
La rétine a pendant longtemps été décrite comme un simple dispositif de conversion de signaux lumineux en signaux électriques, le cerveau étant en charge des traitements considérés comme complexes, en particulier la détection de bordures ou de mouvements. Ce n’est qu’au milieu du XXe siècle qu’a émergée l’idée que des traitements complexes de l’information pouvaient avoir lieu de façon précoce dès la rétine. Ces traitements permettent de transmettre l’information visuelle de façon efficace.
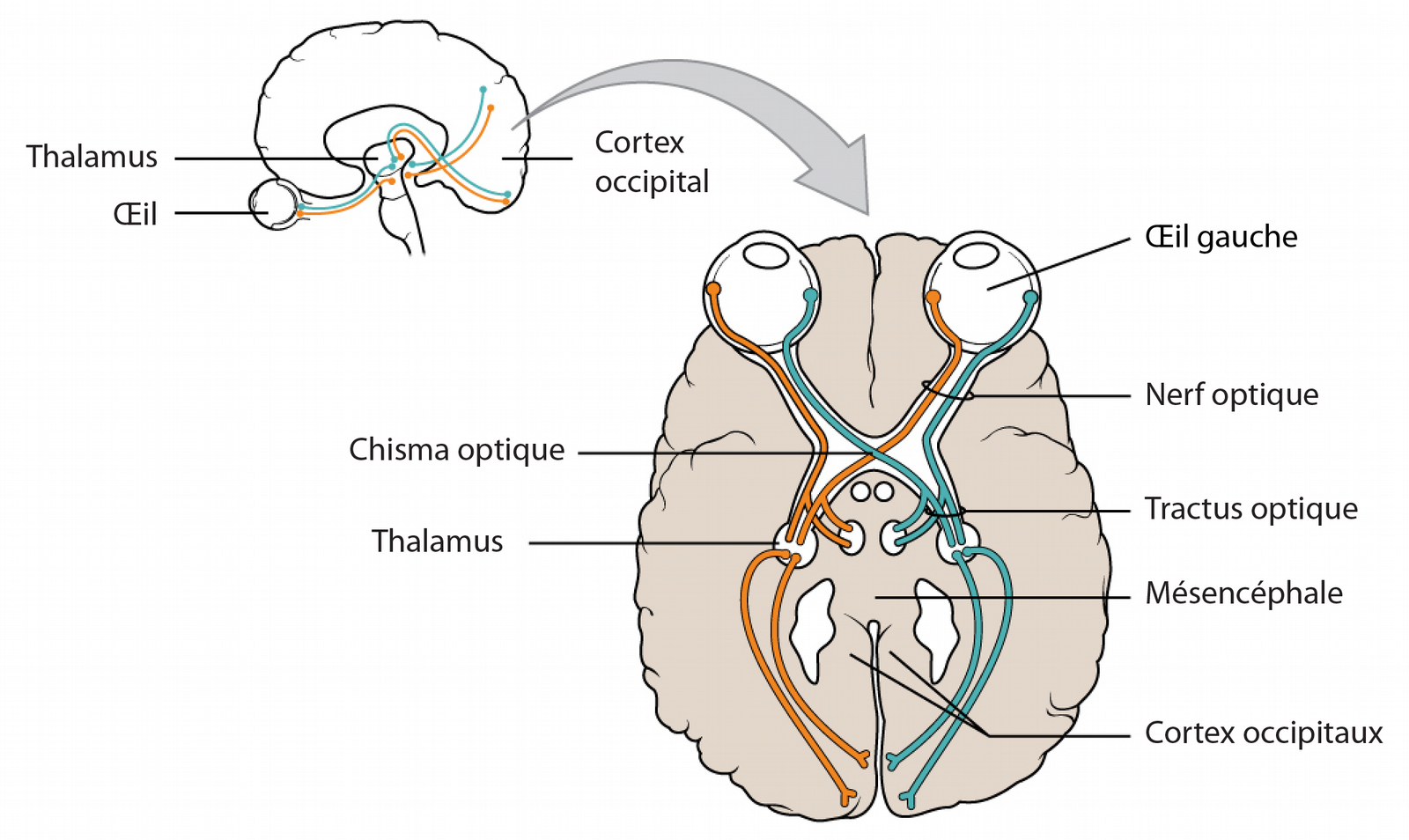
La vision est la capacité à détecter, intégrer et interpréter les rayons lumineux provenant de l’environnement. Partagée par de nombreuses espèces animales, cette faculté leur permet de s’orienter et d’interagir avec leur environnement. Chez les Vertébrés, les organes impliqués dans la vision sont les yeux et le cerveau (voir Figure 1). L’œil est parfois assimilé à un simple capteur tandis que le cerveau serait le lieu de reconstruction et d’interprétation de l’image (Gollisch & Meister, 2010). Pourtant la rétine, loin d’être un capteur passif, joue un rôle majeur dans le traitement de l’information visuelle.
