

Le cycle de vie des lis (espèces du genre Lilium) est un cycle typique des Embryophytes, c'est-à-dire où une génération sporophytique, diploïde et produisant les spores, alterne avec une génération gamétophytique, haploïde et produisant des gamètes. Plus spécifiquement, ce cycle permet de mettre en évidence des caractéristiques de la reproduction des Angiospermes : fleur, carpelle, fruit, double fécondation… Les structures associées à la reproduction sexuée des lis présentent l'avantage de pouvoir s'observer facilement à la loupe binoculaire et au microscope.
Les Angiospermes dominent la plupart des écosystèmes terrestres et constituent à ce titre le premier groupe en termes de biomasse (au moins 100 fois plus que celle de tous les animaux réunis !). C’est également, après les Insectes, le second taxon d’êtres vivants pluricellulaires concernant le nombre d’espèces (respectivement 1 million et 300 000 espèces connues). Cette biomasse et cette diversité témoignent d’un succès évolutif certain !
L’appareil végétatif des Angiospermes est globalement similaire à celui des Acrogymnospermes (plantes à graines nues actuelles, regroupant notamment les pins, sapins, cyprès, cycas, ginkgos…). Leur appareil végétatif ne semble donc pas, à première vue, la raison de ce succès évolutif. La grande nouveauté se situe plutôt au niveau de leur système reproducteur : la fleur, qui contient des carpelles qui se transforment en fruits (principale synapomorphie du groupe). Aussi, nous chercherons à répondre dans cet article aux questions suivantes : comment se déroule la reproduction sexuée des Angiospermes ? En quoi est-elle similaire à celle des autres Embryophytes et quelles sont les différences ?
Pour comprendre les grandes lignes de la reproduction des Angiospermes, nous allons nous intéresser à la reproduction des lis (ou lys, genre Lilium). Les fleurs de lis sont particulièrement faciles à étudier du fait de leurs grandes dimensions, permettant de réaliser aisément coupes et dissections florales. Elles ont de fait beaucoup été utilisées pour des coupes histologiques qui ont historiquement permis de comprendre la reproduction des Angiospermes. Par ailleurs, certaines espèces et variétés sont cultivées pour l’ornement et sont à ce titre souvent disponibles tout au long de l’année chez les fleuristes.
La reproduction sexuée du lis permet de générer de nouveaux individus, génétiquement différents de leurs parents. Cette diversité est permise par les brassages génétiques réalisés par la méiose et la fécondation (étapes caractéristiques de la reproduction sexuée des Eucaryotes). Chez les Angiospermes, les individus feuillés sont diploïdes1. À la différence des Animaux, où la méiose produit des gamètes haploïdes qui fusionnent directement pour former une cellule œuf diploïde (le zygote), chez les Angiospermes, la méiose produit des spores haploïdes, mâles ou femelles selon qu’elles sont produites par des organes mâles ou femelles. Ces spores sont produites par les individus feuillés que l’on qualifie ainsi de sporophytes (Figures 1 et 2). Ces spores haploïdes ne sont pas dispersées dans le milieu extérieur (contrairement à ce qui se produit chez les mousses et les fougères) et produisent, par mitose, des gamétophytes mâles et femelles. Le gamétophyte mâle, le grain de pollen, produit deux gamètes mâles. Le gamétophyte femelle, le sac embryonnaire, produit un gamète femelle. La fécondation de celui-ci par un gamète mâle produit un nouveau sporophyte diploïde. Le cycle de vie des Angiospermes comprend donc deux générations, l’une diploïde (le sporophyte), l’autre haploïde (les gamétophytes, mâle et femelle), comme chez toutes les Embryophytes (plantes terrestres).
Pour comprendre la reproduction du lis, nous commencerons par étudier la fleur et la formation des gamétophytes dans les pièces fertiles (étamines et pistil). Nous pourrons ensuite décrire les processus qui permettent ultimement le rapprochement des gamètes et la fécondation. Enfin, nous verrons la structure des graines et des fruits, et leur développement à partir des ovules et des carpelles. Pour argumenter toutes les étapes, nous travaillerons à la fois à partir d’échantillons frais et de coupes à main levée facilement réalisables en travaux pratiques avec une classe, et à partir de préparations histologiques du commerce.
La fleur épanouie et la formation des gamétophytes dans les pièces fertiles
Une fleur est constituée de pièces florales stériles et fertiles disposées en verticilles sur un axe, le tout axillé par une bractée
Les fleurs de lis sont produites vers l’apex des pieds feuillés. Elles sont colorées, et organisées en inflorescence distincte du reste de la plante (ici on parle de grappe, car les pédoncules floraux et les entre-nœuds entre les fleurs sont allongés). Ceci permet d’être plus visible vis-à-vis des pollinisateurs.

Chez cette espèce ornementale, vendue chez les fleuristes, les fleurs sont regroupées en inflorescence terminale (de type grappe). Chaque fleur mesure ici environ 15 cm de diamètre.

A : base de l’appareil végétatif. B : inflorescence. 1. Extrémité des étamines correspondant à l’anthère. 2. Pistil, composé, du haut vers le bas sur le dessin, de l’ovaire, du style et du stigmate. La présence de trois lobes dans le stigmate, visible sur le détail de stigmate en bas à gauche du dessin du pistil, indique la présence de trois carpelles soudés dans le pistil. 3. Coupe transversale de pistil au niveau de l’ovaire, montrant la présence de trois loges renfermant des ovules ; chaque loge correspond à un carpelle. 4. Vue externe du fruit en cours de déhiscence. 5. Coupe transversale de fruit, montrant les graines contenues à l’intérieur (dans la partie supérieure du dessin). 6. Graine.
La fleur de lis est produite par un individu diploïde chez lequel a lieu la méiose. Cette division cellulaire produit des spores, haploïdes. Le plant de lis, comme il produit des spores, est appelé sporophyte.
L’observation attentive d’une fleur épanouie (Figure 1) permet déjà d’en décrire assez bien la structure. Pour comprendre cette dernière plus en détails, il est possible de réaliser une coupe longitudinale de fleur épanouie (Figure 3, schématisée Figure 4), une dissection florale (Figure 5) et une coupe transversale de bouton floral (Figures 6 et 7).
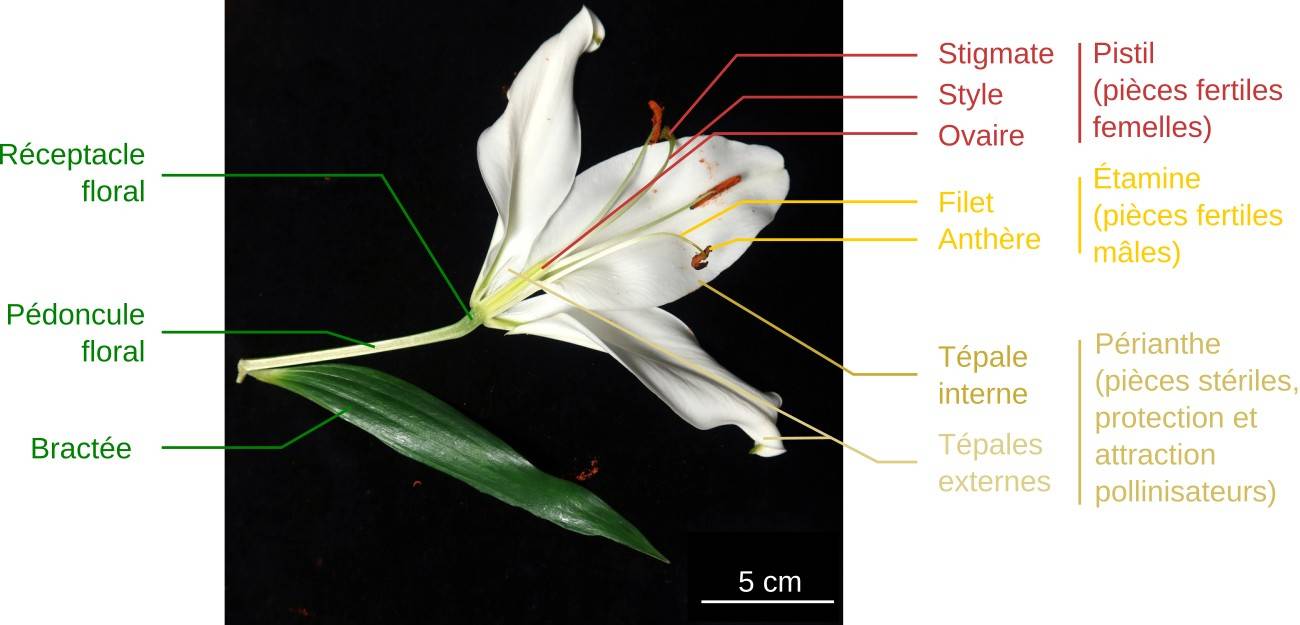
Cette coupe permet d’observer l’agencement spatial de l’ensemble des pièces d’une demi-fleur, avec à la base la bractée, le pédoncule et le réceptacle floral, et les verticilles de pièces florales (tépales externes et internes, étamines, pistil).
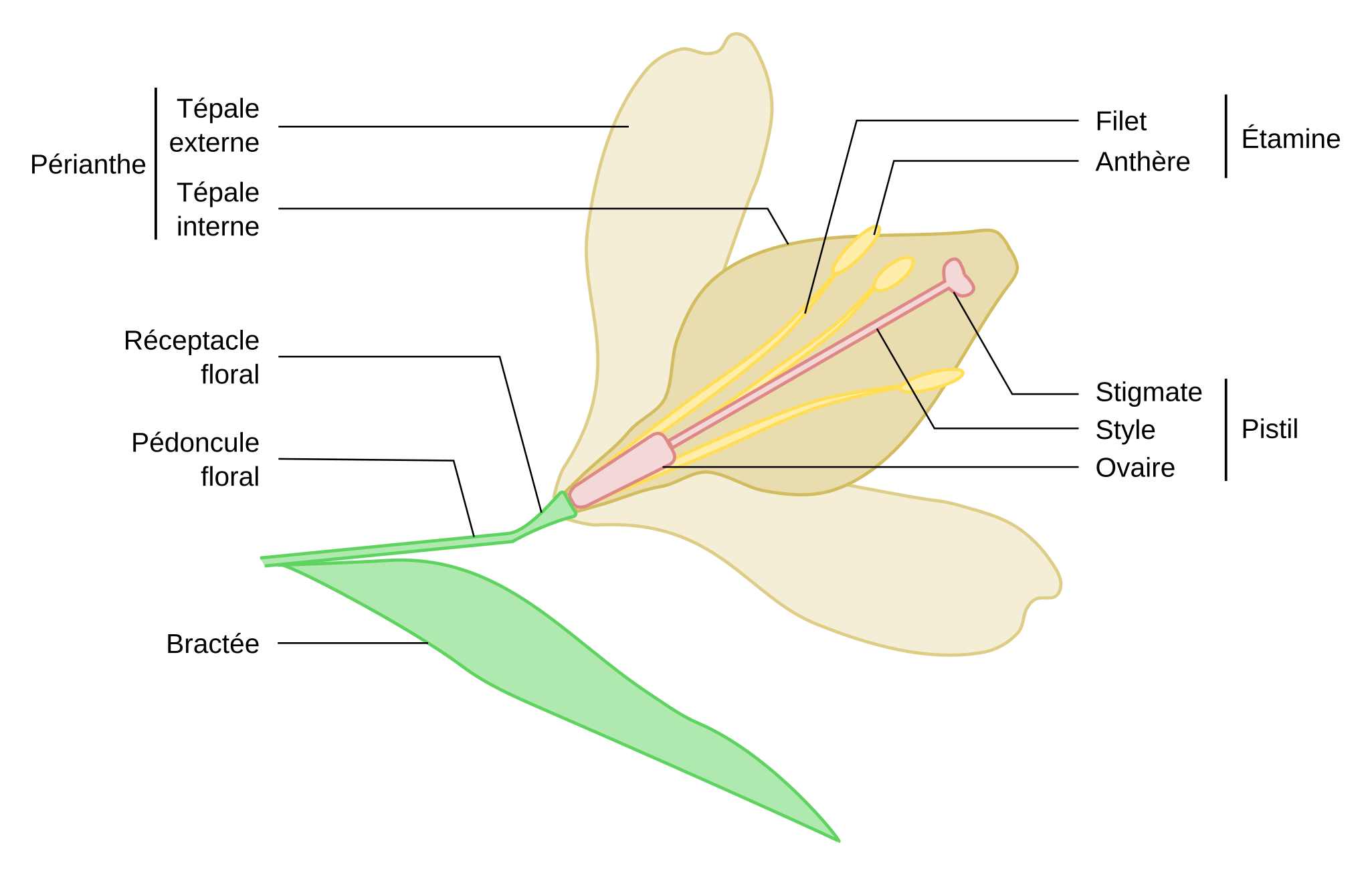
Les pièces florales forment quatre verticilles, c’est-à-dire quatre cercles concentriques de pièces similaires. Elles s’attachent au réceptacle floral, renflement de l’extrémité du pédoncule floral. Chez le lis, les pièces florales des deux premiers verticilles ne sont pas distinguables morphologiquement et sont appelées tépales. Le troisième verticille correspond à l’androcée, formé par les étamines. Le gynécée est formé de trois carpelles soudés en un seul pistil.
