

Cet article fait partie du dossier "La morphogenèse végétale et l'environnement". Il présente l'influence de trois facteurs de l'environnement entraînant une modification du port de la plante dans l'espace, l'action du facteur étant qualifiée de dirigée. Il s'agit de l'influence de la lumière, de la gravité et du contact, dont les réponses de la plante se nomment, respectivement, phototropisme, gravitropisme et thigmotropisme.
Le phototropisme
Mise en évidence chez le coléoptile de blé
Le coléoptile d’une germination de blé est un bon matériel pour mettre en évidence la réaction phototropique. Les photographies suivantes montrent des barquettes de germinations réalisées à l’obscurité (à gauche). Les barquettes de droite ont été soumises pendant 5 heures à un éclairement directionnel de faible intensité. Les réactions diffèrent selon l’âge des germinations.

Colonne de gauche: coléoptiles cultivés à l'obscurité et colonne de droite : coléoptiles soumis pendant 5 heures à un éclairement directionnel de faible intensité.
Ligne 1: Les germinations sont très jeunes. Les coléoptiles sont en pleine phase de croissance. Aucun d’entre eux n’est percé par la feuille. Ils se penchent vers la lumière venant de droite. La réaction phototropique est générale.
Ligne 2: Les germinations sont plus âgées. Les coléoptiles sont en fin de croissance. Certains d’entre eux sont percés par la feuille. Certains se penchent vers la lumière venant de gauche. La réaction phototropique est moins générale mais encore bien visible.
Ligne 3: Les germinations sont âgées. Les coléoptiles ont terminé leur croissance. Tous ont été percés par la feuille. Ils ne se penchent pas vers la lumière venant de droite. La réaction phototropique ne se réalise plus.
Cette expérience montre à l’évidence que la réaction phototropique est en relation avec la croissance. L’expérience suivante permet de préciser certains points.
Expérimentation et interprétation
Voici, schématisé, un protocole expérimental simple permettant de mettre en évidence la réaction phototropique d’une plante, sur l’exemple des coléoptiles de blé.
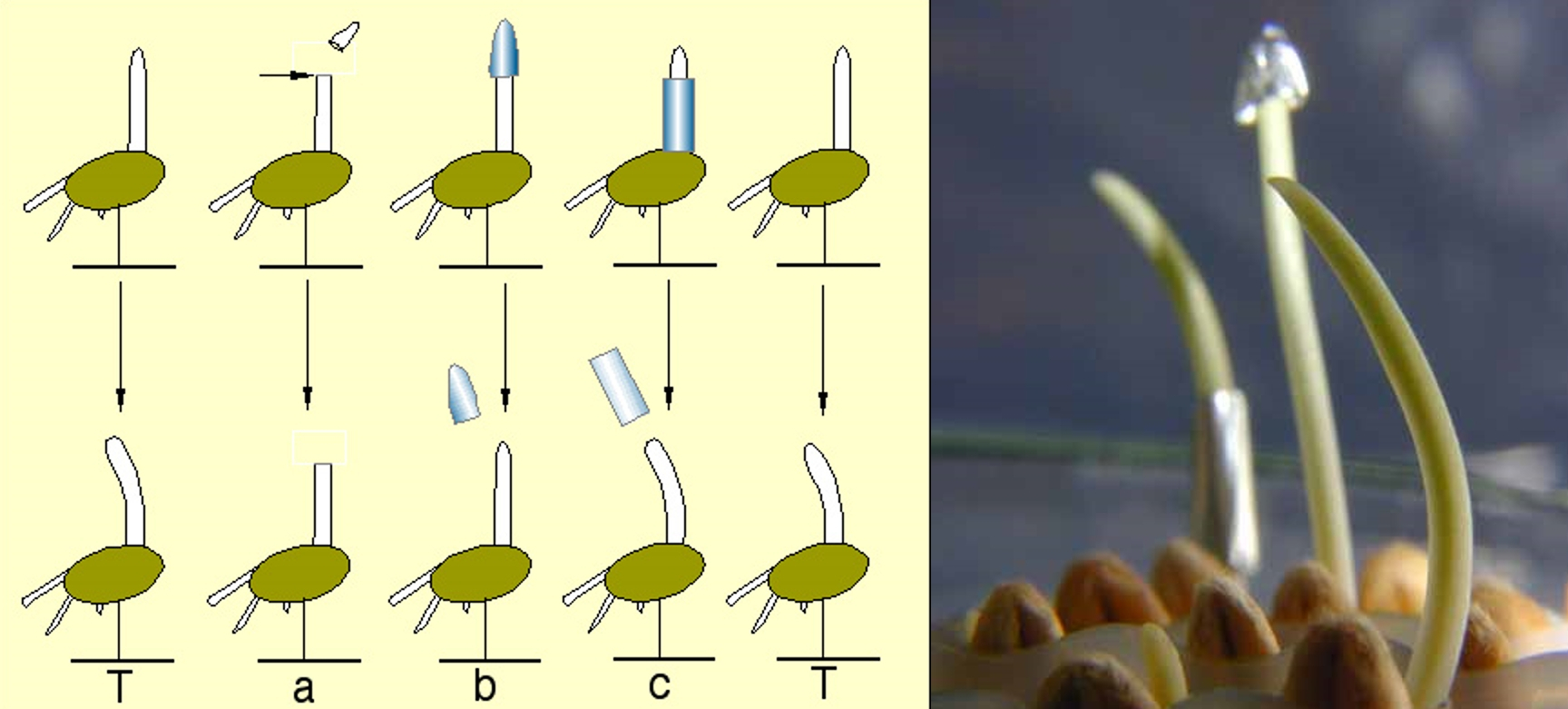
À gauche: schéma du protocole expérimental. Les coléoptiles sont soit intacts, soit décapités, soit recouverts d’un capuchon de papier aluminium, soit habillés d’un petit cylindre de papier aluminium. Ils sont ensuite soumis pendant 3 heures à un éclairement directionnel de faible intensité (venant ici de gauche).