La rétine fonctionne-t-elle comme le capteur d’un appareil photo ?
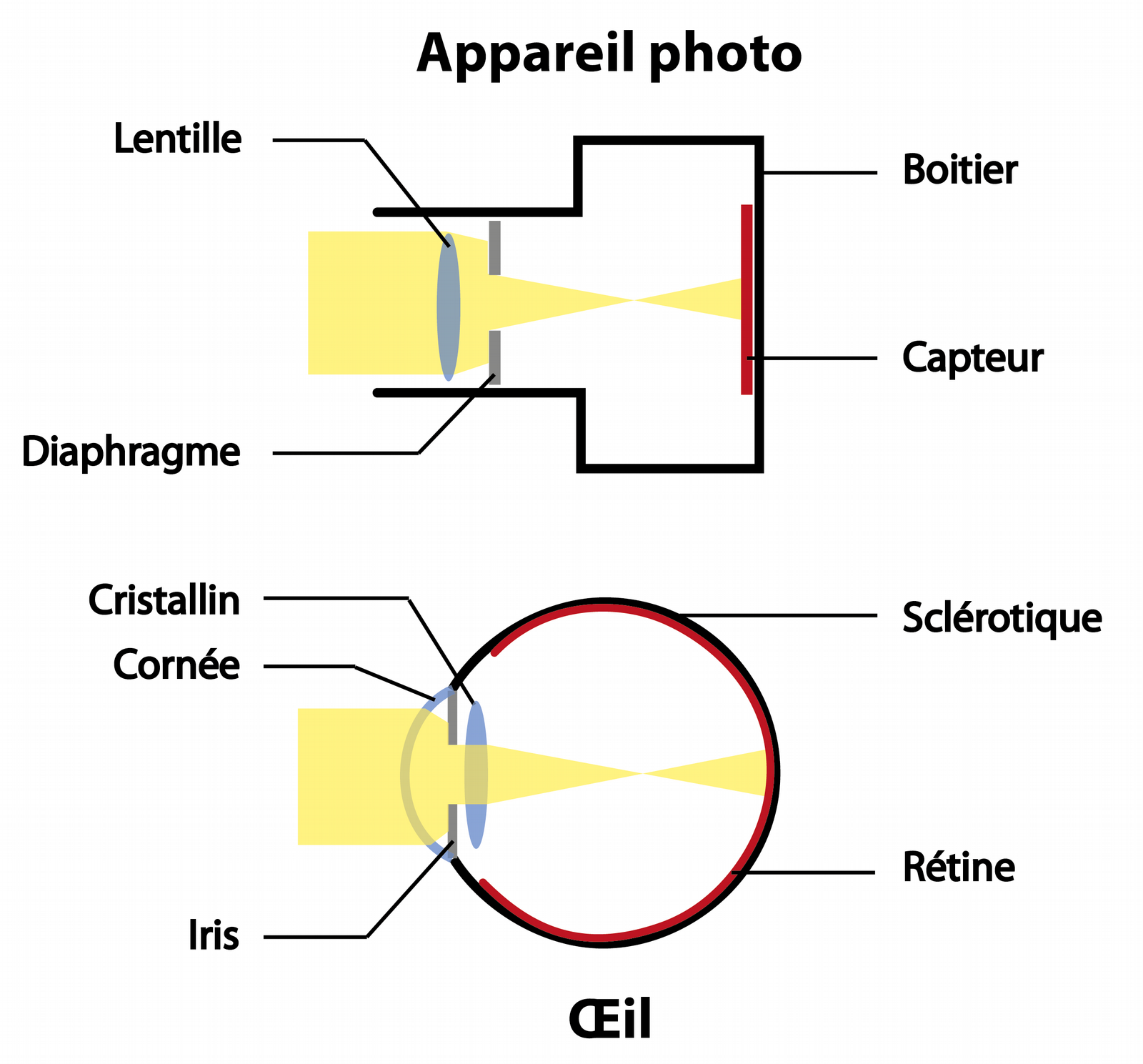
L’œil est parfois comparé à une caméra, ayant pour rôle de capturer une image brute. Ils partagent en effet des caractéristiques communes : la présence d’un jeu de lentilles (la cornée et le cristallin), et d’un diaphragme (l’iris) qui concentrent la lumière sur un capteur (la rétine ; voir Figure 2).
Le capteur d’une caméra numérique est composé d’une mosaïque de récepteurs capables de mesurer la quantité (l’intensité lumineuse) et la qualité (la longueur d’onde) de la lumière (voir Figure 3, B et D). Trois récepteurs, sensibles respectivement à la lumière bleue, rouge et verte, sont combinés pour produire un pixel, et l’assemblage de ces pixels sur l’ensemble du capteur permet de reconstruire l’image. Le nombre de récepteurs dans le capteur détermine directement la définition de l’image. Un plus petit nombre de récepteurs donnera une image mal définie, alors qu’un grand nombre de récepteurs permet d’avoir une image contenant plus d’informations.