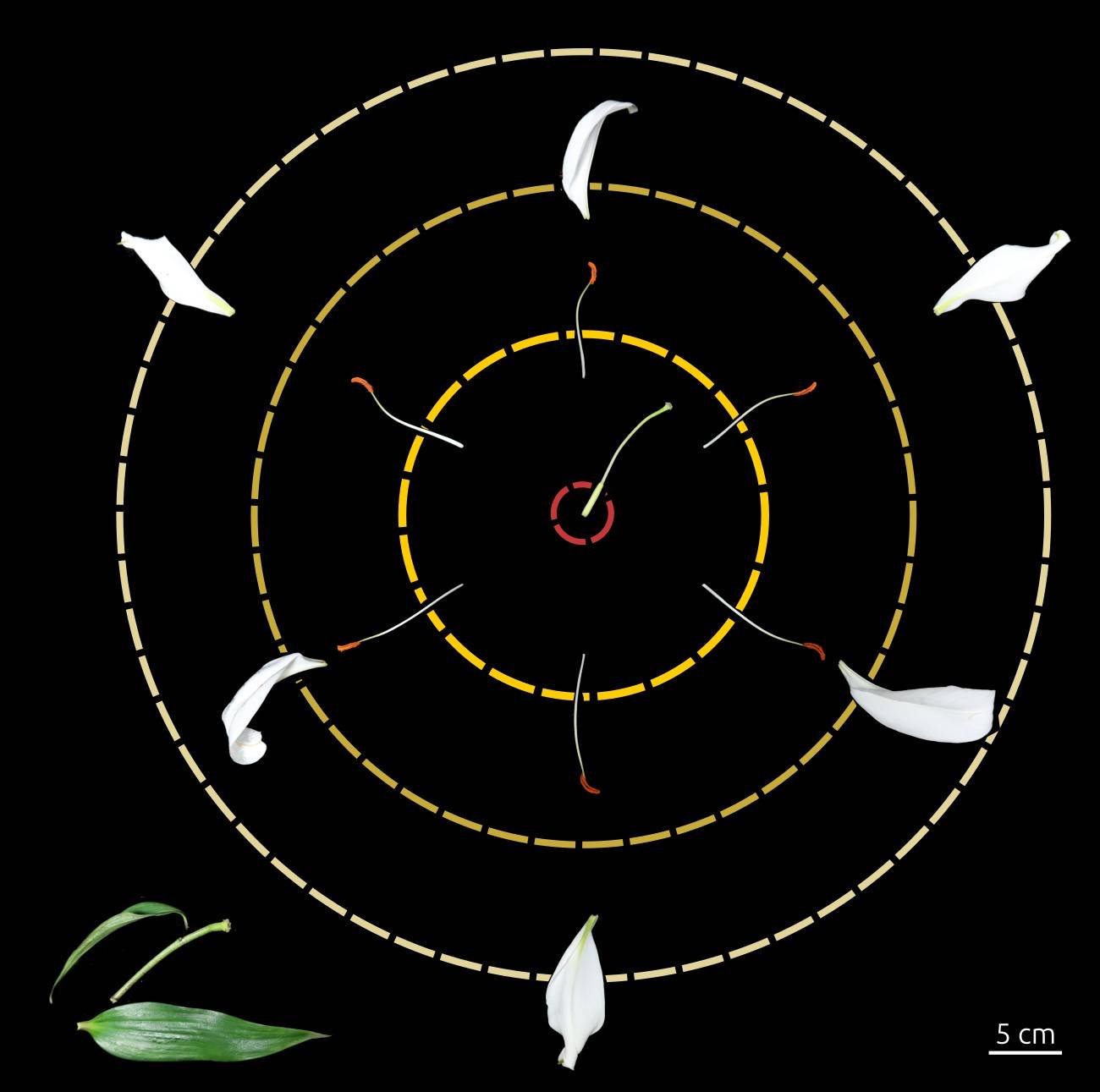
Dissection florale de lis avec les pièces de la base de la fleur en bas à gauche (bractée primaire et secondaire, pédoncule floral, réceptacle floral) et au centre les verticilles de pièces florales (de l’extérieur vers l’intérieur : 3 tépales externes ; 3 tépales internes ; 6 étamines ; 3 carpelles soudés formant un pistil).
Coupe transversale de bouton de fleur de lis (Lilium sp.) réalisée à main levée (à gauche) et schéma d’interprétation correspondant (à droite). On distingue bien les verticilles externes et internes de tépales (trois tépales chacun, l’ensemble formant le périanthe), le verticille de six étamines (qui forment l’androcée) et le pistil (ou gynécée), formé de trois carpelles soudés.
L’image résulte de l’assemblage de 11 clichés en microscopie optique. On distingue bien les verticilles externes et internes de tépales (trois tépales chacun, l’ensemble formant le périanthe), le verticille de six étamines (qui forment l’androcée) et le verticille de trois carpelles soudés pistil (ou gynécée).
La fleur de lis commence au niveau de la bractée (définie comme la feuille axillant une fleur), qui est ici grande, plane et photosynthétique. La fleur est située à l’aisselle de la feuille et de la tige, elle est portée par un pédoncule floral (il y a souvent une sorte de bractée secondaire sur le pédoncule floral des lis ornementaux, Figures 1 et 2). Sur le réceptacle floral (extrémité renflée du pédoncule) sont disposés plusieurs verticilles, c’est-à-dire plusieurs cercles concentriques de pièces florales :
- deux verticilles de 3 + 3 tépales (qui forment le périanthe), grandes structures foliacées colorées et odorantes. Très visibles et odorants, les tépales attirent les pollinisateurs après l’épanouissement de la fleur et, avant cela, protègent les pièces fertiles se formant à l’intérieur ;
- un verticille de six étamines, qui forment l’androcée. Chaque étamine est constituée d’un filet surmonté d’une anthère. Nous verrons juste après comment l’anthère permet la production et la libération des grains de pollen, gamétophytes mâles.
- un verticille de trois carpelles soudés par la longueur, qui forment le pistil, ou gynécée (le nombre de lobes du stigmate, ici trois, permet de supposer que le gynécée est formé de trois carpelles). Chaque carpelle est constitué, de la base vers l’apex :
- d’un ovaire, renflé et qui protège les ovules dans lesquels sont produits les gamétophytes femelles ;
- d’un style, allongé et creux, par lequel passent les tubes polliniques qui permettent la fécondation ;
- d’un stigmate, lobé et collant, qui reçoit et fait germer les grains de pollen.
Les études de biologie moléculaire ont permis de démontrer que les différentes pièces florales sont des feuilles modifiées portées par un rameau réduit spécialisé dans la reproduction (théorie de la métamorphose initialement proposée par Goethe en 1790).
Pour conclure, une fleur peut être définie morphologiquement comme un ensemble de pièces stériles et fertiles disposées en verticilles sur un axe court, rattaché au reste de la plante par un pédoncule floral (parfois très court). Selon les espèces, une bractée est présente ou non à la base du pédoncule. Les fleurs terminales n’ont jamais de bractée. Une fleur peut aussi être définie, de façon complémentaire, par sa fonction : il s’agit d'un système qui produit les gamétophytes mâles et femelles, et permet leur rapprochement. Nous allons maintenant nous intéresser à cette fonction.
Lames histologiques interactives
Les lames histologiques présentées dans cet article sont également disponibles, en version interactive (mise en évidence des structures au survol de la légende), sur le site internet des collections pédagogiques du département de biologie de l’École normale supérieure de Lyon. Le contenu de ce site est présenté dans l’article de Planet-Vie intitulé Lames histologiques, photographies de spécimens, modèles 3D… découvrez le site des collections de l'ENS Lyon !
Production des gamétophytes mâles : les grains de pollen
La partie fertile d’une étamine est l’anthère. Pour en décrire la structure, il faut réaliser des coupes dans le bouton floral avant l’épanouissement de la fleur. L’anthère comporte notamment des sacs polliniques protecteurs, où sont produits les grains de pollen (gamétophytes mâles).
Structure de l’anthère
En haut : coupe à main levée ; en bas : préparation histologique. La symétrie est bilatérale, avec quatre sacs polliniques reliés par un connectif.
L’anthère est formée de deux paires de sacs polliniques à l’intérieur desquels se forment les grains de pollens. À maturité, les sacs polliniques s’ouvrent au niveau de la zone de déhiscence entre chaque paire et libèrent le pollen. L’anthère est portée par le filet, le tout constituant l’étamine.
En coupe, la symétrie bilatérale de l’anthère évoque la silhouette d’un papillon. Chacune des quatre « ailes » est un sac pollinique (sporange mâle, voir l’encadré Quelques éléments de vocabulaire sur la reproduction des Angiospermes), qui protège la sporogenèse et la production des grains de pollen (gamétophytes mâles), avant de les libérer. Au centre, le connectif relie les sacs polliniques et transmet les nutriments apportés par un faisceau cribro-vasculaire.
Chaque sac pollinique est constitué de plusieurs couches de cellules (assises) jouant chacune un rôle différent :
- l’épiderme protège l’ensemble de la dessication ;
- l’assise mécanique présente une paroi lignifiée et agit comme un ressort au moment de l’épanouissement. Cela permet la déhiscence de l’anthère, c’est-à-dire son ouverture, et la présentation des grains de pollen aux pollinisateurs ;
- le tapis permet la nutrition des cellules participant aux différentes étapes de la formation des grains de pollen.
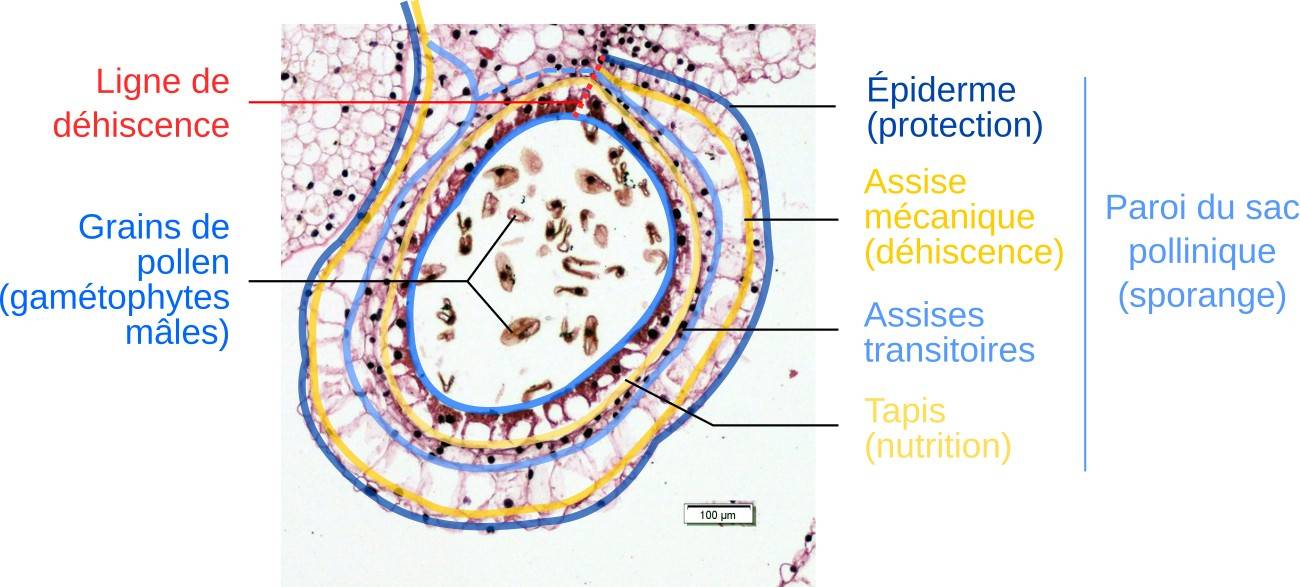
La paroi du sac est constituée de plusieurs assises qui assurent des fonctions différentes dans la protection et la formation des grains de pollen.
Quelques éléments de vocabulaire sur la reproduction des Angiospermes
Chez les Angiospermes, deux types de spores sont produites : on parle d’hétérosporie. Ces spores diffèrent par leur taille : les grosses spores sont qualifiées de mégaspores (ou macrospores) et les petites de microspores. Les mégaspores sont produites dans les mégasporanges, sur une mégasporophylle (le carpelle, une feuille modifiée), et donnent par mitoses les mégagamétophytes. Les microspores sont produites par les microsporanges, sur une microsporophylle (l’étamine, une autre feuille modifiée) et donnent par mitoses les microgamétophytes. Par définition, les structures impliquant un plus grand investissement énergétique sont qualifiées de femelles, et les structures impliquant un plus faible investissement énergétique de mâles. Aussi, pour simplifier le jargon pour des lecteurs non habitués, nous avons fait le choix dans le présent article de simplifier le vocabulaire avec mégaspore = spore femelle ; mégasporange = sporange femelle ; mégagamétophyte = gamétophyte femelle ; microspore = spore mâle ; microsporange = sporange mâle ; microgamétophyte = gamétophyte mâle.
Structure des grains de pollen
Les grains de pollen sont facilement observables en les prélevant sur les anthères des fleurs épanouies et en les déposant entre lame et lamelle. Il s’agit de structures d’environ 100 µm, ovoïdes et présentant une paroi ornementée. Les gouttelettes jaunes à la surface sont collantes (c’est le pollenkitt). Associées aux reliefs en réseau, elles permettent aux grains de pollen de mieux adhérer aux pollinisateurs. Une fente, l’aperture, permet la germination de la cellule végétative et la formation du tube pollinique lorsque le pollen rentre en contact avec le stigmate et se réhydrate.

La paroi a un aspect réticulé dû à la présence d’épaississements plus marqués à certains endroits. On distingue une zone plus uniforme formant un sillon : c’est l’aperture, qui joue un rôle dans les échanges d’eau avec l’extérieur et dans la germination du pollen.
Des coupes fines au microtome sur des préparations fixées de pollen permettent d’observer le contenu. La structure peut également être appréhendée en utilisant des colorants nucléaires sur du pollen frais.
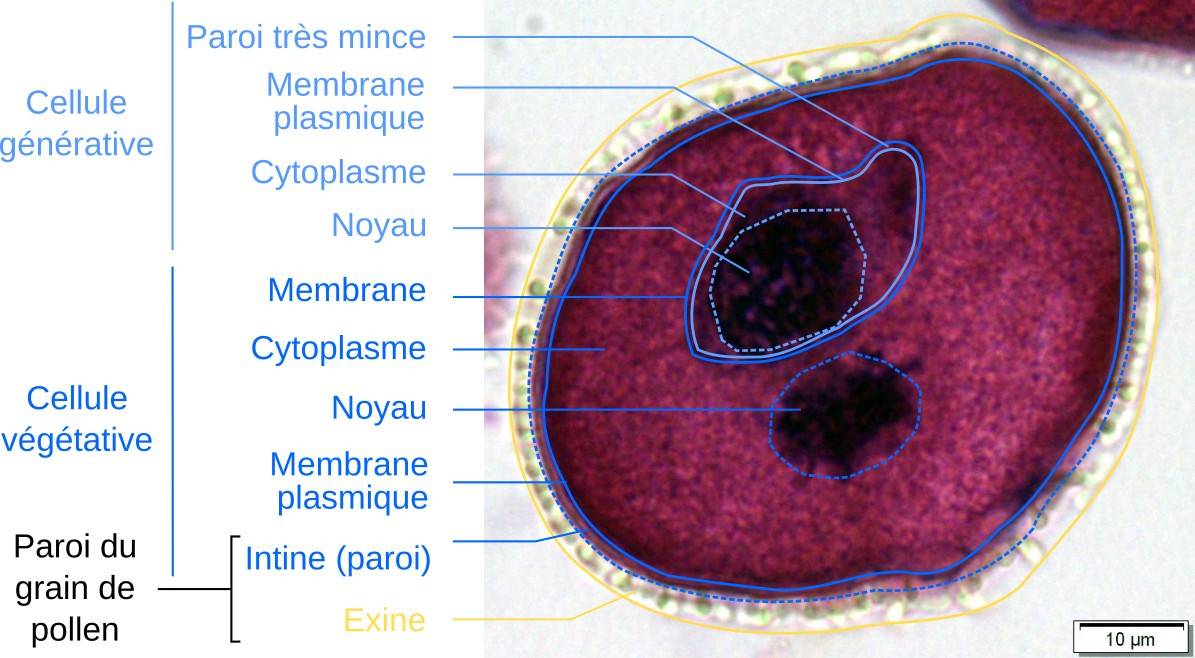
L'observation en microscopie optique permet de distinguer la cellule générative, incluse dans la cellule végétative. Le grain de pollen est entouré d’une paroi.
Le grain de pollen est le gamétophyte mâle. Il est constitué de seulement deux cellules haploïdes contenues dans une paroi complexe et très résistante. Le gamétophyte mâle, tout comme le gamétophyte femelle (voir plus bas), est donc très réduit, comme chez toutes les plantes à graines actuelles mais contrairement à ce que l’on peut observer chez les Embryophytes dites « à spores libres » (mousses et fougères). La cellule végétative occupe la majorité du volume. Après la pollinisation, elle permettra la formation du tube pollinique, qui transporte les gamètes. À l’intérieur de la cellule végétative se trouve la cellule générative. Elle se divisera encore une fois par mitose et formera deux gamètes mâles.
