Les rétinites pigmentaires forment un groupe de maladies héréditaires se caractérisant par une perte progressive de la vision, suite à la dégénérescence des photorécepteurs. D’origine génétique, elles constituent un modèle de choix pour la recherche de traitements, notamment par thérapie génique. Nous développons ici une étude réalisée par une équipe chinoise et allant de la modélisation d’une rétinite pigmentaire chez la souris au test d’un nouveau traitement par thérapie génique. Ce dernier repose sur une stratégie de réécriture de base dérivée des outils CRISPR/Cas9 et utilisant comme vecteurs des virus adénoassociés recombinants.
Les dégénérescences rétiniennes constituent une importante source de perte de la vision à l’échelle mondiale. Parmi ces pathologies, les rétinites pigmentaires sont des dégénérescences d’origine génétique qui constituent un modèle privilégié pour la recherche de traitements, notamment par thérapie génique. En effet, les gènes responsables des rétinites pigmentaires chez l’être humain peuvent être mutés chez un animal de laboratoire (souris ou rat le plus souvent ; chat, chien ou macaque plus exceptionnellement). Ces modèles animaux permettent une meilleure compréhension des mécanismes à l’origine des pathologies et de tester de nouvelles pistes de traitements. Ce processus est malheureusement plus compliqué dans les pathologies multifactorielles ou provoquées par le vieillissement (comme la dégénérescence maculaire liée à l’âge ou DMLA), plus difficiles à recréer artificiellement chez les animaux.
Modélisation d’une rétinite pigmentaire chez la souris
Yidong Wu et son équipe 1 ont travaillé avec des souris mutantes rd10 possédant une mutation dans le gène de la phosphodiestérase PDE6B, une des protéines de la cascade de transduction du signal lumineux dans les photorécepteurs. Dans cette lignée de souris, la mutation provoquée empêche l’expression de la phosphodiestérase.
Les chercheurs ont commencé par observer le phénotype rétinien des souris, pour vérifier qu’il reproduit bien le phénotype des rétinites pigmentaires. Les rétines des souris mutantes rd10 présentent une couche nucléaire externe (celle qui contient les noyaux des photorécepteurs) beaucoup plus fine que chez les souris sauvages. En effet, cette couche nucléaire externe contient alors un nombre très réduit de noyaux. Par ailleurs, les rétines des souris mutantes ne présentent plus de rhodopsine ni de segments externes des photorécepteurs (le segment externe est le compartiment spécialisé dans la photosensibilité chez les photorécepteurs, cônes ou bâtonnets). Ces trois indicateurs sont liés à la dégénérescence des bâtonnets :
- les noyaux des bâtonnets forment la quasi-totalité des noyaux de la couche nucléaire externe, leur dégénérescence diminue drastiquement le nombre de noyaux présents ;
- lors de leur dégénérescence, le segment externe est l’une des premières structures affectée et perdue par les bâtonnets ;
- une fois les bâtonnets dégénérés, la rhodopsine, qui leur est spécifique et nécessaire à leur photosensibilité, n’est plus exprimée dans la rétine.
Le phénotype observé mime donc bien une rétinite pigmentaire chez la souris.
Exemple de traitement par virus adénoassocié recombinant et réécriture de base chez les souris rd10
L’équipe de recherche a utilisé une stratégie reposant sur un virus adénoassocié recombinant et un outil d’édition génétique dérivé de CRISPR/Cas9, la réécriture de base (ou édition de base ; base editing en anglais).
Un virus adénoassocié est un petit virus non pathogène incapable de se répliquer de façon autonome (d’où le nom d’adénoassocié, car ces virus nécessitent la co-infection avec d’autres vecteurs viraux comme les adénovirus dans leur réplication). Le terme de virus adénoassocié recombinant est utilisé lorsqu’une capside virale de virus adénoassocié est vidée du matériel génétique normal du virus puis recombinée avec un contenu génétique artificiel. La capside sert alors de vecteur. Dans le cadre de l’étude de Wu et de ses collaborateurs, le ciblage spécifique des cellules photoréceptrices est obtenu grâce à un certain sérotype de virus adénoassocié, dont les protéines de capside permettent une interaction spécifique avec les cellules photoréceptrices.
Le contenu génétique empaqueté dans ces vecteurs correspond à une séquence codant des protéines du système CRISPR/Cas9 et à un ARN guide permettant à ces protéines de corriger uniquement et spécifiquement le gène muté de la PDE6B.