La rétine de son côté est un fin tissu, composé principalement de neurones, qui couvre le fond du globe oculaire. Comme le capteur d’une caméra, la rétine possède une mosaïque de récepteurs, les photorécepteurs, qui transforment les radiations lumineuses du domaine visible (entre 370 et 730 nm chez l’être humain) en signaux électriques (voir Figure 3, A et C). Là encore, les récepteurs sont sensibles à différentes longueurs d’onde. Chez les humains les photorécepteurs sont de deux sortes : les cônes, séparés en trois types sensibles respectivement aux couleurs rouge, verte et bleue ; et les bâtonnets, qui sont impliqués lors des basses luminosités comme lors de la vision crépusculaire. Les signaux électriques produits par les photorécepteurs sont transmis à différentes régions du cerveau et notamment au cortex visuel primaire nécessaire à la vision consciente. Celui-ci a une organisation rétinotopique, c’est-à-dire que chaque secteur du cortex visuel correspond à un secteur de la rétine, et donc à un fragment du champ visuel. L’information visuelle est aussi transmise à d’autres parties du cerveau : l’hypothalamus pour recaler notre horloge interne sur le cycle diurne, le tronc cérébral pour participer aux mouvements des yeux et de la tête et aussi pour régler le diamètre la pupille.
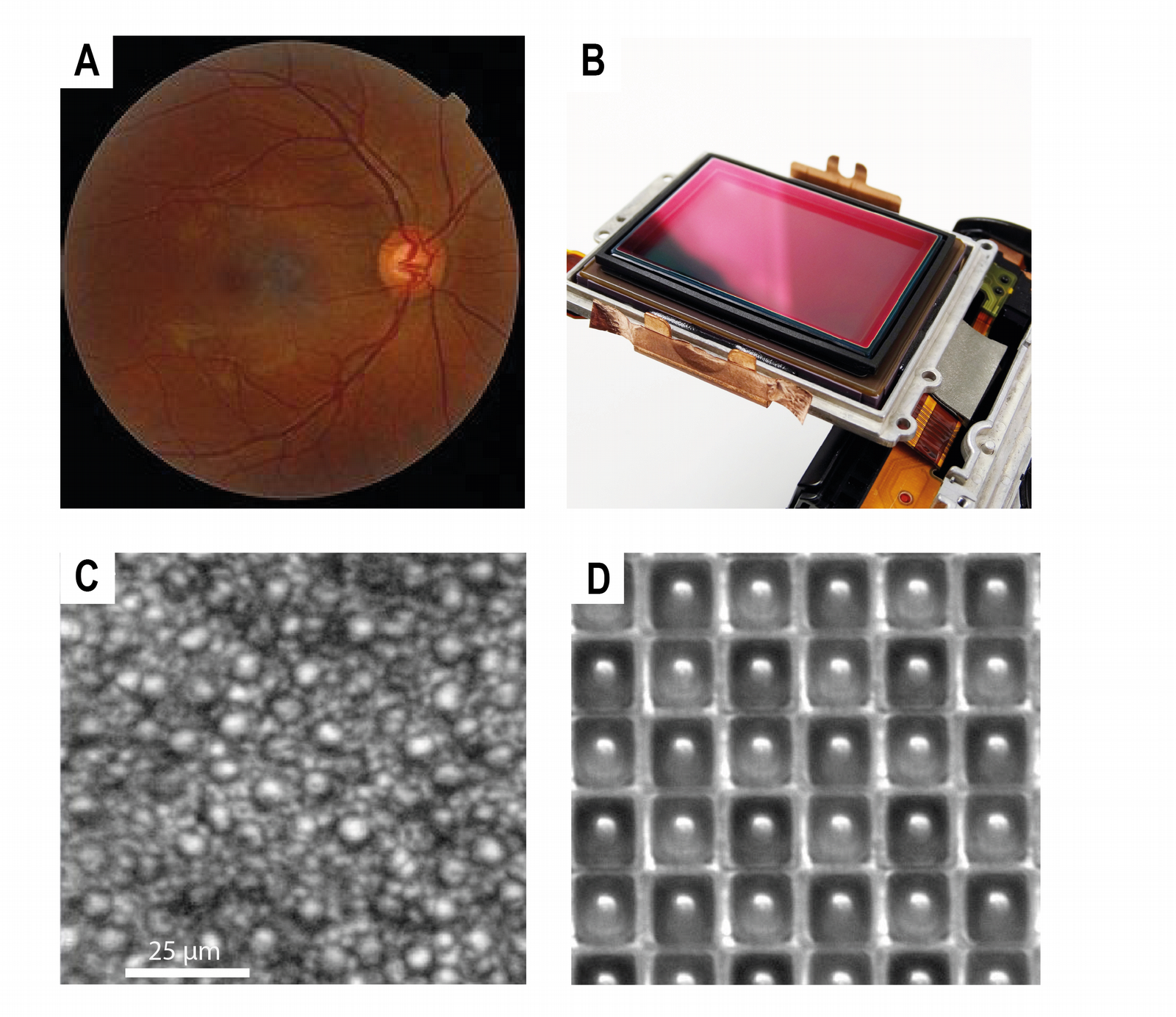
A. Rétine humaine. B. Capteur de caméra numérique. C. Mosaïque de photorécepteurs dans la rétine humaine. Les grandes cellules correspondent aux cônes, les petites aux bâtonnets. D. Mosaïque de récepteurs dans le capteur d’une caméra numérique.