Les cellules sont très déshydratées, et contiennent des réserves (lipides, glucides notamment), ce qui est typique de formes de vie ralentie. La partie interne (intine) de la paroi protectrice du grain de pollen est pectocellulosique, et la partie externe (exine) est composée de sporopollénine. Cette dernière est le composé biologique le plus résistant connu, et assure à ce titre une protection mécanique et chimique. Elle assure également une protection vis-à-vis des ultra-violets.
Le grain de pollen (gamétophyte mâle) est formé par méiose suivie de deux mitoses
Des coupes à différents moments de la formation des anthères permettent de décrire le processus de formation des grains de pollen. Les figures 13 et 14 témoignent d’étapes importantes de ces processus.
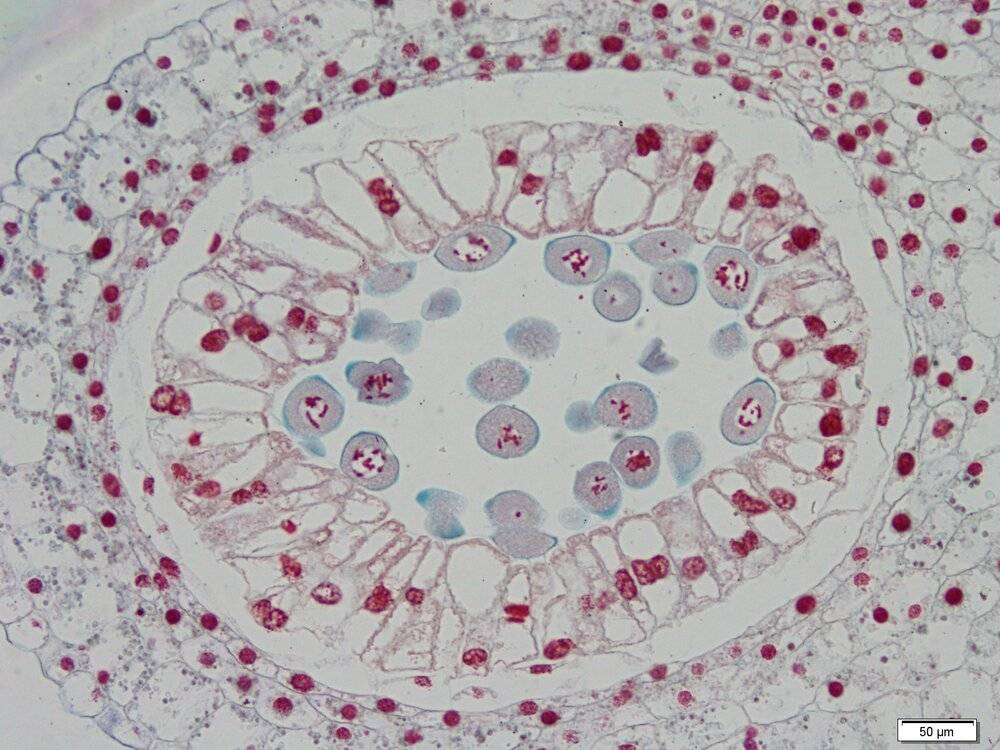
Le tissu au centre du tapis est constitué de cellules mères de spores qui prolifèrent et s’entourent d’une paroi formée de callose (épaississements en bleu) (ici en prophase de mitose).
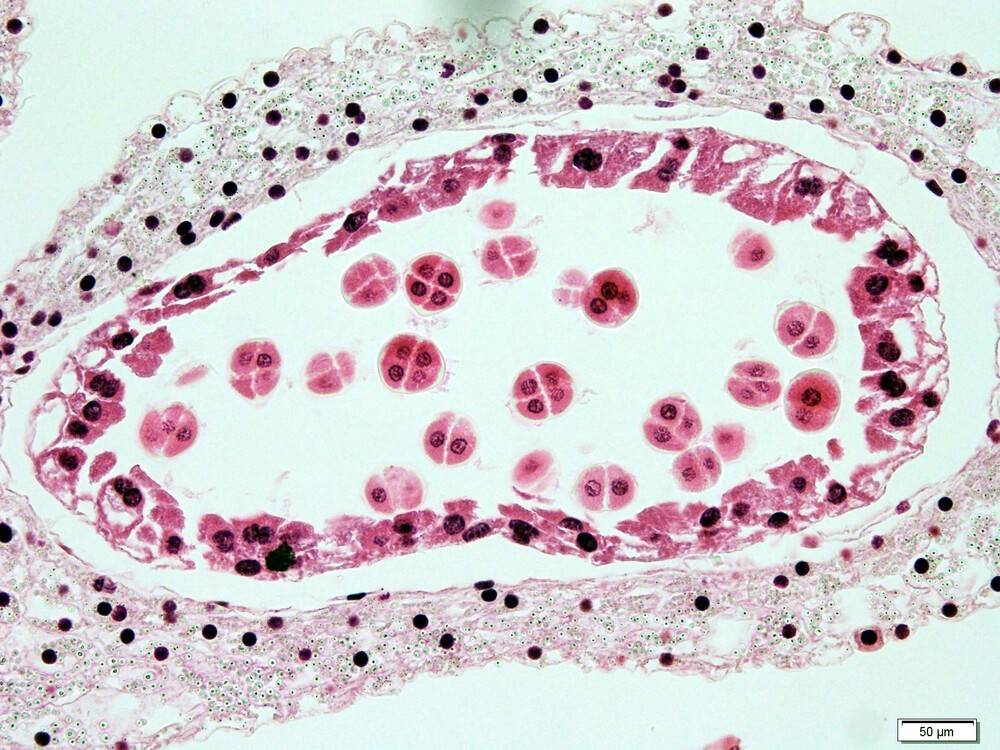
Les cellules regroupées par quatre sont le résultat typique d’une méiose. Elles sont contenues dans une paroi de callose, qui est ensuite dégradée et permet à chaque cellule de former indépendamment un grain de pollen.
La formation des grains de pollen commence par la prolifération de cellules mères de spores, contenues dans les sacs polliniques. Ces cellules mères des spores s’entourent d’une paroi de callose et subissent une méiose qui produit quatre spores mâles haploïdes, processus qui marque le passage dans la génération gamétophytique du cycle de vie. Les spores se forment à l’intérieur de la paroi de callose de la cellule mère et sont également séparées par des parois de callose formées durant la méiose, formant une structure appelée tétrade.
La paroi de callose est ensuite dégradée et chaque spore se divise par mitose, pour donner la cellule végétative et la cellule générative, cette dernière étant incluse dans le cytoplasme de la cellule végétative. En plus de leur rôle nutritif déjà présenté, les cellules du tapis s’immiscent entre les spores et synthétisent la sporopollénine de l’exine. La cellule végétative produit ensuite la paroi d’intine.
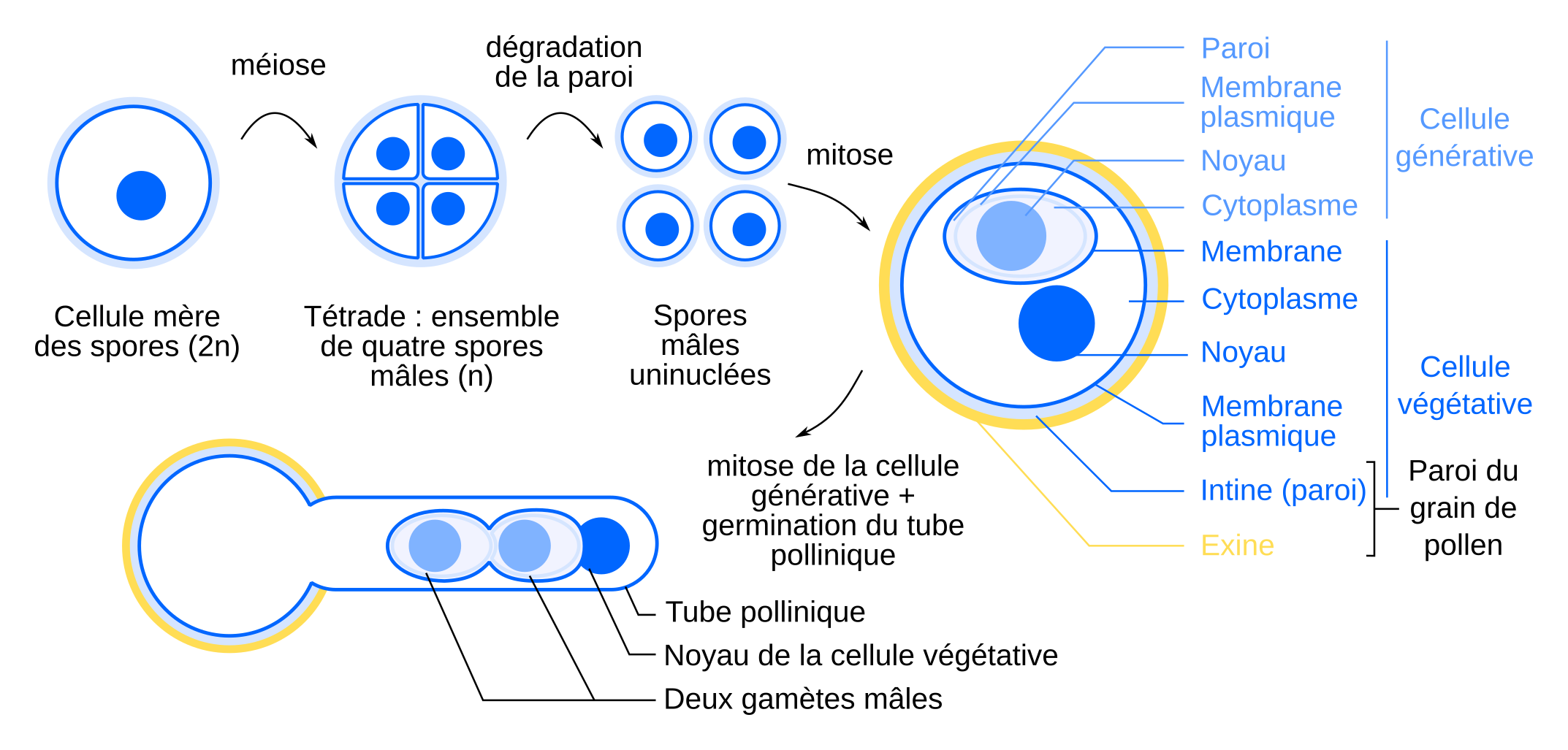
Les grains de pollen sont produits dans les sacs polliniques des anthères. Ces sacs polliniques contiennent les cellules mères des spores, diploïdes. Lorsqu’une cellule mère des spores entre en méiose, elle produit quatre spores mâles haploïdes contenues dans une même paroi. Après dégradation de la paroi, chaque spore subit une mitose aboutissant à un grain de pollen bicellulaire, composé d’une cellule végétative à l’intérieur de laquelle est incluse une cellule générative. Notez qu’alors que la plupart des schémas de grain de pollen représentent une seule membrane entourant la cellule générative, cette cellule est en réalité entourée de sa membrane plasmique, de sa paroi et d'une membrane issue de la cellule végétative 12. Cette cellule générative produit deux gamètes mâles par mitose. Ceux-ci sont acheminés jusqu’à l’ovule grâce à la formation d’un tube pollinique par la cellule végétative.
Production des gamétophytes femelles : les sacs embryonnaires
La partie fertile du pistil est son ovaire. Sa structure à différentes échelles microscopiques peut être observée en réalisant des coupes transversales. Nous allons voir que cela permet de distinguer un ensemble de structures emboîtées, avec des carpelles et des ovules protecteurs, dans lesquels sont produits les sacs embryonnaires (gamétophytes femelles).
L’ovaire est constitué de carpelles qui protègent les ovules
L’ovaire forme une sorte d’hexagone lobé et plus ou moins régulier selon les espèces de lis. Il est constitué de trois lames (les carpelles) repliées sur elles-mêmes et soudées l’une à l’autre. Cela constitue des enveloppes complètement closes renfermant des structures globuleuses, les ovules.
Durant leur formation, les ovules sont protégés de la dessication et des agressions physiques (notamment par les consommateurs : herbivores, frugivores, parasites…) par l’épiderme et l’épaisse paroi des carpelles. Ces mêmes structures protègent, après la fécondation, les graines issues des ovules. Durant l'ensemble du développement des ovules et des graines, des faisceaux cribro-vasculaires apportent les nutriments nécessaires.
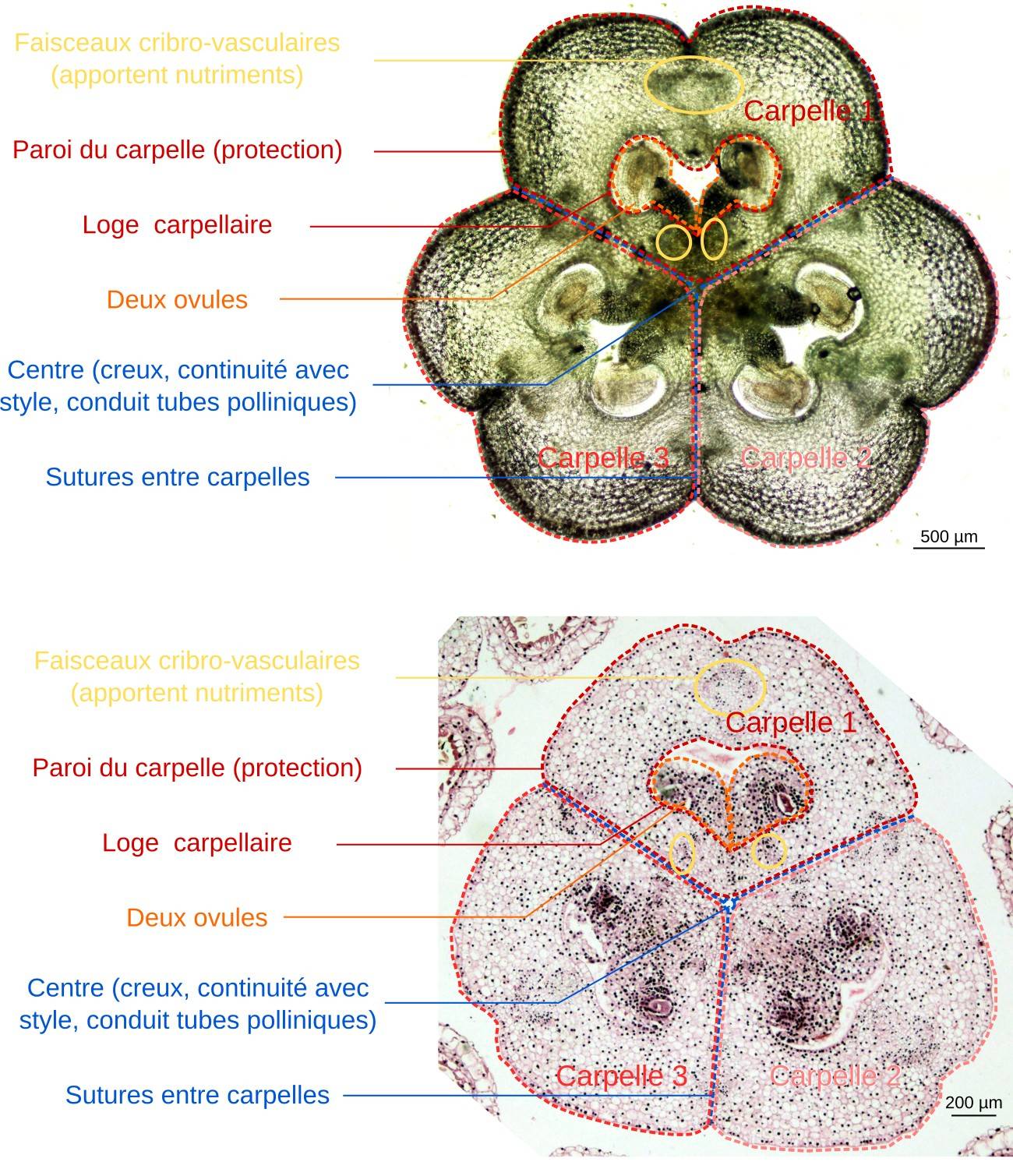
En haut : coupe à main levée ; en bas : préparation histologique. Ces coupes permettent de décrire la structure des carpelles, leur agencement et leur soudure, ainsi que d’observer en détail des ovules.