Après une phase de test pour choisir le contenu génétique le plus adapté, l’équipe de chercheurs a testé son assemblage le plus prometteur en injectant sous la rétine des souris mutantes rd10 l’une des trois solutions suivantes :
- une solution saline ne contenant pas de capside virale du tout ;
- une capside contenant le système d’édition du génome, mais avec un ARN guide ne ciblant aucun gène et ne permettant donc pas la correction du gène PDE6B muté ;
- une capside contenant le système d’édition du génome, avec un ARN guide ciblant la mutation du gène PDE6B portée par les souris rd10.
Pour disposer de témoins à l’intérieur même des souris testés, l’injection a systématiquement été réalisée du côté temporal de la rétine (à droite pour l’œil droit, à gauche pour l’œil gauche). Il est ainsi possible de comparer la rétine côté temporal avec la rétine côté nasal, car les virus restent localisés côté temporal, ils ne diffusent pas jusqu’au côté nasal.
Expression de PDE6B dans les différentes conditions
Pour vérifier si le système de réécriture de base a bien permis, après correction du gène, de rétablir l’expression de la phosphodiestérase PDE6B, les auteurs ont réalisé des Western blots (transferts de protéines) et des immunohistochimies (Figure 1).
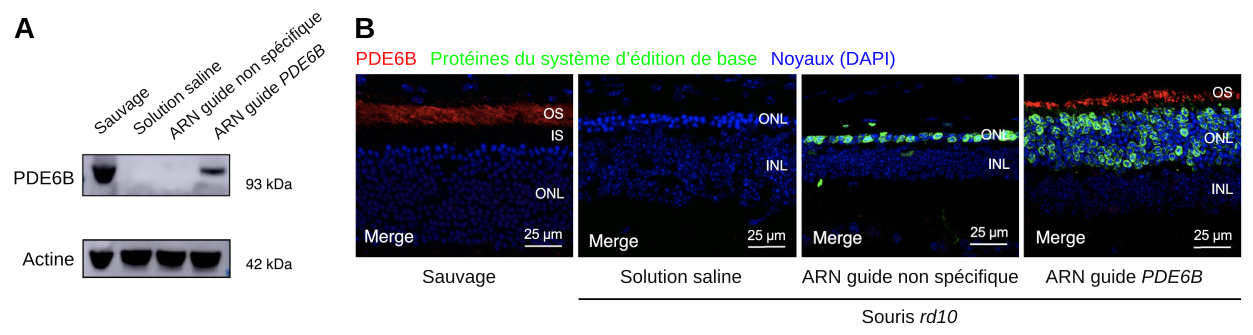
A. Transfert de protéines (Western blot) dirigé contre la protéine PDE6B. L’actine sert de témoin de charge. Les souris mutantes rd10 ont été injectées avec une solution saline, un système de réécriture de base contenant un ARN guide non spécifique ou un système de réécriture de base contenant un ARN guide spécifique de la mutation du gène PDE6B portée par les souris rd10.
B. Des coupes de rétine sont colorées avec un intercalant de l’ADN, le DAPI, révélant ainsi les noyaux (bleu). PDE6B et les protéines du système de réécriture de base sont révélées par des anticorps spécifiques couplés respectivement à une molécule fluorescente rouge ou verte. OS : couche des segments externes des photorécepteurs ; ONL : outer nuclear layer, couche nucléaire externe, contenant les noyaux des photorécepteurs ; INL : inner nuclear layer, couche nucléaire interne, contenant les noyaux des cellules bipolaires notamment.
Le Western blot confirme l’absence de production de PDE6B chez les souris rd10 injectées avec une solution saline ou un système de réécriture de base non fonctionnel (Figure 1A). Il montre également que le système de réécriture de base fonctionnel permet de rétablir la production de PDE6B chez les souris mutantes.
L’immunohistochimie détecte les protéines du système de réécriture de base (en vert), qui sont bien sélectivement exprimées dans les cellules de la couche nucléaire externe, c’est-à-dire dans les photorécepteurs (Figure 1B). De plus, la phosphodiestérase PDE6B est exprimée chez les souris rd10 uniquement dans la condition utilisant l’ARN guide spécifique de la mutation du gène PDE6B. Ce système permet de rétablir l’expression de PDE6B dans la zone où cette protéine est exprimée physiologiquement, à savoir au niveau du segment externe des photorécepteurs.
