumycètes, suggérant qu’elles sont apparues chez le plus récent ancêtre commun des Mucoromycètes, Ascomycètes et Basidiomycètes.
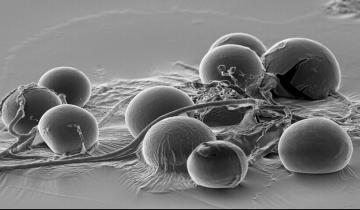
Le phylum des Mucoromycètes comprend un petit groupe d’Eumycètes d’importance écologique majeure : les Gloméromycètes (environ 160 espèces). Plus de 70 % de la flore terrestre établit une association symbiotique unique, appelée endomycorhize, avec une ou plusieurs espèces de Gloméromycètes. Le cycle de vie des Gloméromycètes est mal connu ; en particulier, l’existence d’une reproduction sexuée n’a pas été clairement démontrée, bien que le génome des gloméromycètes contienne des gènes de type sexuel. En revanche, les Gloméromycètes produisent des structures globuleuses riches en lipides, appelées spores, qui sont facilement observables à la loupe binoculaire ou au microscope.
Les Mucoromycètes comptent aussi des espèces saprophytes (comme celles du genre Rhizopus, moisissures communes), pathogènes (Mucor) ou endophytes de plantes. Un groupe récemment décrit, l’ordre des Endogonales, comprend même des espèces ectomycorhiziennes, ce qui indique que l’ectomycorhize est apparue plusieurs fois indépendamment dans l’évolution des Eumycètes. Gloméromycètes et Endogonales, grâce aux symbioses développées avec les plantes, sont très probablement impliqués dans la colonisation du milieu aérien par les premières plantes terrestres.