L’ovaire de lis est formé de trois carpelles soudés. Chaque carpelle contient une loge carpellaire le long de laquelle sont disposés les ovules, par groupes de deux. Les ovules sont reliés au carpelle par l’intermédiaire du funicule.
Les ovules sont des structures protectrices de la sporogenèse et du développement du gamétophyte femelle
Les ovules des Angiospermes sont des structures pluricellulaires à l’intérieur desquels est produit le gamète femelle, l’oosphère. Malgré l’emploi du même nom, les ovules des Angiospermes ne doivent donc pas être confondus avec les ovules des Métazoaires, qui sont quant à eux des gamètes, c’est-à-dire des cellules haploïdes directement impliquées dans la fécondation.
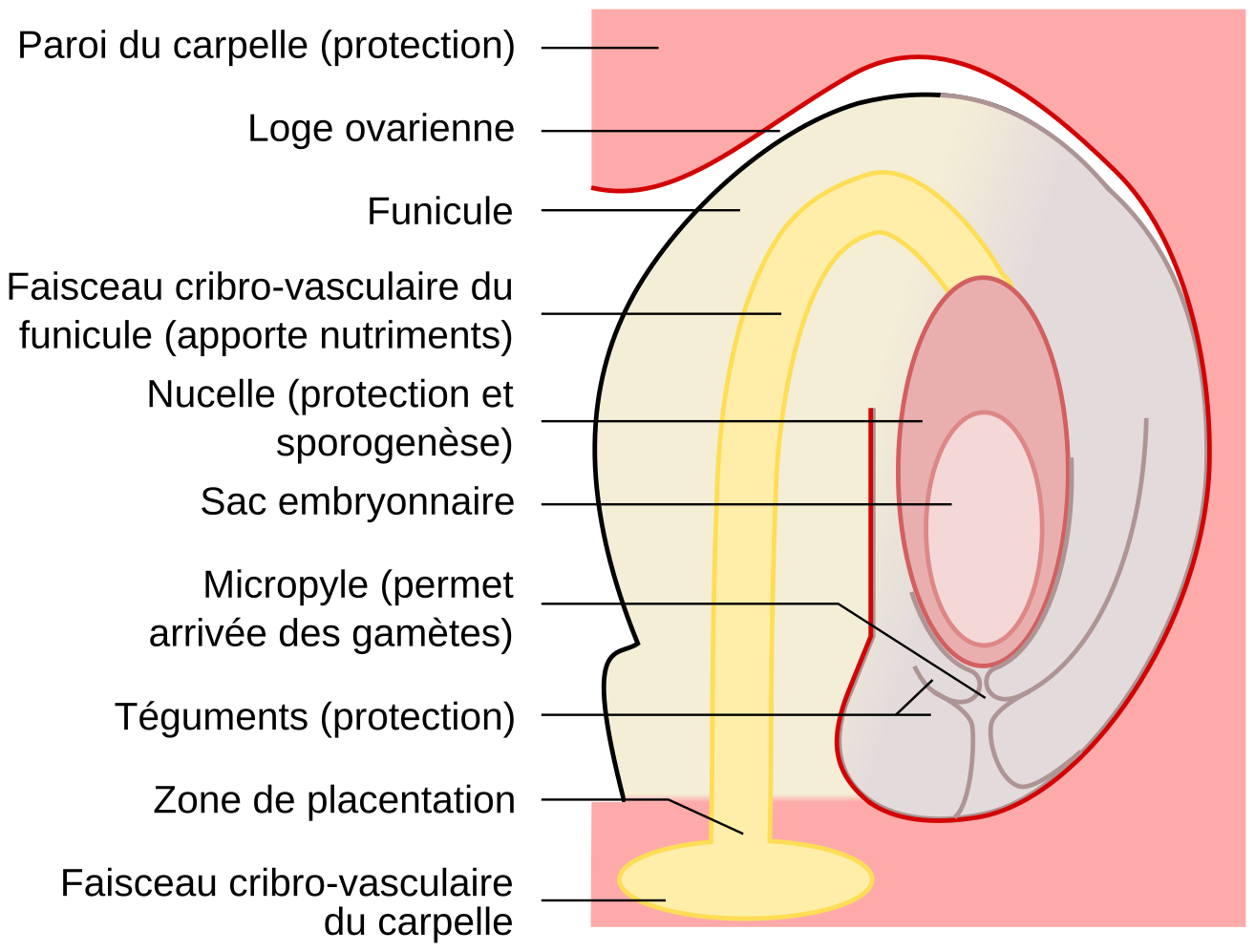
L’observation d’un ovule de lis permet de distinguer, de l’extérieur vers l’intérieur :
- deux téguments protecteurs (caractéristique des Angiospermes, les graines des Gymnospermes n’en possédant qu’un seul). Ils sont discontinus au niveau du micropyle, qui permet, à terme, l’arrivée des gamètes mâles ;
- le nucelle ou sporange femelle, un tissu siège de la sporogenèse, puis du développement du gamétophyte femelle ;
- le sac embryonnaire, qui correspond au gamétophyte femelle. Il est très réduit, constitué de seulement sept cellules, avec huit noyaux chez le lis, comme chez de nombreuses espèces d’Angiospermes1. L’oosphère, le gamète femelle (haploïde), se trouve du côté du micropyle avec deux synergides. Une cellule centrale comporte deux noyaux polaires. Trois antipodes se trouvent à l’opposé du micropyle (côté chalaze). Nous verrons que la ploïdie de l’ensemble est complexe chez le lis.
Le funicule relie le corps de l’ovule (au niveau de la chalaze) au carpelle (au niveau du placenta). Un faisceau cribro-vasculaire passe au travers et apporte les nutriments.
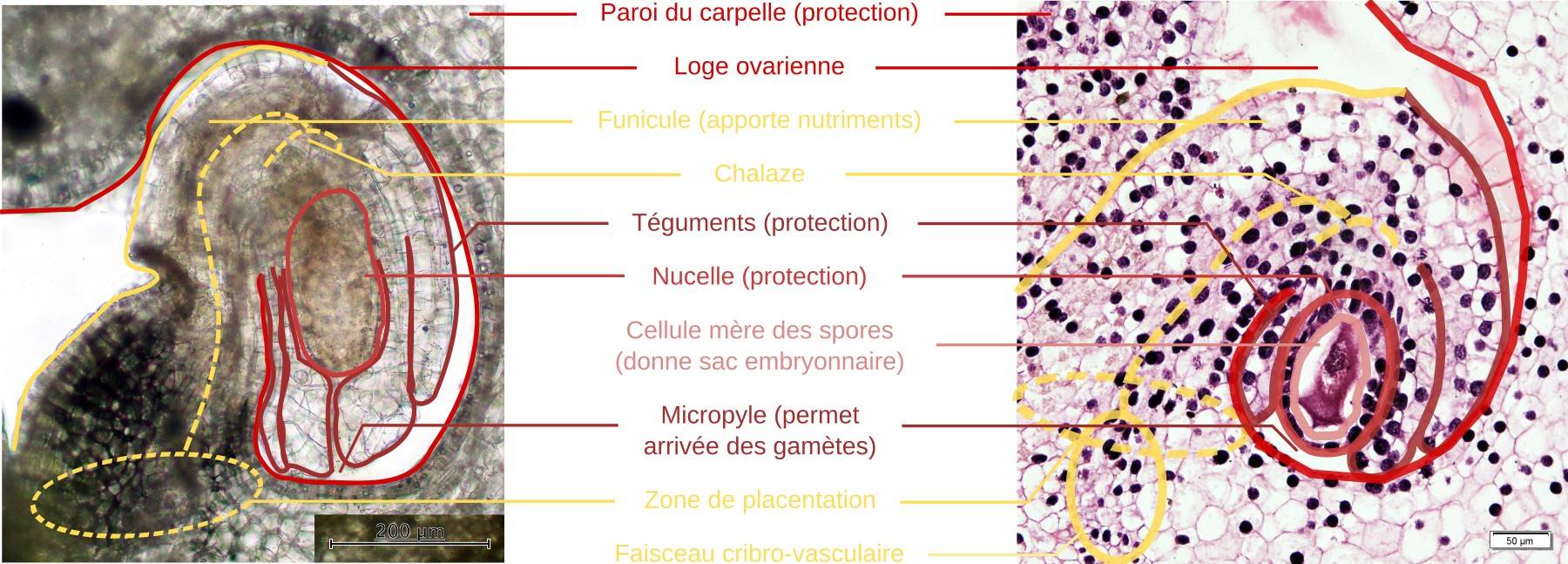
À gauche : coupe à main levée ; à droite : préparation histologique (stade avant méiose, toutes les cellules visibles sont diploïdes et produites par le sporophyte). On distingue bien les deux téguments protecteurs, discontinus au niveau du micropyle, le nucelle (sporange femelle) et la structure qui assure la nutrition de l’ensemble (placenta, funicule).
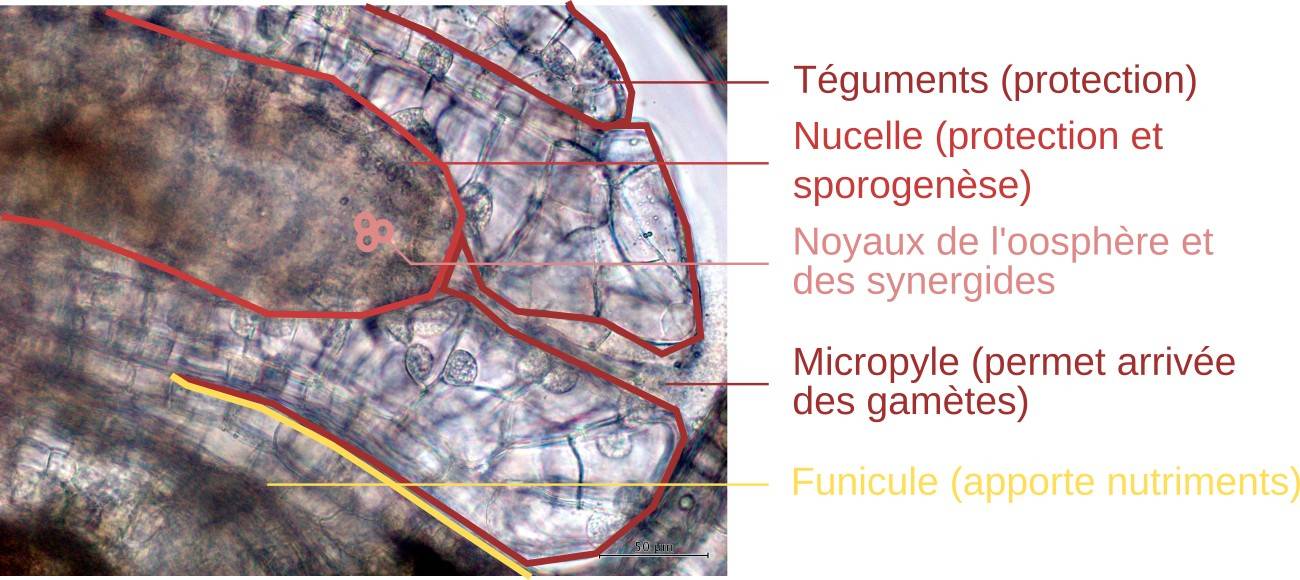
Les ovules sont situés dans les ovaires. Ils sont reliés à la paroi du carpelle par le funicule. L’ovule est constitué, de l’extérieur vers l’intérieur, de deux téguments protecteurs, du nucelle et du sac embryonnaire qui lui-même contient le gamète femelle. Après la double fécondation, l’ovule se transforme en graine.
Le sac embryonnaire (gamétophyte femelle) est formé par méiose suivie de deux mitoses irrégulières
Des coupes dans le pistil à différents moments de sa formation permettent de comprendre les processus de formation du sac embryonnaire. La figure 21 témoigne de la méiose, une étape importante1.
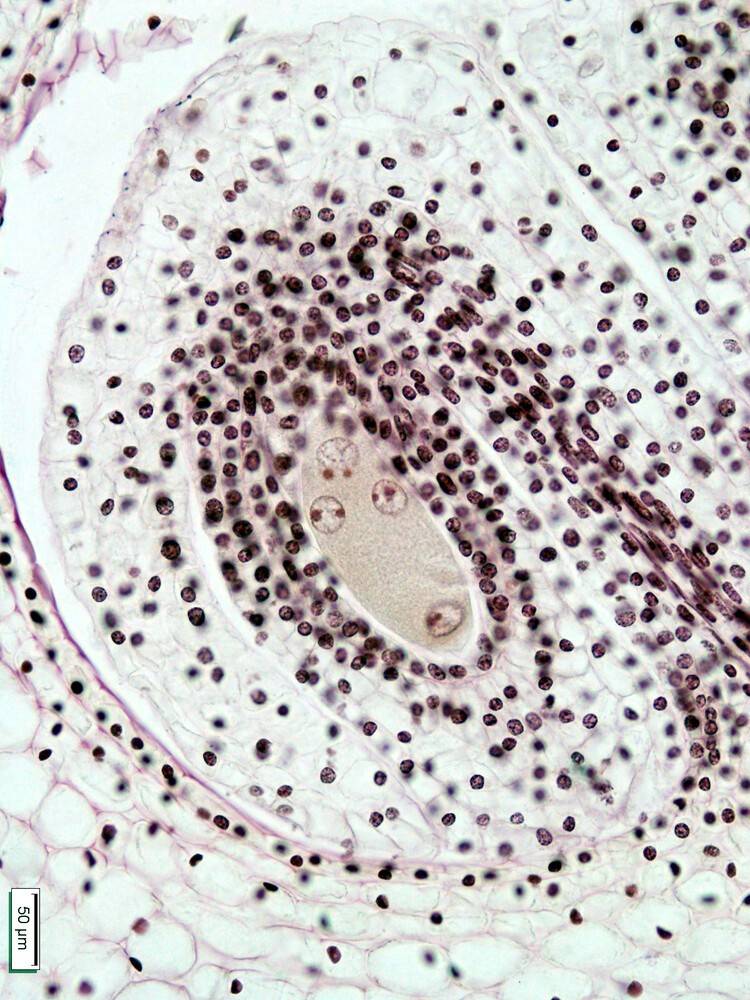
À l’issue de la méiose, les 4 noyaux, tous haploïdes à ce stade, ne sont pas séparés par des membranes.