Crédits : A : Ske, CC BY-SA, Wikimedia ; B : Pxfuel ; C : Burns, S. A et coll. (2019). Adaptive optics imaging of the human retina. Progress in retinal and eye research, CC BY-NC-ND ; D : Jackshencn, reproduit avec autorisation, The Landingfield
Étant donné ces similitudes, on pourrait penser que le cerveau fonctionne comme un ordinateur associé à une caméra, dont le capteur (la rétine) enregistre sous forme de pixels la quantité et la qualité de la lumière en chaque point du champ visuel grâce à une mosaïque de récepteurs. En réalité, la rétine constitue bien plus qu’un capteur traduisant une image en pixels. Elle est capable de réaliser des transformations complexes de l’information avant son envoi au cerveau.
Le traitement des informations visuelles par la rétine
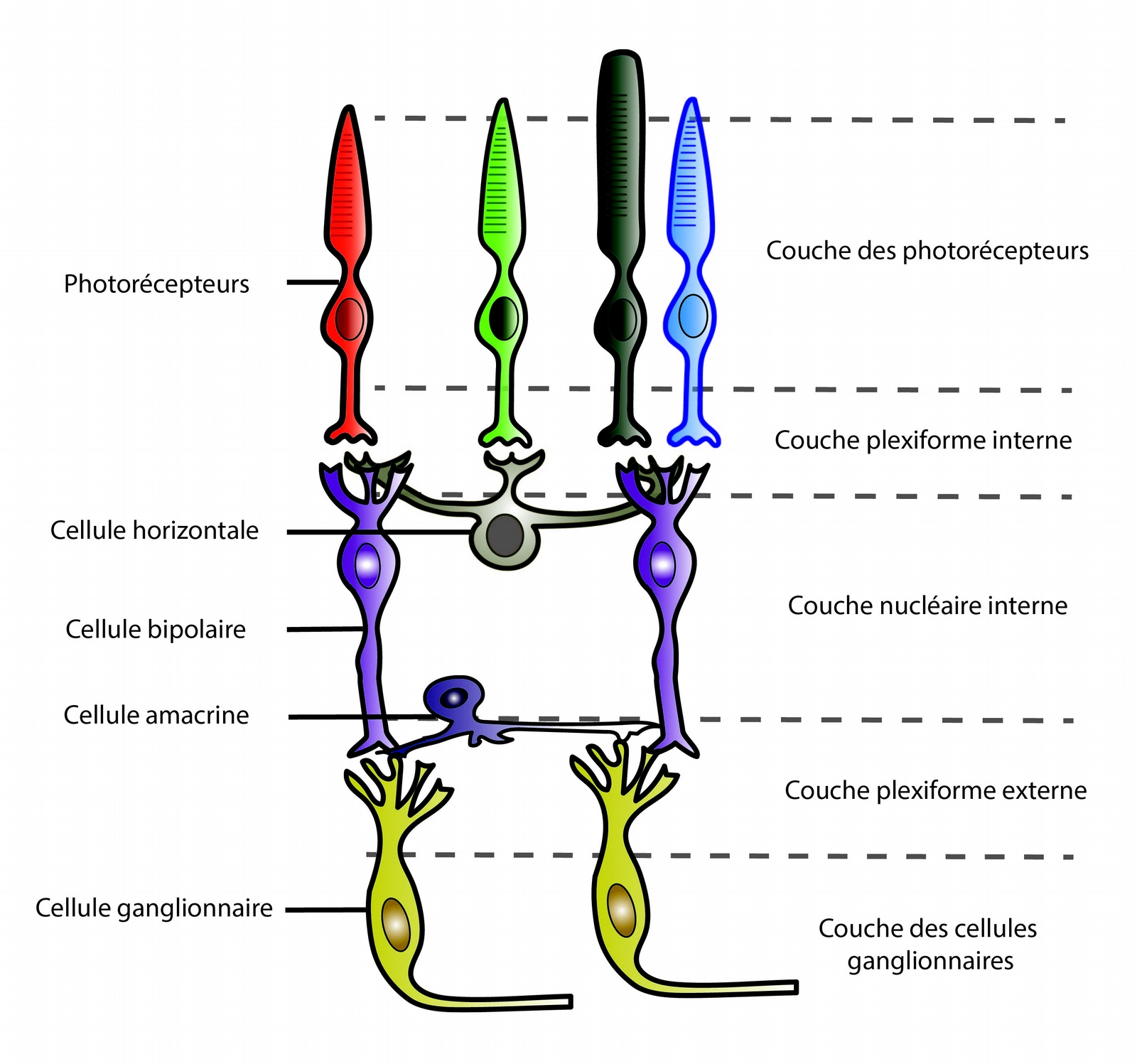
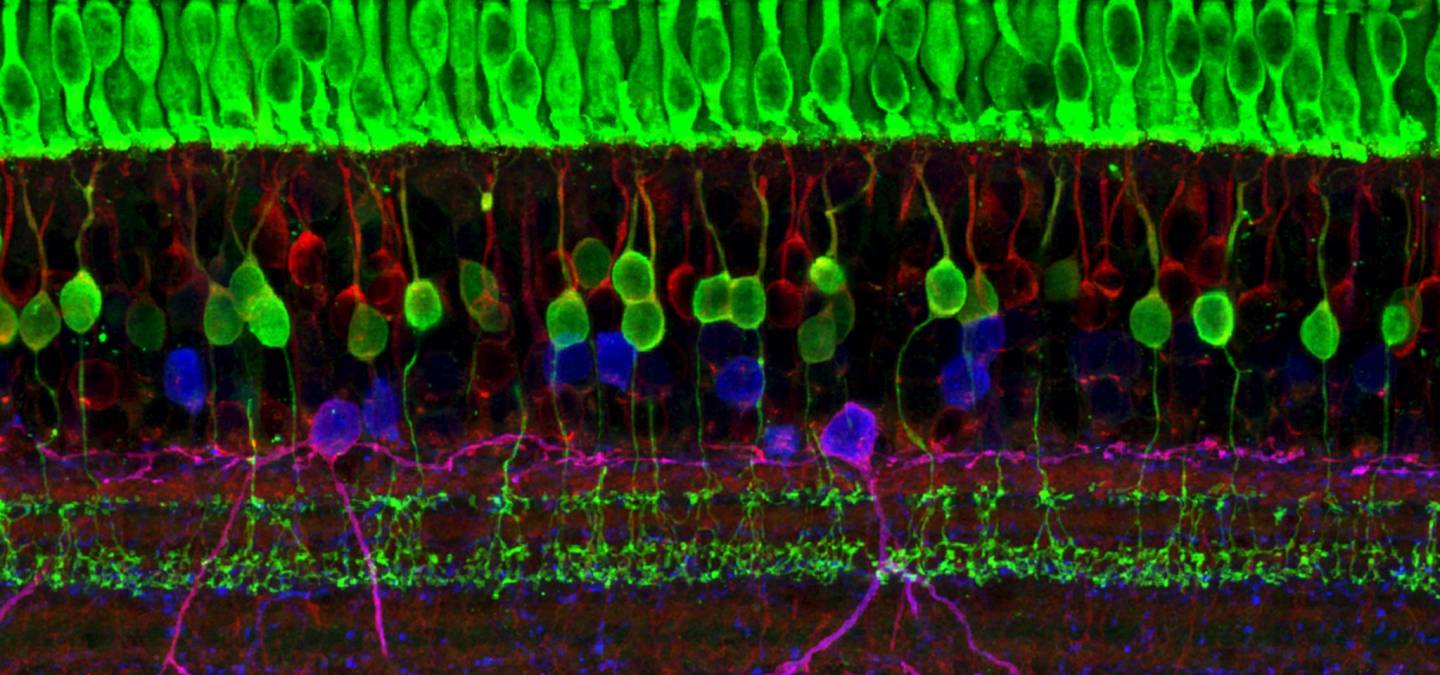
Bien que le nombre de sous-types cellulaires varie entre les espèces de Vertébrés, les grandes classes de cellules et l’organisation de la rétine en couches est largement conservée.
Dans la rétine, le signal électrique produit par les photorécepteurs n’est pas transmis directement au cerveau. En effet, la rétine présente une structure complexe : elle est composée de cinq grandes classes de cellules, organisées en cinq couches (voir Figure 4). C’est la dernière couche, formée par les cellules ganglionnaires, qui transmet les informations au cerveau via le nerf optique.
Le réseau de neurones allant des photorécepteurs aux cellules ganglionnaires joue un rôle primordial dans la vision. En effet, il permet de transformer l’information brute générée par les photorécepteurs en y distinguant plusieurs caractéristiques de l’image vue, comme les bordures, les couleurs et les mouvements. Ces différents canaux d’informations parallèles sont traités par des types de cellules ganglionnaires distincts (voir Figure 5). C’est à partir de ces informations prétraitées que le cerveau reconstruit l’image.