Les Ascomycètes sont le plus grand phylum d’Eumycètes, avec environ 32 000 espèces décrites. Ils rassemblent des organismes aussi divers que des moisissures de fromage (comme Penicillium roqueforti), des champignons comestibles (les morilles, les truffes…), des levures (par exemple Saccharomyces cerevisiae), des pathogènes (comme l’agent de la graphiose de l’orme, Ophiostoma ulmi) ou encore des lichens. Parmi les Ascomycètes mycorhiziens, on trouve des espèces ectomycorhiziennes (dont les truffes) mais aussi des espèces formant des mycorhizes éricoïdes avec des espèces d’Éricacées (groupe des Hélotiales).
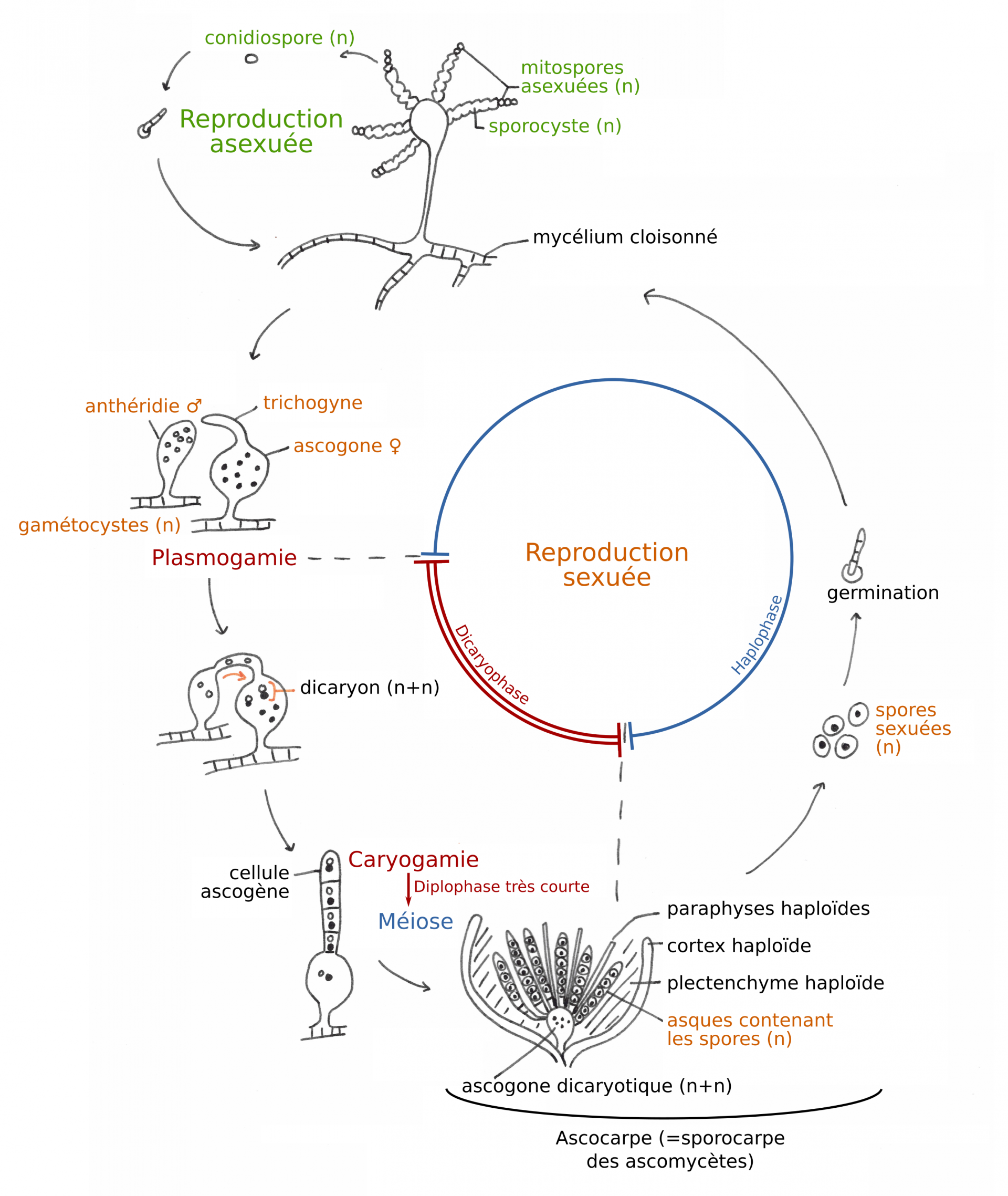
Les Ascomycètes tirent leur nom d’une particularité de leur cycle de vie (figure 7) : lors de la reproduction sexuée, deux mycéliums haploïdes fusionnent (plasmogamie), ainsi que leurs noyaux (caryogamie). Le noyau diploïde subit ensuite une méiose qui met en place les spores dans des asques, cellules reproductrices spécialisées à l’intérieur desquelles a lieu la méiose. Ces spores issues d’une reproduction sexuée (fécondation puis méiose) sont appelées ascospores, pour les différencier des conidiospores, qui sont issues du bourgeonnement de cellules plus ou moins spécialisées (reproduction asexuée).
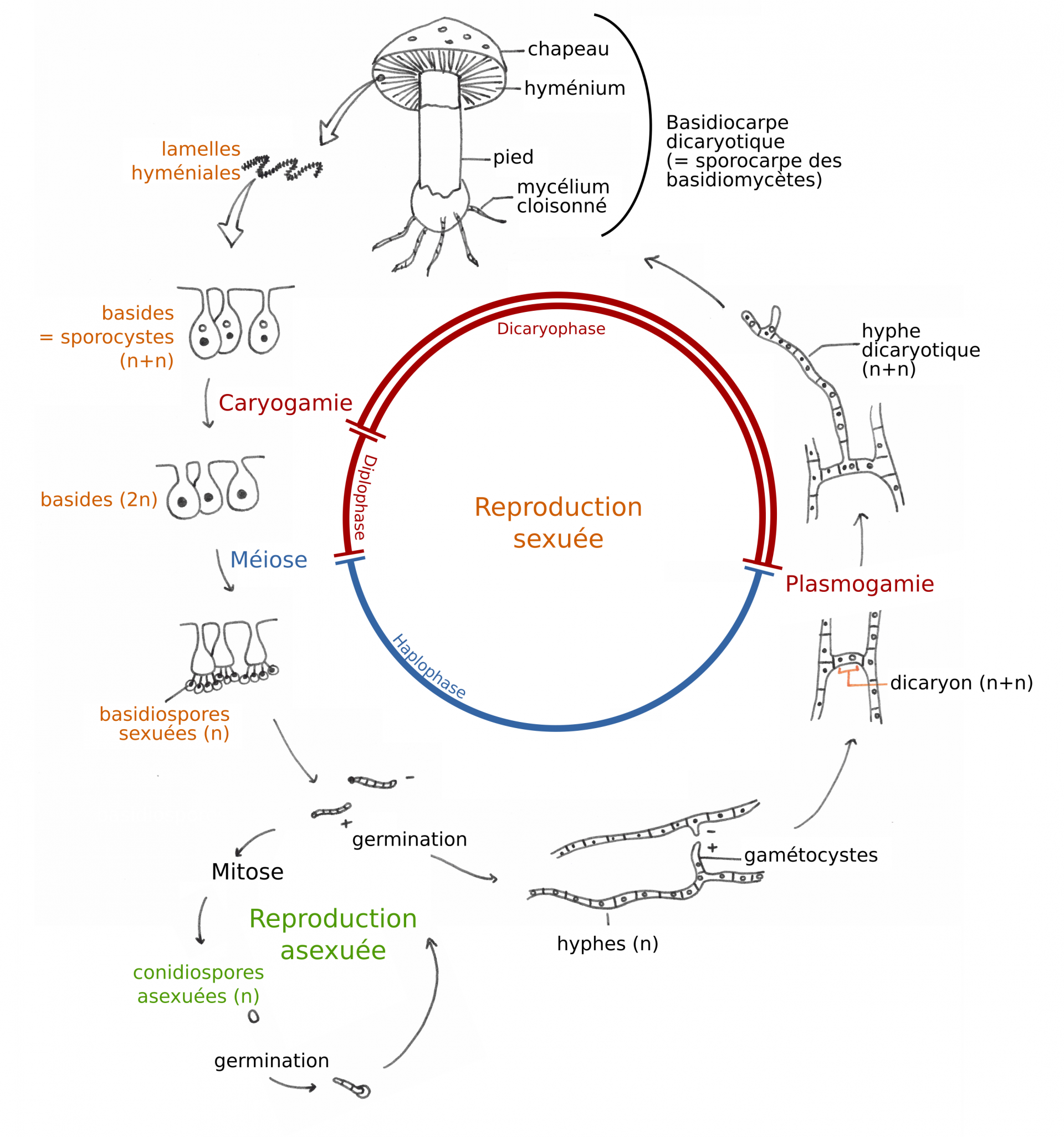