La formation du sac embryonnaire commence par la méiose de la cellule mère des spores (visible Figure 18). Quatre noyaux sporiques sont formés dans une cellule, sans cloisonnement membranaire, la cytocinèse se produisant ici après la caryocinèse. La méiose marque le passage à la phase haploïde (gamétophytique) du cycle de vie.
Un des noyaux reste indépendant tandis que les trois autres fusionnent en un noyau triploïde1. Chacun de ces deux noyaux se divise deux fois par mitose. Les cloisonnements cellulaires (cytocinèse) n’interviennent qu’à la fin des divisions, et de façon irrégulière (7 cellules de dimensions inégales formées pour 8 noyaux). Le noyau haploïde, côté micropyle, est à l’origine de l’oosphère (gamète femelle), des deux synergides et d’un des noyaux polaires. Le noyau triploïde est à l’origine des trois antipodes et de l’autre noyau polaire.
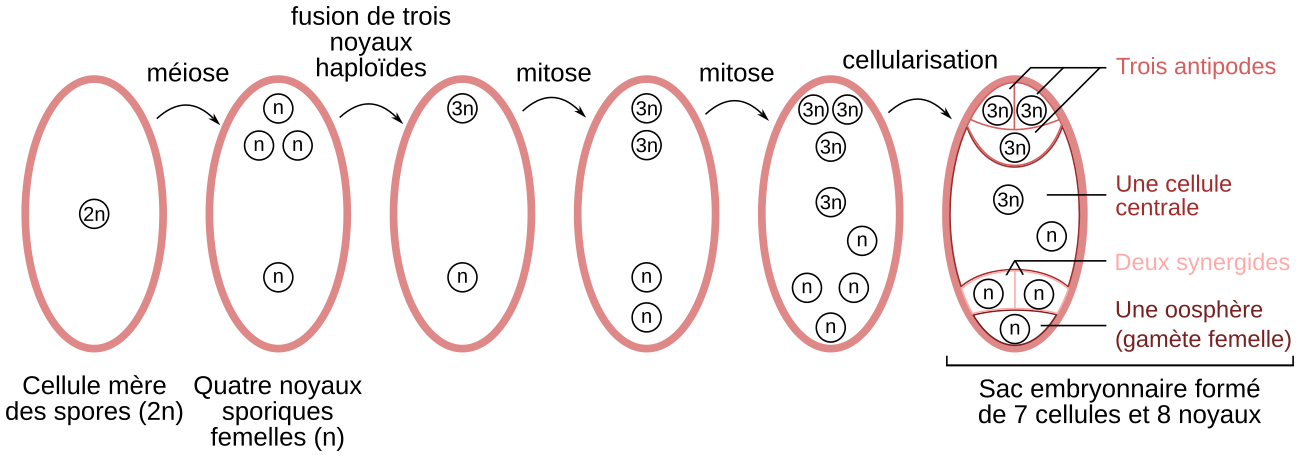
Le sac embryonnaire se forme à partir d’une cellule mère des spores, contenue dans le nucelle. Après méiose, quatre noyaux haploïdes sont produits au sein d’une même cellule (il n’y a pas de cytocinèse après les divisions nucléaires). Trois des noyaux haploïdes fusionnent en un noyau triploïde. Après deux mitoses, toujours sans cytocinèse, huit noyaux sont obtenus. Enfin, l’ensemble se cellularise pour former un sac embryonnaire constitué de sept cellules : trois antipodes à noyau triploïde, une cellule centrale contenant un noyau triploïde et un noyau haploïde, deux synergides haploïdes et une oosphère (gamète femelle) haploïde. Notez que les modes de formation des sacs embryonnaires sont très diversifiés selon les Angiospermes et que le cas exposé ici pour le lis n’est donc qu’un exemple parmi d’autres.
De la pollinisation à la fécondation
La pollinisation : transport des grains de pollen des anthères aux stigmates
À l’épanouissement de la fleur, les sacs polliniques des anthères s’ouvrent par déformation de l’assise mécanique et rupture de la fente de déhiscence. Cela libère les grains de pollen (qui forment une poudre orangée sur n’importe quel objet en contact avec les anthères, ou sur une surface située en dessous). La quantité de grains de pollen est extrêmement variable entre les espèces, de quelques dizaines à plusieurs milliers.


Des insectes (en particulier les Lépidoptères Sphingidés, famille des sphinx) viennent boire du nectar produit à la base des tépales. Au passage, ils prélèvent involontairement du pollen sur leur corps en touchant les étamines. Le pollen prélevé peut être déposé, tout aussi involontairement, par contact avec le stigmate d’une fleur (la même, une autre du même plant, ou sur un autre plant). C’est la pollinisation, soit le transport du pollen d’une anthère à un stigmate. La pollinisation par les insectes est la plus commune chez les Angiospermes, et se base sur une relation mutualiste entre les plantes et les pollinisateurs : la plante bénéficie du transport de son information génétique qui permet sa reproduction sexuée, tandis que l’insecte trouve de la nourriture.

Une deuxième modalité de pollinisation chez le lis fait intervenir le vent et la gravité. Ce transport n’est pas directionnel, mais une fleur de lis produit d’importantes quantités de pollen et présente un stigmate avec une surface assez étendue. La probabilité d’une pollinisation réussie reste importante. Ce type de pollinisation permet notamment une pollinisation de secours, au cas où la fleur n’ait pas reçu de visiteurs.
En horticulture, les humains participent aussi à la pollinisation en utilisant volontairement un objet pour prélever et déposer le pollen.
La pollinisation permet, quel qu’en soit le mode, le rapprochement des gamétophytes. Cependant, à ce stade, il reste aux gamètes mâles plusieurs centimètres à parcourir pour atteindre le sac embryonnaire. Nous allons voir maintenant que la suite se passe dans le style.
Le tube pollinique croît et transporte les gamètes mâles jusqu’au sac embryonnaire
Réaliser des coupes longitudinales dans des styles quelques heures après la pollinisation permet de décrire le mécanisme qui permet d’aboutir à la fécondation. On observe en effet que les grains de pollen présentent de longues protubérances, les tubes polliniques, qui parcourent le style vers l’ovaire.
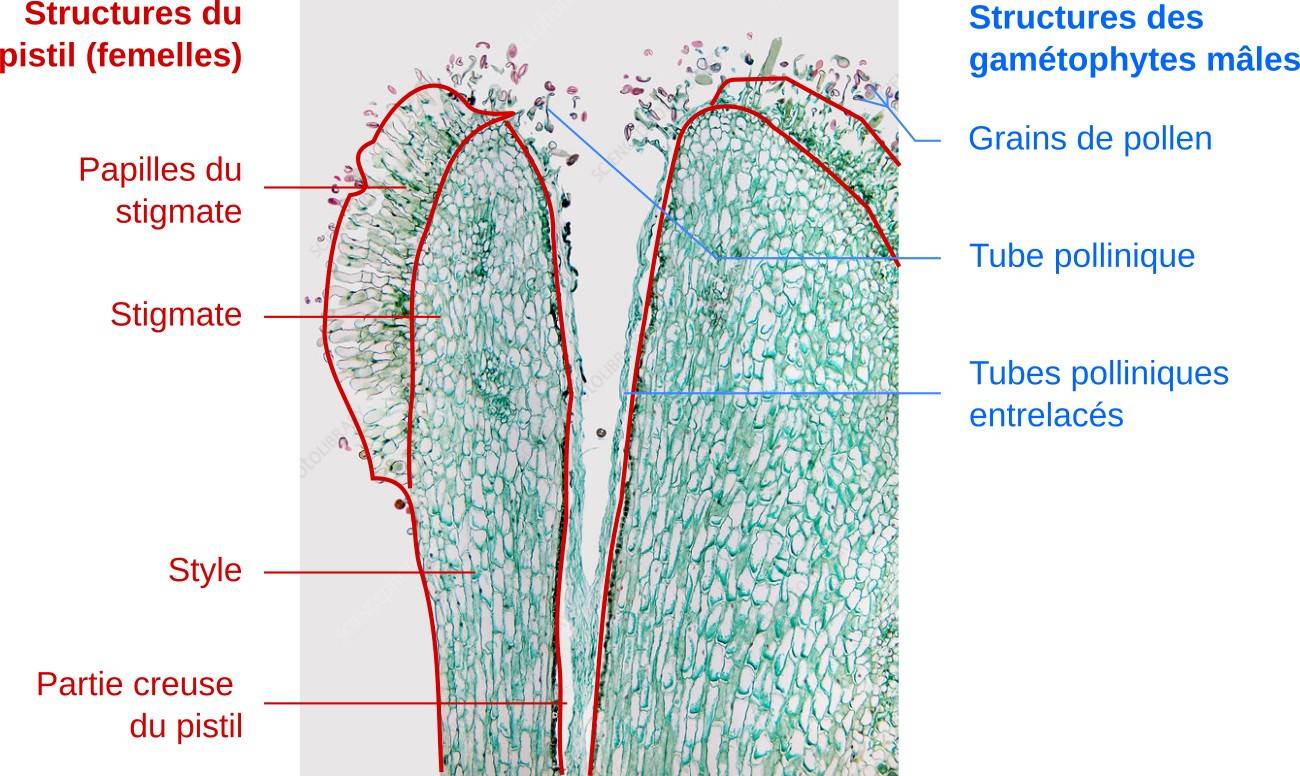
Coupe longitudinale de style de lis, observé en microscopie optique. Les tubes polliniques entrent dans le style creux de la fleur, où ils s’orientent vers les ovules. Les tubes polliniques sont tellement nombreux et entrelacés qu’ils forment de véritables cordons. On peut se demander si cet aspect résulte d’une pollinisation artificiellement abondante, ou s’il se produit en conditions naturelles.

Il est aussi possible de faire germer les tubes polliniques artificiellement pour les étudier. Chez le lis, il suffit de déposer le pollen sur une gélose qui contient du saccharose. Cependant, si cela fonctionne bien pour du lis sauvage, la germination des grains de pollen est plus délicate à obtenir pour des lis ornementaux, peut-être parce qu’ils sont reproduits principalement de façon asexuée et que les grains de pollen ne sont plus fonctionnels.
Ces observations ont permis de constater que le tube pollinique croît rapidement (de l’ordre de 1 cm/h). Cette croissance est assurée par turgescence de la vacuole de la cellule végétative, cette dernière produisant aussi de la paroi sur son extrémité apicale. Le métabolisme intense du grain de pollen est assuré par un transfert de nutriments depuis le style. La croissance du tube pollinique est contrôlée par le style (qui contrôle également le nombre de tubes polliniques qui se forment si plusieurs grains de pollen sont déposés sur le stigmate, ce qui est généralement le cas), puis orientée par chimiotactisme jusqu’à un ovule grâce à des substances chimiques émises par les synergides du sac embryonnaire.
En parallèle, la cellule générative se divise par mitose, ce qui forme deux gamètes mâles. Ils sont transportés par le tube en croissance jusqu’à une synergide où ils sont déversés dans le sac embryonnaire. Dans d’autres espèces d’Angiospermes, les deux gamètes sont formés avant la dispersion du pollen et donc avant la germination du tube pollinique.
Formation des zygotes : une double fécondation siphonogame
À la fin du XIXe siècle, la production de deux gamètes mâles dans le tube pollinique est établie, mais elle pose la question de leur fonction. C’est en en 1898, en observant des coupes de lis pollinisés avec les microscopes les plus performants de l’époque, que le pharmacien et botaniste Léon Guignard y répond1.
Ces coupes montrent qu’après avoir été déversés au niveau du sac embryonnaire, les deux noyaux spermatiques fécondent respectivement l’oosphère et la cellule centrale. Les deux noyaux spermatiques assurent donc une double fécondation ! Il s’agit une synapomorphie des Angiospermes. Le rapprochement des gamètes étant assuré par le tube pollinique, on parle de siphonogamie. Ce mécanisme est également présent chez les Pinopsida (Conifères et Gnétales), et sous une forme un peu différente chez les Cycadales et le Ginkgo. Chez les plantes terrestres dites « à spores libres » (Bryophytes, Lycophytes et Polypodiopsides), les gamètes mâles sont véhiculés par de l’eau, dans le milieu extérieur.
Le zygote diploïde issu de la fécondation de l’oosphère par un gamète mâle est qualifié de zygote principal (futur sporophyte). La fusion triple des noyaux polaires (respectivement triploïde et haploïde dans le cas du lis) et du noyau du gamète mâle produit quant à elle un zygote dit accessoire, pentaploïde.
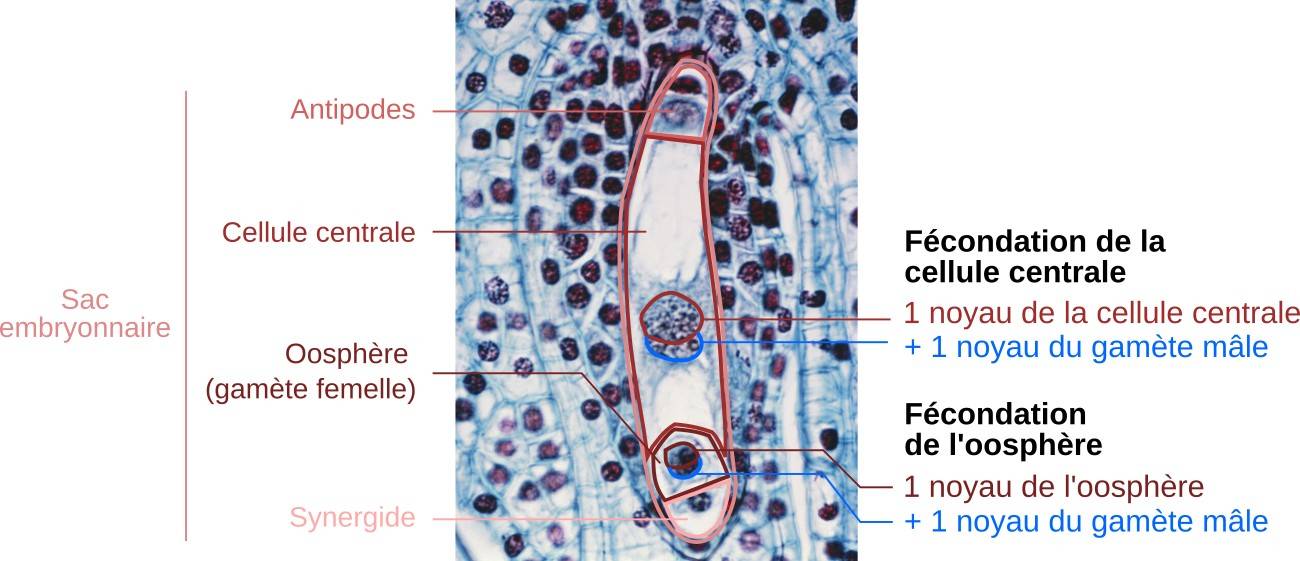
Observation en microscopie optique d’un sac embryonnaire après l’arrivée du tube pollinique. Le micropyle est situé vers le bas de l’image. On distingue bien la double fécondation avec les deux fusions nucléaires : fécondation de l’oosphère par l’un des noyaux spermatiques et fécondation de la cellule centrale tétraploïde par l’autre noyau spermatique.