Quantification de la dégénérescence des photorécepteurs
L’observation de l’épaisseur de la couche nucléaire externe donne une bonne estimation de la dégénérescence des bâtonnets : si elle est très épaisse, peu de bâtonnets ont dégénéré, si elle est fine, la dégénérescence a été très marquée.
Les résultats présentés en Figure 2 montrent que le système thérapeutique proposé par l’équipe chinoise permet bien une dégénérescence cellulaire beaucoup moins marquée que dans les souris rd10 où la mutation du gène PDE6B n’est pas corrigée. Le sauvetage n’est cependant pas complet, puisque l’épaisseur de la couche nucléaire externe reste inférieure à celle observée chez les souris sauvages.
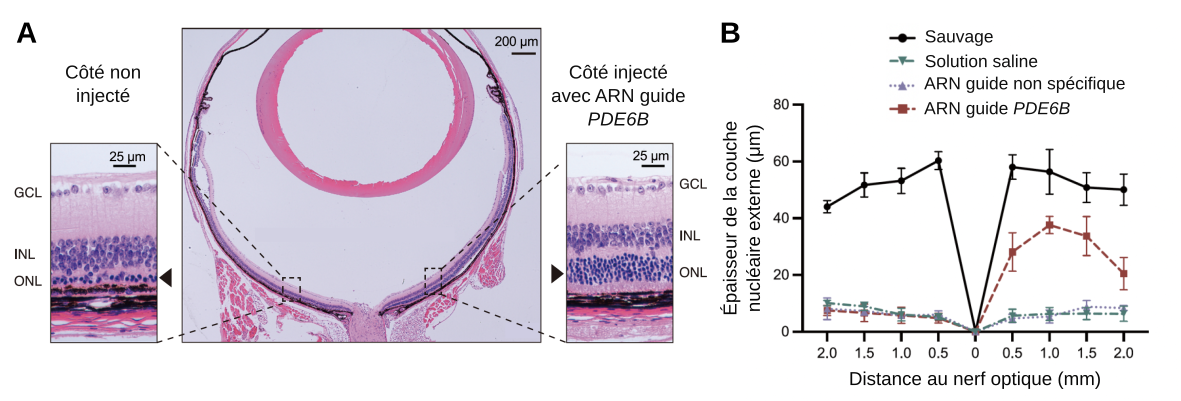
A. Histologie en coloration hématoxyline éosine des parties nasale et temporale chez une souris mutante rd10 injectée avec le système de réécriture de base fonctionnel. La couche nucléaire externe (ONL) est repérée par une tête de flèche. INL : inner nuclear layer, couche nucléaire interne, contenant les noyaux des cellules bipolaires notamment. GCL : ganglion cell layer, couche des cellules ganglionnaires.
B. Épaisseur de la couche nucléaire externe chez les souris sauvages et chez les souris rd10. Les souris mutantes rd10 ont été injectées, côté temporal (moitié droite du graphique), par une solution saline, un système de réécriture de base contenant un ARN guide non spécifique ou un système de réécriture de base contenant un ARN guide spécifique de la mutation du gène PDE6B portée par les souris rd10. La moitié gauche du graphique correspond au côté nasal.
Test comportemental de la vision des souris
Pour tester si les photorécepteurs génétiquement modifiés permettaient effectivement de restaurer la vision chez les souris rd10, un test comportemental d’orientation dans une piscine a été réalisé (Figure 3). Les souris sont déposées dans une piscine où une plateforme visible leur permet de se mettre au sec. Les souris sauvages détectent rapidement la plateforme, et progressent au cours du temps : plus elles réalisent le test, plus elles se repèrent et se placent rapidement sur la plateforme. Une souris dont la vision est déficiente mettra plus de temps à trouver la plateforme, qu’elle atteindra le plus souvent par hasard, en explorant la piscine, aucun autre indice sensoriel n’étant à sa disposition (pas d’odeur notamment, ce qui est important chez les souris qui utilisent bien plus leur odorat que les êtres humains).