Les différents types de cellules ganglionnaires sont sensibles à différents aspects de l’image, comme les contours (type cellulaire 1) ou l’opposition de couleurs (types cellulaires 2 et 3). En réalité, ce n’est pas moins d’une vingtaine de types cellulaires différents qui sont impliqués dans le traitement rétinien de l’image chez l’être humain.
La rétine possède donc une mosaïque apparentée à des pixels, mais celle-ci est constituée par les cellules ganglionnaires. Considérons un cas simplifié, comprenant seulement trois types de cellules ganglionnaires, représentant respectivement les contours, l’opposition de couleur rouge-vert, et l’opposition de couleur bleu-jaune (Figure 5). Par opposition de couleur, on entend la comparaison de l’intensité des deux couleurs. Pour chaque fragment du champ visuel, la rétine envoie au cerveau non pas une seule valeur de luminosité, mais plusieurs informations :
- la présence de bordures, signalées par un contraste de luminosité important ;
- la comparaison vert-rouge ;
- et la comparaison bleu-jaune.
Des régions de la rétine, comprenant un exemplaire de chaque type de cellules ganglionnaires, peuvent donc être considérées comme des « super-pixels », envoyant au cerveau l’ensemble de ces informations.
Une autre façon de décrire l’organisation de la rétine consiste à considérer l’ensemble des cellules ganglionnaires d’un même type sur toute sa surface. Ce groupe de cellules forme un film rétinien (Roska, 1999), collectant un seul type d’information visuelle (par exemple les contours de l’image), sur l’ensemble du champ visuel (voir Figure 6). La superposition de tous les films rétiniens permet de reconstituer l’image complète.
En réalité, la rétine est constituée de beaucoup plus que trois types de cellules ganglionnaires. En combinant des informations sur leur anatomie et leur signature électrophysiologique, plus de 40 types cellulaires ont pu être décrits chez la souris (Baden et coll., 2016). Bien que le rôle exact de chacun dans la perception finale de l’image soit encore mal compris, chaque type cellulaire présente une sensibilité particulière à différents aspects de l’image. Par exemple, le type nommé W3 (ou G10) est sensible aux différences de contraste locales, pouvant servir de base pour la détection de contours, alors que le type G14 est sensible à la présence de mouvements dans les directions cardinales (mouvement vertical ou horizontal). D’autres types cellulaires sont impliqués dans l’anticipation des mouvements (Berry et coll., 1999), ou encore dans la détection des changements de direction (Schwartz et coll., 2007). Chez l’être humain, on estime à une vingtaine le nombre de types de cellules ganglionnaires, mais une fonction n’a été décrite avec précision que pour la moitié environ (Dacey, 2011).

Les cercles représentent la mosaïque de cellules ganglionnaires vues du dessus, leur couleur indiquant leur type. À gauche, chaque image représente le film rétinien transmis par chacun des trois types de cellules ganglionnaires illustré en figure 5. À droite, les images montrent comme l’assemblage de ces films rétiniens, sur une partie ou l’ensemble du champ visuel, permet de reconstituer l’image vue.
La rétine est donc capable de traitements complexes de l’information visuelle. À partir du milieu du xxe siècle, un cadre théorique a également été proposé pour comprendre l’origine de ce type de traitements. La principale hypothèse de cette théorie est que la rétine réalise un codage efficace de l’information (Attneave, 1954).
Le codage efficace réalisé par la rétine
L’organisation globale de la rétine, ainsi que les grandes classes de cellules qui la composent, sont très conservées au sein des Vertébrés, alors que les schémas d’organisation plus fins peuvent varier de façon importante entre les espèces. Mais la plupart des structures semblent conférer à la rétine la même capacité à effectuer des traitements complexes de l’information, en particulier l’extraction parallèle de caractéristiques (Baden et coll., 2018). De plus, ce type de structure permet à la rétine de réaliser un codage dit efficace. Il s’agit d’une forme de compression et de tri de l’information.
D’un point de vue théorique, une rétine « idéale » transmettrait un maximum d’information visuelle pertinente au cerveau. En pratique cependant, il s’opère nécessairement un compromis entre la quantité et la qualité de l’information visuelle transmise au cerveau, et le coût métabolique qu’ont l’acquisition, le traitement et le transport de cette information. En effet, les systèmes nerveux sensoriels sont très coûteux en énergie ; le maintien du potentiel de membrane des neurones et l’émission de potentiels d’action implique une machinerie cellulaire complexe et consommatrice d’ATP. La charge métabolique liée au fonctionnement de la vision peut être réduite en restreignant le nombre de cellules, ce qui permet aussi de restreindre la taille du nerf optique et donc de répondre à une contrainte anatomique. Elle peut également être diminuée en limitant l’activité de ces cellules, correspondant au nombre de potentiels d’action émis.