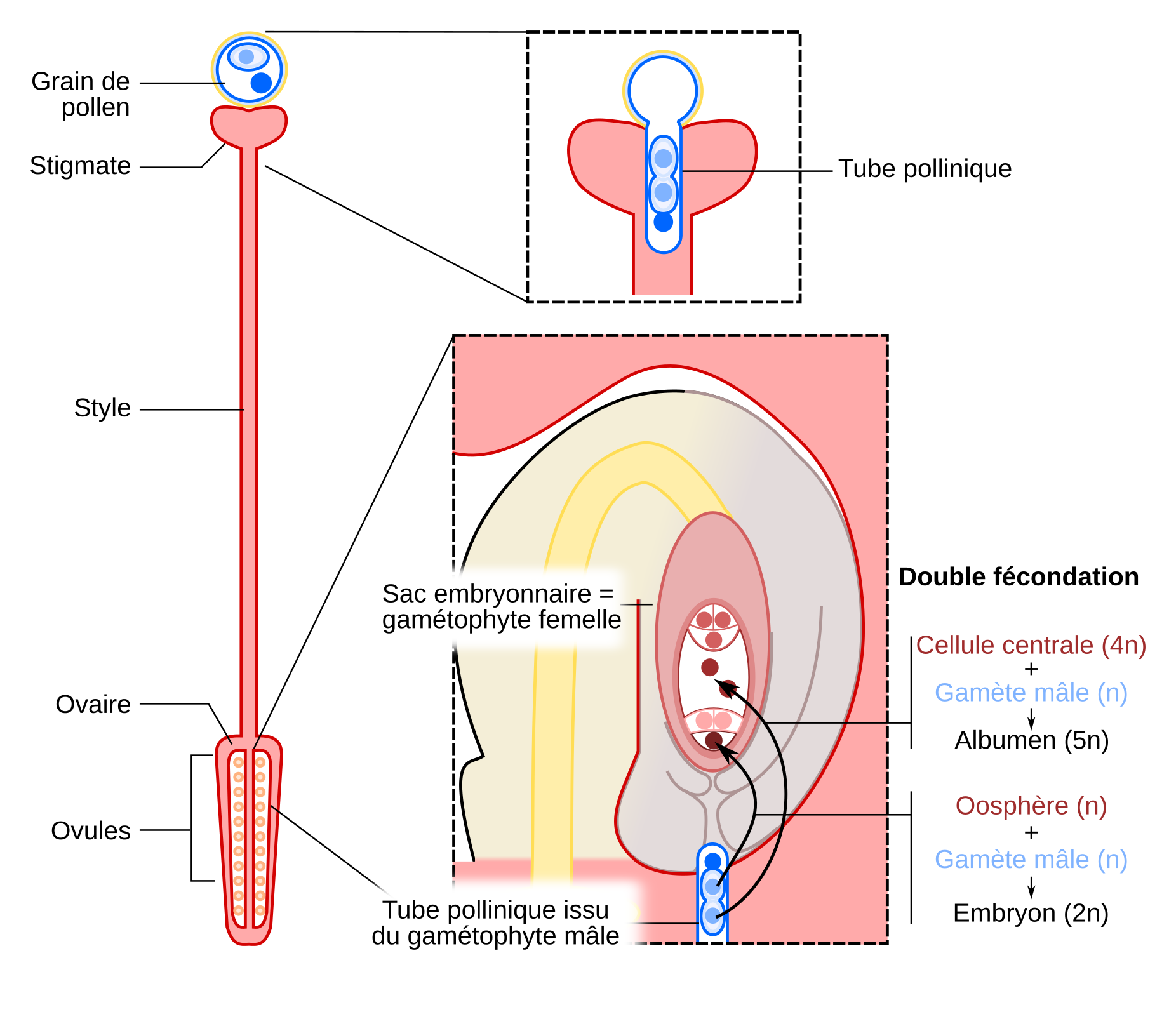
Suite à son dépôt par le vent ou par un pollinisateur sur le stigmate, le grain de pollen forme un tube pollinique dont la croissance permet aux gamètes mâles d’être acheminés depuis le stigmate jusqu’à un ovule en passant par le style. Le tube pollinique arrive finalement au niveau du micropyle d’un ovule et entre en contact avec le sac embryonnaire. L’un des gamètes mâles féconde alors la cellule centrale tandis que l’autre féconde l’oosphère.
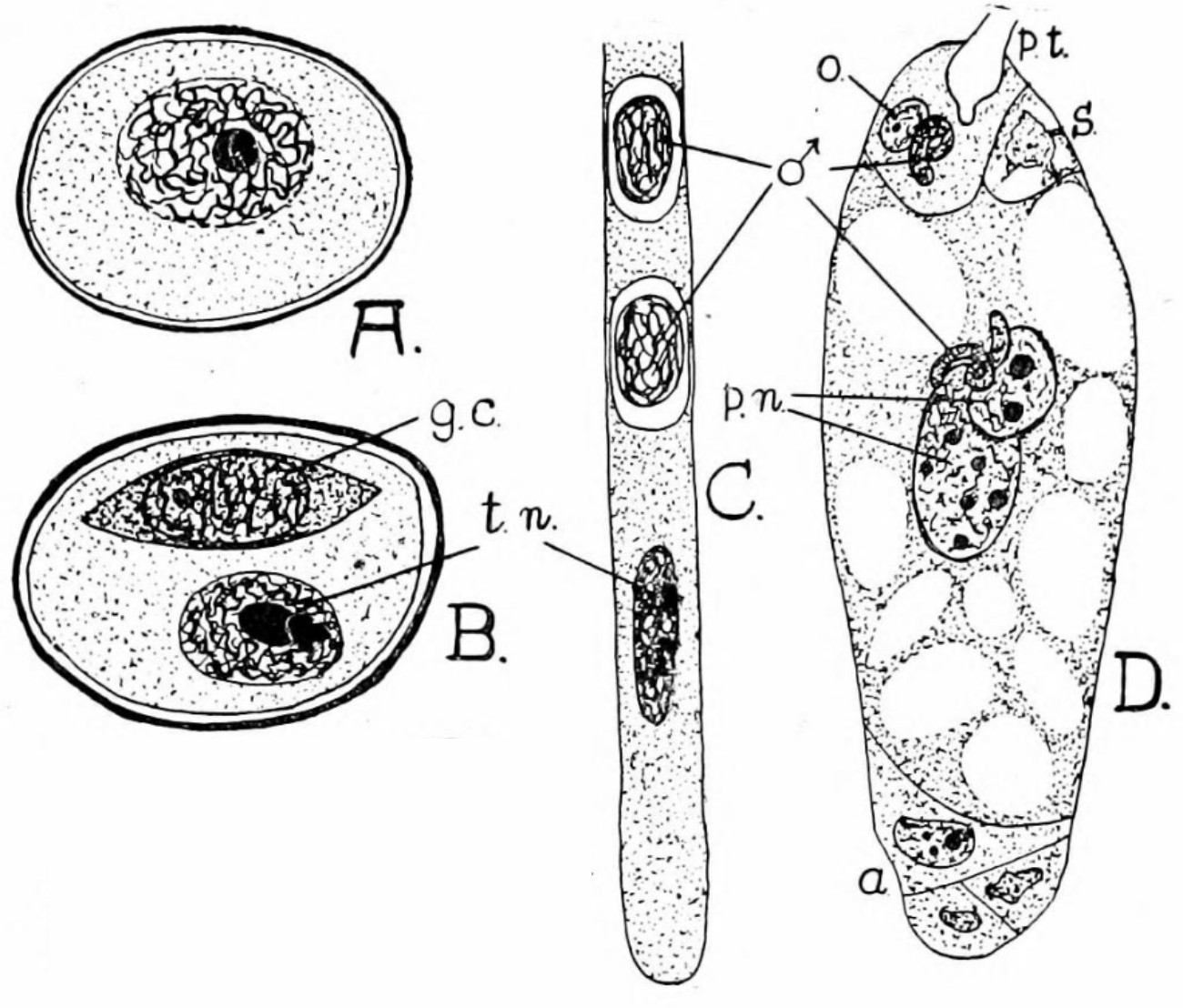
A : Grain de pollen uninucléé. B : Grain de pollen binucléé. C : Portion de tube pollinique avec près de l’apex(vers le bas) le noyau de la cellule végétative et, derrière, les deux gamètes mâles. D : Double fécondation du sac embryonnaire : un noyau mâle est visible au niveau de l’oosphère tandis que l’autre se situe dans la cellule centrale. ♂ : gamètes mâles a : antipodes ; g.c : cellule générative ; t.n : noyau végétatif ; o : oosphère ; p.n ; p.t : tube pollinique ; s : synergide.
Réflexions sur la siphonogamie
On lit (et on entend) souvent qu’avec la siphonogamie, certaines plantes (les Angiospermes, les Pinophytes et les Gnétales) ont complètement « affranchi » leur reproduction vis-à-vis du milieu aquatique. Ce n’est certes pas faux, mais le fait qu’on insiste dessus traduit une vision gradiste de l’évolution qui place certains organismes au-dessus des autres.
En effet, si seule la siphonogamie affranchissait la reproduction vis-à-vis du milieu aquatique, cela sous-entendrait que la zoïdogamie dans une chambre pollinique des Cycadales et du Ginkgo dépend encore du milieu aquatique, ce qui n’est pas le cas. En effet, l’eau impliquée dans cette fécondation est sécrétée par l’organisme lui-même, et non pas présente dans le milieu du fait des conditions abiotiques. Dire que la reproduction zoïdogame des Cycadales et du Ginkgo dépend du milieu aquatique équivaudrait à dire que les Mammifères dépendent encore du milieu aquatique pour réaliser leur fécondation interne zoïdoogame. Cette emphase sur la siphonogamie est donc à proscrire. Il faudrait plutôt considérer que l’affranchissement vis-à-vis du milieu aquatique concerne l’ensemble des espèces qui produisent un tube pollinique, soit toutes les plantes à graines actuelles.
Par ailleurs, les études phylogénétiques et moléculaires récentes montrent que, contrairement à ce qu’on a longtemps pensé, la siphonogamie des espèces actuelles est apparue deux fois indépendamment : chez les Angiospermes et chez les Pinopsida (Conifères et Gnétales) !
Formation des graines et des fruits
Le fruit se développe à partir de l’ovaire et protège et disperse les graines
Après la pollinisation (et les fécondations associées), la fleur fane. Les tépales et les étamines sèchent et tombent, le style et le stigmate se flétrissent. Reste l’ovaire qui grossit et se transforme progressivement en fruit, totalement clos. Il protège et nourrit ainsi la formation des graines (une centaine par fruit environ) situées à l’intérieur. Les graines sont libérées du fruit à maturité. Le fruit est une synapomorphie des Angiospermes (dont l’étymologie signifie justement « graine dans une urne »).
La paroi du fruit s’appelle le péricarpe et est formée de trois couches, de l’extérieur vers l’intérieur :
- l’épicarpe, tissu de revêtement externe du fruit ;
- le mésocarpe, couche intermédiaire ;
- l’endocarpe, couche la plus interne du fruit.
Le fruit mature de lis est un fruit sec, composé des trois carpelles fusionnés qui s’ouvrent par des fentes de déhiscence et libèrent les graines : ce type de fruit s’appelle capsule. La déhiscence se fait sur des fentes passant par le plan de symétrie de chaque carpelle. La texture du mésocarpe et de l’endocarpe (charnue, lignifiée) varie entre les espèces au sein des Angiospermes, et conditionne le mode de dispersion des graines, qui peuvent être dispersées dans le fruit dans le cas des fruits charnus ou des fruits secs qui ne s’ouvrent pas.

On retrouve globalement les structures observées dans l’ovaire de la fleur, dont le volume a considérablement augmenté. Les carpelles forment une enveloppe dans laquelle se développent six rangées de graines. À maturité, des fentes de déhiscence permettront la libération des graines.
La croissance initiale du fruit est contrôlée par la présence d’au moins un embryon fécondé à l’intérieur (via une production d’auxine). Ceci évite des dépenses de ressources à la plante si elle n’a pas été fécondée. Le processus de croissance est permis par quelques mitoses, et principalement par un agrandissement des cellules le composant.
Une fois que le fruit a atteint sa taille définitive, il passe par une phase de maturation. Chez les fruits secs, comme ceux du lis, cela correspond à un dessèchement du péricarpe. Enfin, il y a une étape de sénescence qui correspond à l’ouverture des fentes de déhiscence, ce qui permet la libération des graines.
Après fécondation, les ovules se transforment en graines et l’ovaire se transforme en fruit. La nutrition des graines en croissance est assurée par les faisceaux cribro-vasculaires du péricarpe.
La graine se développe à partir de l’ovule
La graine mature : un emboîtement de générations qui protège et permet le développement du nouveau sporophyte jusqu’à l’autotrophie
Les graines de lis sont libérées à la déhiscence du fruit. Leur structure peut être comprise en les éclairant avec une lumière intense (Figure 32), ou réalisant des coupes (Figure 33).