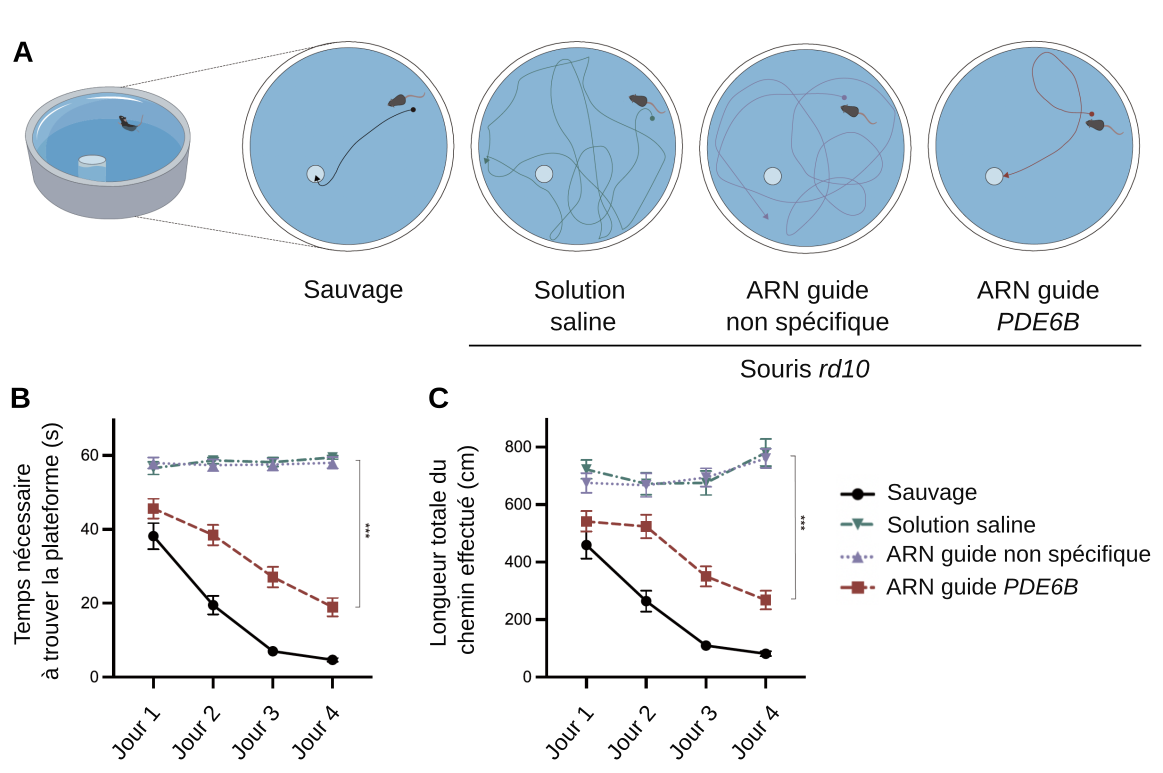
A. Les souris sont déposées dans une bassine d’eau contenant au centre une plateforme.
B et C. Le temps passé dans l’eau et la distance parcourue avant de trouver la plateforme sont mesurées. Les souris mutantes rd10 ont été injectées avec une solution saline, un système de réécriture de base contenant un ARN guide non spécifique ou un système de réécriture de base contenant un ARN guide spécifique de la mutation du gène PDE6B portée par les souris rd10.
Les résultats montrent que les souris mutantes rd10 injectées avec un système de réécriture de base fonctionnel présentent des trajectoires bien plus efficaces pour retrouver la plateforme que les souris rd10 injectées avec une solution saline ou avec un système de réécriture de base de base non fonctionnel. Cela permet de démontrer une restauration au moins partielle des capacités visuelles chez les souris rd10 traitées. Les souris traitées n’ont donc pas seulement vu la dégénérescence des photorécepteurs limitée, mais ces photorécepteurs préservés permettent une vision fonctionnelle chez ces animaux.
Conclusion
Les virus adénoassociés sont utilisés avec succès dans des thérapies géniques actuellement en essais cliniques voire, pour certaines, déjà arrivées jusqu’aux patients. L’utilisation d’outils génétiques dérivés de CRISPR/Cas9 est, elle, encore en développement. Dans cette étude, l’utilisation d’un modèle animal de rétinite pigmentaire a permis aux chercheurs de réaliser une preuve de concept sur l’efficacité d’un traitement par thérapie génique par édition du génome rassemblant ces deux outils. Les auteurs espèrent dans le futur pouvoir transférer cette stratégie chez l’humain et proposer de nouvelles pistes thérapeutiques aux patients.