Dans la rétine, une seule cellule ganglionnaire est connectée à plusieurs photorécepteurs par l’intermédiaire d’une ou plusieurs cellules bipolaires (Gollisch & Meister, 2010). Ce schéma de connexion est désigné par le terme convergence, et limite le nombre de cellules ganglionnaires. Dans la rétine des Primates, il n’y a que dans une petite région centrale permettant la vision précise des détails, la fovéa, que l’on retrouve une cellule ganglionnaire par photorécepteur.

On observe une convergence, c’est à dire une diminution du nombre de cellules des photorécepteurs jusqu’aux cellules ganglionnaires. Dans le même temps, on observe une diversification des types cellulaires, passant de 4 pour les photorécepteurs à plus de vingt pour les cellules ganglionnaires. Ces chiffres sont valables pour l’être humain (Kolb, 1995) ; le nombre de cellules et de types cellulaires peut varier entre les espèces (Baden et coll., 2018).
Si le nombre de cellules diminue d’un facteur 100 entre les photorécepteurs et les cellules ganglionnaires, le nombre de types cellulaires, traitant des informations différentes, passe quant à lui de quatre à plusieurs dizaines (voir Figure 7).
La charge métabolique est également réduite par la restriction de l’activité des cellules ganglionnaires. Il a été montré que la rétine est un détecteur de changements, qui ne transmet au cerveau que les informations pertinentes, surprenantes, en réduisant les corrélations temporelles et spatiales dans les images perçues (Pitkow & Meister, 2012). En effet, il serait coûteux et peu efficace que face à une scène invariable, chaque cellule ganglionnaire transmette en continu la même information au cerveau. En réalité, les cellules ganglionnaires ne signalent que les changements qui surviennent dans leur fragment de champ visuel. Les scènes visuelles étant globalement statiques, c’est-à-dire fortement corrélées temporellement, l’activité de cellules signalant seulement les changements est donc fortement limitée.
Par ailleurs, les scènes visuelles sont fréquemment composées de surfaces uniformes, il est donc peu efficace pour deux cellules voisines de transmettre toutes les deux la même information. Grâce à des connexions horizontales au sein de la rétine, les cellules ganglionnaires sont capables de comparer l’information contenue dans leur fragment de champ visuel à celle contenue dans les fragments à proximité, pour ne signaler que les informations qui se démarquent spatialement de leur environnement. Cette réduction des corrélations spatiales réduit également l’activité globale des cellules ganglionnaires dans la rétine. (voir Figure 8).
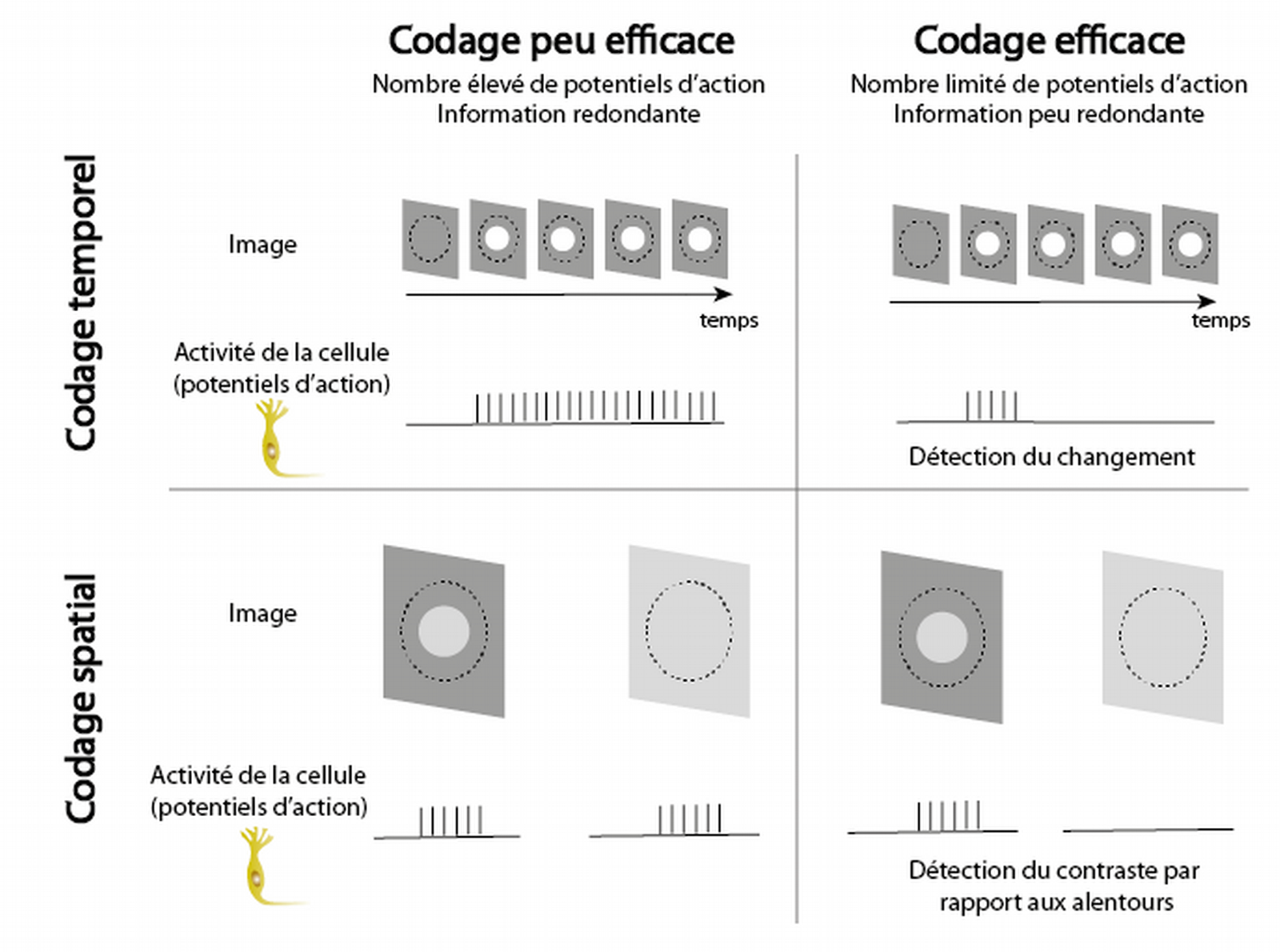
Le cercle en pointillé indique la région du champ visuel à laquelle le neurone représenté correspond. Le codage spatial et temporel des fragments d’images par chaque cellule ganglionnaire peut être plus ou moins efficace. La limitation du nombre de potentiels d’action en détectant les changements et les contrastes plutôt que la luminosité en continu permet de rendre le codage plus efficace.