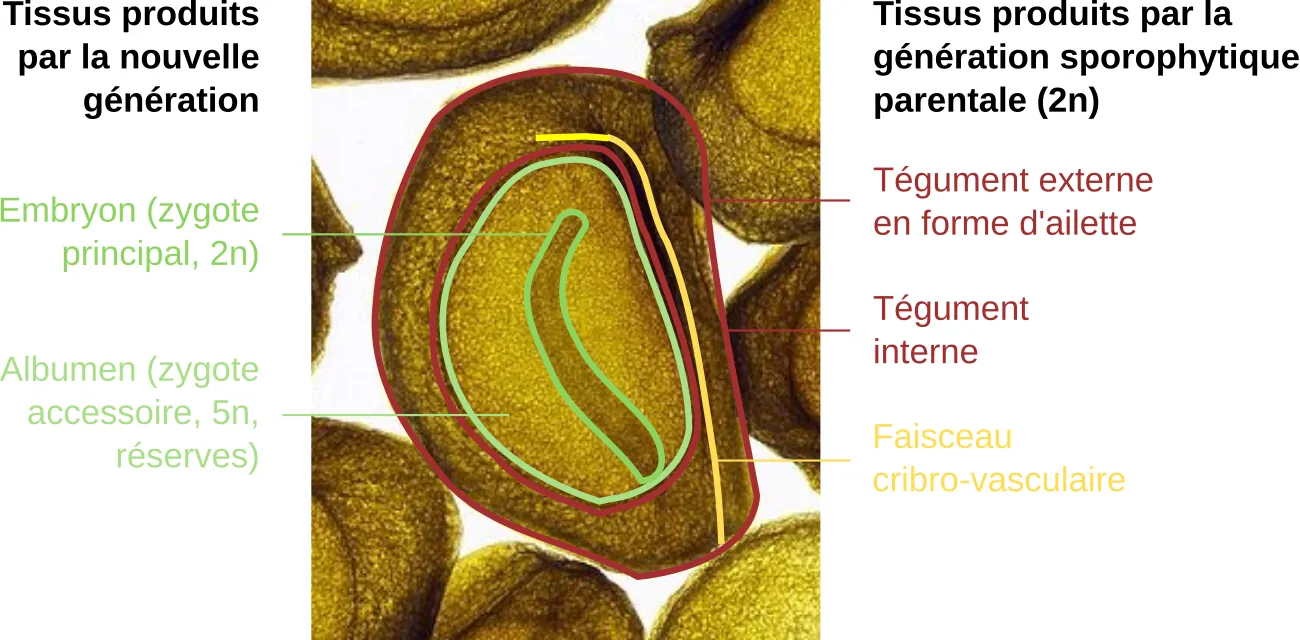
La transparence permet de retrouver globalement les mêmes structures que celles de l’ovule (celles produites par le sporophyte parental), auxquelles s’ajoutent les structures produites par les nouvelles générations (embryon, ou zygote principal diploïde ; et albumen, ou zygote accessoire, un tissu de réserve pentaploïde). Le volume de l’ensemble a considérablement augmenté, et le tégument externe a pris une forme d’ailette qui permettra au vent de disperser la graine. Le zygote principal est différencié avec une jeune racine en bas du cliché annoté, et une tigelle (avec un seul cotylédon et une feuille, indistinguables l’un de l’autre sur cette image).
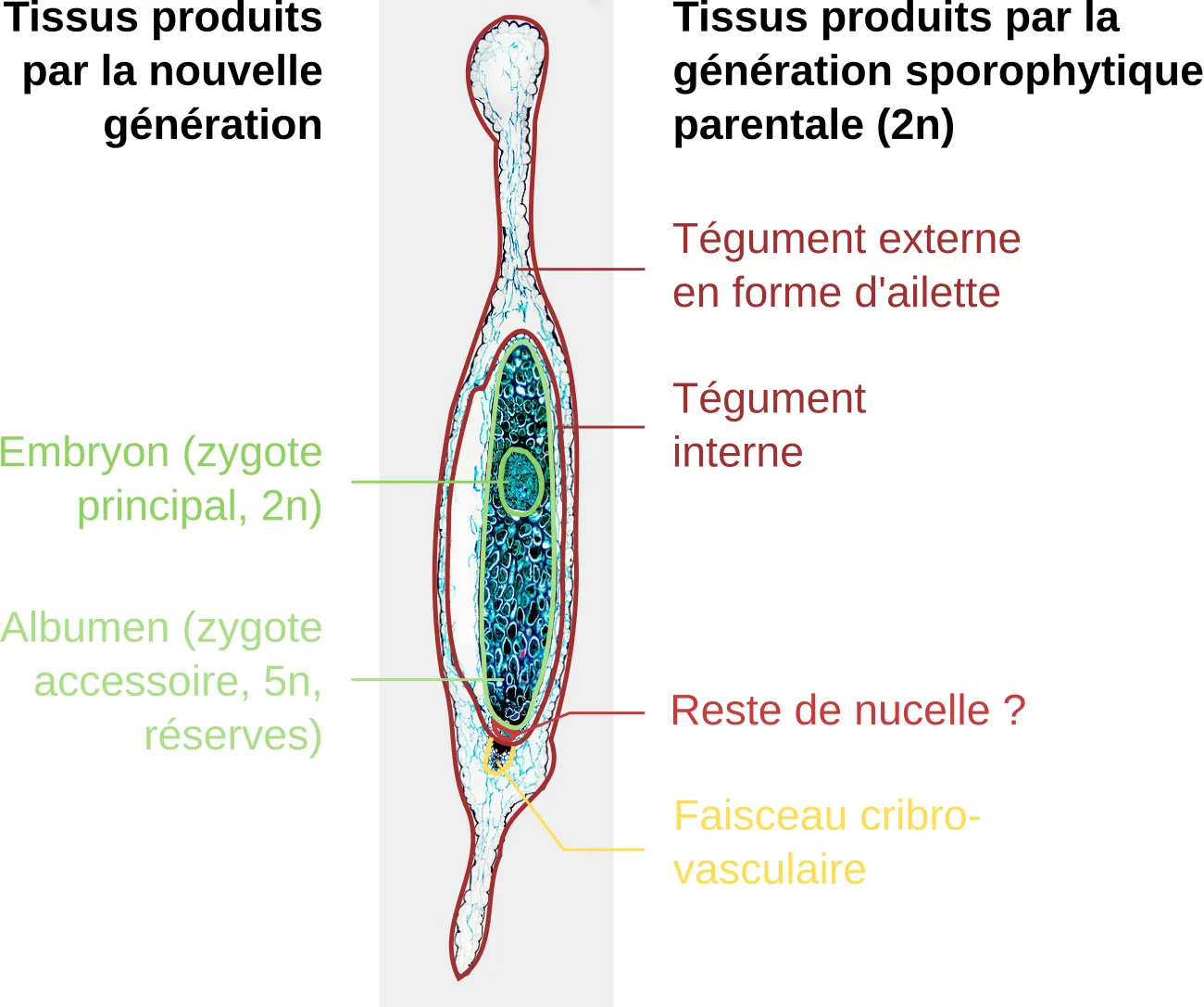
Cette image montre comment une graine est un véritable emboîtement de générations en poupées russes. Une structure inhabituelle est présente entre l’albumen et le tégument interne sur le bas du cliché, il s’agit vraisemblablement du reste du nucelle.
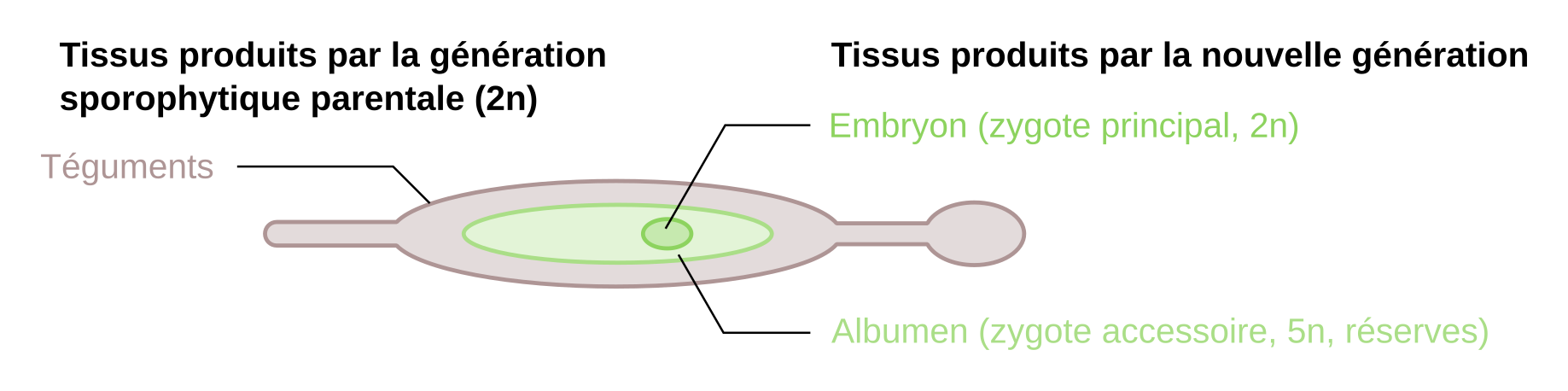
La graine est issue de la transformation de l’ovule suite à la double fécondation. L’embryon provient de la fécondation de l’oosphère et l’albumen de celle de la cellule centrale. Les téguments sont ceux qui entouraient déjà l’ovule.
La graine est constituée de trois générations emboîtées en poupées russes. Au centre se trouve l’embryon. C’est la nouvelle génération sporophytique diploïde, formée par croissance et différenciation du zygote principal. L’embryon est polarisé avec une partie racinaire et une partie caulinaire comprenant un seul cotylédon, typique des Monocotylédones (le reste des Angiospermes, ainsi que la plupart des Acrogymnospermes produisent des embryons à deux cotylédons ou plus), ainsi qu’un méristème apical et une première feuille.
Autour de l’embryon se trouve l’albumen. Il s’agit d’un tissu de réserve formé par multiplication cellulaire du zygote secondaire (pentaploïde chez le lis). Les cellules sont de grande taille et contiennent des réserves qui assurent les besoins du zygote principal au moment de la germination, et jusqu’à l’autotrophie. Ces réserves comprennent différents types chimiques : des glucides qui sont des molécules de réserves métaboliques présentes de façon abondante chez les végétaux, mais aussi des lipides et des protides qu’on ne trouve que dans les graines et les grains de pollen. Les lipides ont en effet un coût énergétique de production important (environ 25 % de l’énergie des glucides utilisés dans leur synthèse est perdue), mais présentent un rapport énergie/masse qui est avantageux pour des formes de dispersion. Les réserves protidiques sont également importantes pour la synthèse de protéines avant que la plante ne puisse puiser de l’azote dans le sol (directement ou via des mycorhizes) et en synthétiser par elle-même.
L’albumen et l’embryon sont protégés par deux téguments, produits par la génération sporophytique parentale diploïde. Cette fonction de protection est assurée par une sclérification des tissus (accumulation de lignine dans les parois cellulaires). Le tégument interne est étroitement accolé à l’albumen. Le tégument externe forme des extensions en ailettes qui favorisent la dispersion de la graine par le vent. Un reste de nucelle semble également visible sur la coupe de graine (Figure 33). La fonction ici n’est pas claire, mais chez d’autres espèces, le nucelle peut former un tissu de réserve continu (appelé périsperme).
Développement et maturation des graines
L’embryon mature est polarisé avec une tigelle et une radicule (respectivement jeune tige et jeune racine) et comprend des milliers de cellules. Le développement embryonnaire implique donc une prolifération cellulaire et une différenciation.
La première division du zygote est polarisée et produit une cellule basale et une cellule apicale. La cellule basale produit le suspenseur, qui enfouit l’embryon dans les tissus nourriciers de l’albumen, et la partie terminale de la racine (coiffe). La cellule apicale produit le reste de l’embryon. Cette partie se met en place par une prolifération initiale à l’origine d’une forme globulaire, puis par la formation d’organes (racine avec méristème apical racinaire ; tigelle avec cotylédon, méristème apical caulinaire et première feuille). L’embryon prend ainsi son aspect définitif jusqu’à la germination.
L’albumen se développe encore plus précocement par prolifération cellulaire. C’est d’ailleurs l’une des fonctions probables de l’albumen : proliférer sans dépendre de contraintes développementales (y compris chez des espèces où il n’est plus présent à la maturité de la graine). L’albumen n’est initialement pas cellularisé (il est laiteux comme l’eau de coco), et les membranes et les parois se mettant en place plus tardivement.
Les téguments croissent autour de l’ensemble et se sclérifient.
Après la prolifération de l’albumen et le développement embryonnaire, une phase de maturation a lieu. C’est à ce moment que les réserves de la graine sont mises en place et qu’elle se déshydrate (autour de 10 % d’eau dans le poids sec chez la plupart des espèces). Elle entre ensuite en vie ralentie.
Les processus liés au développement et à la maturation de la graine nécessitent un certain équilibre : croissance de l’embryon jusqu’à un certain stade mais pas au-delà, déshydratation intense sans mourir. Cela implique un contrôle étroit qui est assuré par les signalisations antagonistes de deux types d’hormones végétales (gibbérellines et acide abscissique).
Libération des graines, dispersion et germination

Le fruit du lis, de type capsule, libère les graines par une déhiscence qui se fait selon des fentes qui parcourent le plan de symétrie du carpelle.
Les graines sont libérées par déhiscence des fruits. Elles sont transportées par la gravité (barochorie) ou par le vent, grâce à l’ailette formée par le tégument (anémochorie). Une graine ne germe pas tout de suite après avoir été libérée de son fruit, même si elle se trouve dans des conditions favorables. En effet, elle présente un état de vie ralentie appelé dormance. Il s’agit d’une adaptation à la saisonnalité, qui empêche une germination avant que la mauvaise saison ne soit passée ; celle-ci ne peut se produire qu’au printemps suivant. Les cultivateurs savent accélérer le processus en plaçant les graines de lis dans de la vermiculite humide soumise quelques jours au froid, puis au chaud, puis au froid. Le froid et la chaleur semblent donc être deux des facteurs qui permettent la levée de dormance chez les lis.
Quand la dormance est levée et que la graine se trouve dans des conditions favorables (présence d’eau, température favorable permettant le métabolisme), la graine germe. Ce processus commence par une imbibition, c’est-à-dire par une réhydratation des tissus grâce à l’entrée d’eau dans la graine qui gonfle passivement. L’embryon reprend progressivement un métabolisme normal. Des protéines de dégradation des réserves sont synthétisées à partir des acides aminés issus des réserves protidiques. À partir de là, la plantule croît activement, produit d’abord une racine puis un appareil caulinaire. Chez le lis martagon, la germination est hypogée, c’est-à-dire que le développement de la plantule se fait initialement sous le sol, et la première feuille n’apparaît qu’au printemps. Chez d’autres espèces, la germination est épigée avec un développement rapide de feuilles en surface. La plantule prend directement une forme de bulbe, que des racines contractiles participent à enfouir dès le début de sa vie. Quand les structures de prélèvement sont suffisamment développées, la plante est autotrophe et indépendante et continue à croître.
Tous les ans, la partie aérienne meurt avant l’automne, et la plante résiste à la mauvaise saison sous forme de bulbe situé sous terre. Les deux ou trois premières années, les tiges formées ne produisent que des feuilles. C’est au bout de trois à quatre ans, et chaque printemps, que la plante devient mâture et produit, en plus des feuilles, une grappe de fleurs à l’extrémité de la tige. Le cycle de vie est bouclé, même si l’individu peut vivre plusieurs dizaines d’années.
Bilan : cycle de vie du lis
À partir des différents éléments présentés ici, nous pouvons récapituler le cycle de vie du lis (Figure 36).