Les Basidiomycètes sont le deuxième plus grand phylum d’Eumycètes, avec environ 30 000 espèces décrites, dont un grand nombre d’espèces mycorhiziennes. On compte en particulier des espèces ectomycorhiziennes (comme les bolets, cèpes, amanites, etc.), des espèces formant des mycorhizes éricoïdes (Sérendipitacées) et des espèces formant des mycorhizes à pelotons avec des espèces d’Orchidées (Tulasnellacées, Sébacinales…).
Lors de la reproduction sexuée des Basidiomycètes, deux mycéliums haploïdes fusionnent (plasmogamie) mais leurs noyaux restent indépendants : le mycélium est alors dicaryotique. Ce mycélium dicaryotique met en place les basides, des cellules reproductrices spécialisées à l’intérieur desquelles a lieu la caryogamie (c’est-à-dire la fusion des noyaux du mycélium dicaryotique), puis la méiose, pour donner naissance à des basidiospores haploïdes. Les basides portant les basidiospores sont facilement observables en montant des lamelles de champignon de Paris (Agaricus bisporus) sous microscope. Les Basidiomycètes peuvent aussi se reproduire de manière asexuée en produisant des conidiospores par mitose.
NB : les cycles de vie présentés ici sont ceux d’espèces bien connues, mais de nombreux mystères restent encore à élucider car toutes les espèces ne se reproduisent pas exactement de la même manière (voir par exemple Les secrets de la reproduction de la Truffe du Périgord bientôt percés à jour ?). Néanmoins, les grandes caractéristiques de ces cycles, telles que l’alternance des phases haploïde, diploïde et/ou dicaryotique, sont conservées au sein de chaque phylum.



{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}