Conclusion
Le circuit allant des photorécepteurs aux cellules ganglionnaires permet de transformer et de trier l’information visuelle pour en extraire des caractéristiques (contours, couleurs, mouvement…), transmises au cerveau par différents types de cellules ganglionnaires. De plus, le codage efficace réalisé par la rétine, consistant à trier et compresser ces informations en détectant les changements et les contrastes, permet aux organismes de répondre à des contraintes métaboliques et anatomiques. La rétine est donc plus qu’un simple capteur, où des traitements complexes de l’information sont déjà réalisés.
L’identification des différents types de cellules ganglionnaires, des caractéristiques qu’elles peuvent extraire de l’image, et la compréhension de leur rôle de détecteurs de changement constituent un domaine de recherche très actif, qui présente des enjeux à la fois fondamentaux et cliniques. En effet, une des solutions envisagées pour traiter de nombreuses causes de cécité consiste la mise au point de prothèses de rétine. La transmission des informations visuelles depuis ces rétines artificielles vers le cerveau nécessite de mieux comprendre les mécanismes mis en jeu dans le traitement de ces informations.
Références
- Attneave, F. (1954). Some informational aspects of visual perception. Psychological Review, 61(3), 183–193. https://doi.org/10.1037/h0054663
- Baden, T., Berens, P., Franke, K., Rosón, M. R., Bethge, M., & Euler, T. (2016). The functional diversity of retinal ganglion cells in the mouse. Nature, 529(7586), 345–350.
- Baden, T., Schubert, T., Berens, P., & Euler, T. (2018). The functional organization of vertebrate retinal circuits for vision. Oxford Research Encyclopedia of Neuroscience.
- Berry, M. J., Brivanlou, I. H., Jordan, T. A., & Meister, M. (1999). Anticipation of moving stimuli by the retina. Nature, 398(6725), 334–338. https://doi.org/10.1038/18678
- Dacey, D. (n.d.). Origins of perception : Retinal ganglion cell diversity and the creation of parallel visual pathways. 66.
- Gollisch, T., & Meister, M. (2010). Eye smarter than scientists believed : Neural computations in circuits of the retina. Neuron, 65(2), 150–164.
- Kolb, H. (1995). Facts and figures concerning the human retina.
- Lettvin, J. Y., Maturana, H. R., McCulloch, W. S., & Pitts, W. H. (1959). What the Frog’s Eye Tells the Frog’s Brain. Proceedings of the IRE, 47(11), 1940–1951. https://doi.org/10.1109/JRPROC.1959.287207
- Pitkow, X., & Meister, M. (2012). Decorrelation and efficient coding by retinal ganglion cells. Nature Neuroscience, 15(4), 628.
- Roska, F. W., Botond. (n.d.). Des films sur la rétine. Pourlascience.fr ; Pour la Science. Retrieved May 17, 2020, from https://www.pourlascience.fr/sd/neurosciences/des-films-sur-la-retine-3023.php
- Schwartz, G., Taylor, S., Fisher, C., Harris, R., & Berry, M. J. (2007). Synchronized Firing among Retinal Ganglion Cells Signals Motion Reversal. Neuron, 55(6), 958–969. https://doi.org/10.1016/j.neuron.2007.07.042



{kind=link}
{kind=link}