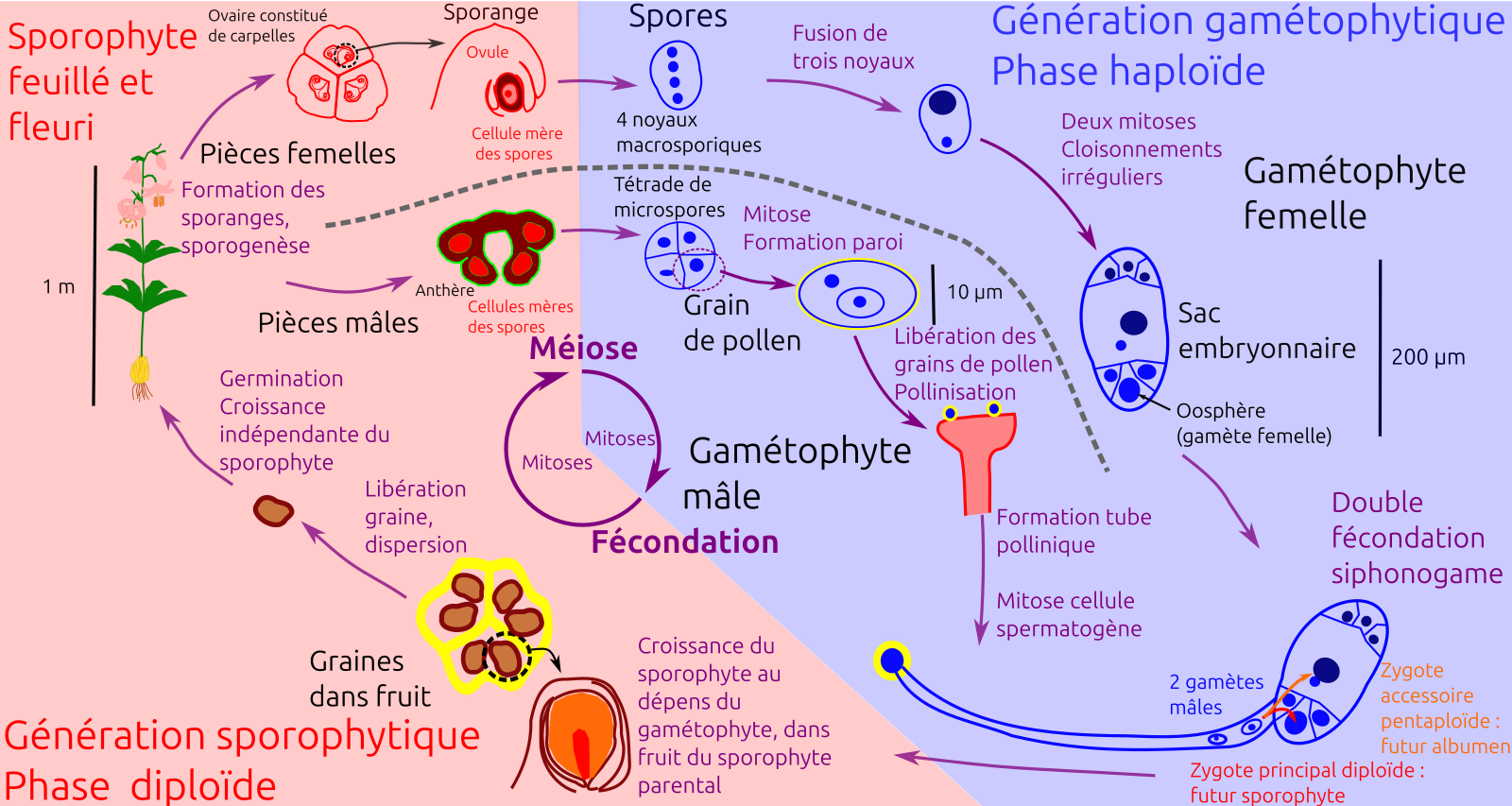
Violet : principales étapes du cycle de reproduction avec au centre les étapes nucléaires fondamentales ; noir : principales structures associées.
Nous pouvons souligner quelques caractéristiques de ce cycle de vie :
-
La présence de deux générations morphologiques distinctes et successives. Le sporophyte, diploïde, correspond au pied feuillé et fleuri. Il produit les spores. Les gamétophytes mâle et femelle (respectivement grains de pollen, libérés, et sac embryonnaire, se développant au sein des tissus du sporophyte), haploïdes, produisent les gamètes. Le cycle de reproduction du lis se caractérise donc par l’alternance entre une génération diploïde et une génération haploïde : il est qualifié d’haplo-diplophasique.
-
Cette alternance de deux générations est permise par les étapes fondamentales de la sexualité eucaryote que sont la méiose et la fécondation. La méiose produit les spores haploïdes, tandis que la fécondation réunit deux cellules haploïdes en une cellule diploïde. Entre chacune de ces deux étapes, des mitoses permettent le développement de deux types d’individus : sporophyte (individu feuillé) et gamétophytes (sac embryonnaire et grain de pollen).
-
La présence d’une génération haplophasique présente l’avantage de pouvoir purger des mutations délétères d’une population. En effet, chez les gamétophytes, haploïdes, chaque allèle n’est présent qu’en un seul exemplaire. Un allèle délétère qui empêcherait le développement des gamétophytes ne pourrait donc être transmis à la génération suivante.
-
La fécondation est strictement indépendante de l’eau du milieu extérieur. Elle est siphonogame c’est-à-dire que les gamètes sont directement apportés jusqu’au sac embryonnaire par l’intermédiaire du tube pollinique. Autrement dit, les gamètes ne sont pas doués d’une mobilité propre grâce à des flagelles, contrairement à ce qui est observé dans les fécondations zoïdogames. Enfin, la fécondation est double : deux gamètes mâles sont transportés jusqu’au gamétophyte femelle par un tube pollinique.
-
La diplophase est dominante. Chez les plantes à graines, ceci est tellement exacerbé qu’on a l’impression à première vue d’un cycle à une seule génération (et ce qui est valable pour des études écologiques).
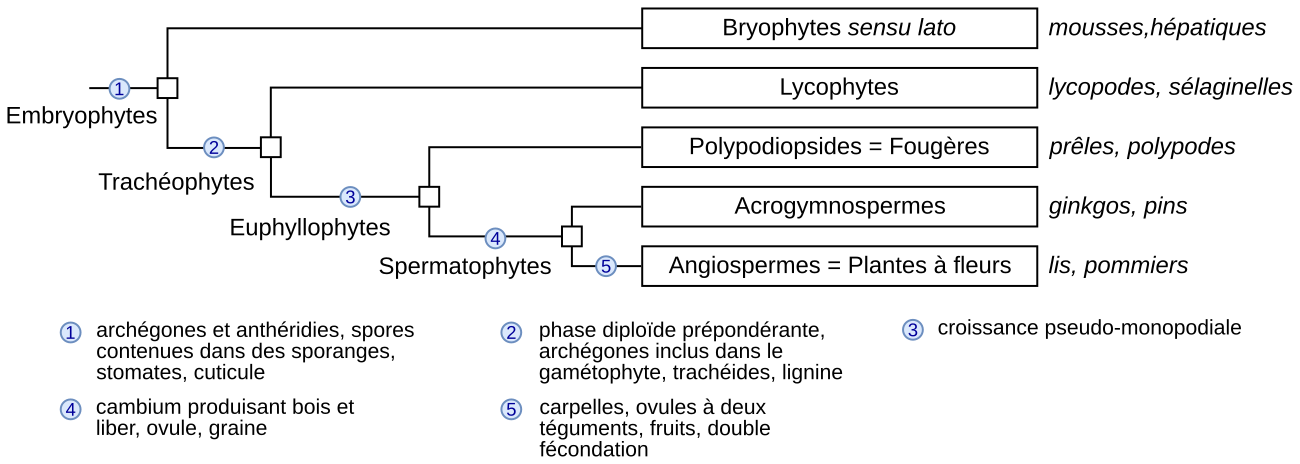
Ces caractéristiques générales du cycle de vie (alternance des générations gamétophytiques et sporophytiques séparées par la fécondation et la méiose) sont les mêmes que chez le polypode, et que chez toutes les Embryophytes en général. Ce mode de développement étant hérité d’un ancêtre commun, les générations sont homologues dans les différents groupes d’Embryophytes.

Certaines caractéristiques du cycle de vie des Embryophytes ont une valeur cladistique en définissant des groupes évolutivement cohérents, c’est-à-dire monophylétiques :
-
Le cycle à deux générations avec fécondation zoïdogame correspond au cycle ancestral des Embryophytes. Il est toujours présent chez les Bryophytes et les Polypodiopsides. Les végétaux de ces lignées se caractérisent également par la présence d’organes spécialisés dans la production des spores (sporanges) et des gamètes (anthéridies et archégones, disparus chez les Angiospermes). Chez les Angiospermes, les sporanges sont réduits à des tissus contenus dans le sac pollinique et dans l’ovule.
-
La présence de plusieurs sporanges sur un même sporophyte et d’une diplophase dominante est partagée par toutes les Trachéophytes actuelles (chez lesquelles on trouve également des éléments vasculaires lignifiés).
-
La présence de graines, qui dérivent d’ovules, est partagée par toutes les Spermatophytes (plantes à graines). Un ovule est une structure de reproduction femelle dans laquelle différentes étapes de la reproduction sont internalisées, ce qui permet leur protection et leur nutrition, et finalement leur raccourcissement. Plus en détail, un ovule comprend des structures protectrices (tégument(s) et nucelle, ce dernier correspondant au sporange femelle) qui protègent la sporogenèse (formation des spores par méiose), le développement du gamétophyte femelle, la fécondation et enfin le développement embryonnaire et la maturation de la graine. Un ovule se transforme en graine, qui est un emboîtement de générations en poupées russes protégé par le (ou les) tégument(s) d’origine parentale. À l’intérieur se trouve l’embryon (nouvelle génération sporophytique), associé à des réserves (d’origine très variable selon les espèces). La graine mature est libérée du sporophyte parental et constitue le stade de dispersion des Spermatophytes.
-
Le carpelle, partie de la fleur qui devient fruit, est sans doute la synapomorphie la plus remarquée des Angiospermes. Le carpelle est souvent interprété comme une sporophylle modifiée et close contenant les ovules puis les graines. Il a rôle très important, en protégeant une grande partie des étapes de reproduction puis en participant à la dispersion des graines.
Plusieurs synapomorphies des Angiospermes ressortent aussi d’études plus approfondies : ovules à deux téguments, gamétophytes très réduits (généralement 7 cellules et 8 noyaux pour le gamétophyte femelle, 3 cellules à maturité pour le gamétophyte mâle), siphonogamie, et double fécondation avec formation d’un zygote principal et d’un zygote accessoire (albumen). La fleur est en elle-même la synapomorphie la plus voyante des Angiospermes, autrement nommées plantes à fleurs.
Conclusion
La reproduction sexuée du lis est typique de la reproduction sexuée des Angiospermes, car on y retrouve toutes les structures et les étapes clés. Elle a lieu sur les pieds feuillés et fleuris, qui sont les sporophytes. La plupart des étapes se déroulent au niveau de la fleur puis du fruit, où elles sont internalisées, ce qui assure la protection et la nutrition des structures impliquées. Les parties fertiles de la fleur sont les étamines et le pistil, et plus spécifiquement les sacs polliniques et les ovules, dans lesquels la sporogenèse a lieu par méiose. Des gamétophytes mâle et femelle (respectivement grain de pollen et sac embryonnaire) se développent ensuite par mitoses. La pollinisation permet le rapprochement des gamétophytes par transport des grains de pollen de l’anthère au pistil. La croissance d’un tube pollinique permet ensuite d’acheminer deux gamètes mâles jusqu’à un sac embryonnaire, où a lieu une double fécondation. La fleur fane et laisse place à un fruit en développement, dans lequel se forment des graines. Chaque graine contient un embryon et un tissu de réserve (albumen). À la maturité des graines et des fruits, les graines sont libérées et dispersées, et forment de nouveaux plants feuillés après germination.
Cependant, l’exemple de la reproduction du lis, présenté ici pour illustrer les mécanismes de la reproduction des Angiospermes, ne saurait rendre compte de la diversité phénoménale des systèmes reproducteurs des Angiospermes : architecture des fleurs (nombre de verticilles et de pièces florales, morphologie des pièces florales…), diversité des fruits (secs ou charnus, déhiscents ou non…), modalités de développement du gamétophyte (ploïdie des noyaux), origine des réserves des graines (albumen, cotylédons), etc. L’étude des structures reproductrices des Angiospermes est intéressante, car elle apporte deux informations : identité et parenté de la plante porteuse ; fonctionnement reproducteur.
La forme des fleurs donne une première indication de la position phylogénétique de la plante qui la porte, car elle est assez bien conservée évolutivement. Linné, éminent naturaliste suédois du XVIIIe siècle, a produit un système de classification original entièrement fondé sur les organes reproducteurs des fleurs. De nombreux systèmes de classification ont été produits avant et après lui, prenant en compte non seulement les fleurs mais également d’autres parties des plantes comme le nombre de cotylédons. Jusqu’à Darwin, ces classifications étaient inscrites dans un cadre de pensée fixiste, ignorant l’évolution biologique. C’est sur l’architecture florale que les premières familles d’Angiospermes ont pu être établies ; leur nombre s’élève aujourd’hui à presque 450. Un certain nombre de ces familles, notamment les plus connues (Asteraceae, Lamiaceae) ont été validées par des classifications moléculaires récentes. La famille des Liliaceae, à laquelle appartient le lis, a en revanche connu une réorganisation drastique à la fin du XXe siècle, une grande partie de ses représentants ayant été réaffectés à d’autres familles (Asparagaceae notamment).
Cette phénoménale diversité des structures de reproduction sexuée des Angiospermes informe aussi de leur diversité de fonctionnement. L’architecture et la couleur des fleurs contrôlent en effet le mode de pollinisation (vent, nombreux et divers insectes, chiroptères, oiseaux, etc.), et de même pour les différents types de fruits qui contrôlent le mode de dispersion (autonome, par le vent, les animaux, etc.).
Il existe actuellement environ 300 000 espèces d’Angiospermes, contre à peine 1000 chez les Acrogymnospermes qui n’ont ni fleur ni fruit. La diversité des Angiospermes est donc probablement à relier à la présence des fleurs et des fruits qui, parce qu’ils dépendent souvent des animaux pour la pollinisation ou la dispersion des semences (fruits ou graines), entraînent une coévolution, voire une coadaptation entre Angiospermes et Animaux. Ainsi une spéciation chez un des partenaires de ces relations mutualistes peut entraîner une spéciation chez l’autre partenaire, ce qui pourrait expliquer la radiation importante des Angiospermes et de leurs principaux pollinisateurs, les Insectes. Les Animaux ont un rôle particulièrement important, car ils permettent la reproduction des Angiospermes même dans de petites populations, grâce à un transport orienté du pollen (à l’inverse d’un transport aléatoire par le vent, principal vecteur de pollen des Acrogymnospermes1). L’existence de petites populations d’Angiospermes est favorable à une évolution séparée de celles-ci et donc à des phénomènes de spéciation. Les Animaux peuvent également favoriser ces phénomènes en dispersant des graines dans des environnements nouveaux. Des études montrent que d’autres caractères ont eu, en complément de la coévolution avec les Animaux, un rôle important dans la diversification des Angiospermes. Il s’agit en particulier de la possibilité d’avoir un cycle de vie annuel, de la capacité à se reproduire de façon sexuée et asexuée, de la capacité à évoluer des nouvelles formes florales (gènes homéotiques), ou de la capacité à former des hybrides polyploïdes.
Remerciements
L’auteur remercie Margarethe Maillart et Damien Mollex pour leur aide technique ayant permis la réalisation de l’article.