À droite: photographie du résultat de cette expérience.
L’expérience précédente met en évidence le rôle fondamental de l’extrémité du coléoptile dans la réponse à la lumière. S’il est présent le coléoptile présente une courbure en direction de la lumière. S’il est absent ou recouvert d’un capuchon d’aluminium, on n’observe aucune courbure vers la lumière. Le fait de cacher, par un manchon d’aluminium, la base du coléoptile, n’empêche pas la courbure vers la lumière. Cette zone ne semble pas sensible à la lumière.
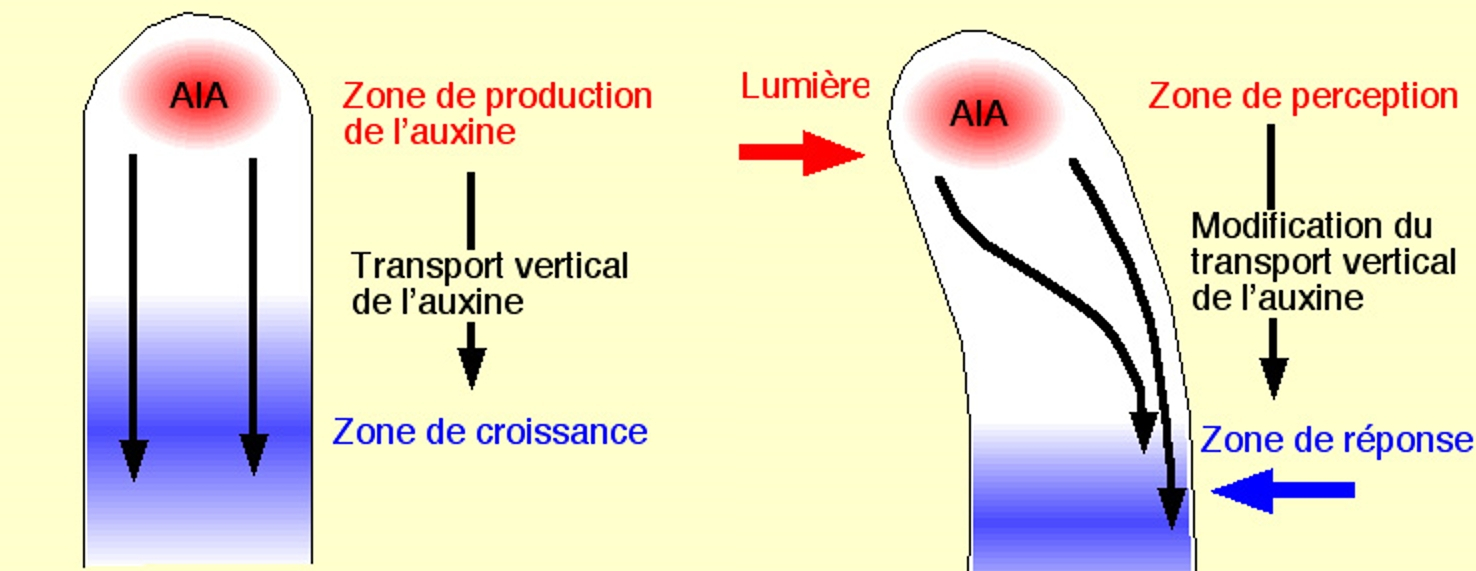
La réponse est une courbure du coléoptile due à une répartition différente de l’hormone auxine (voir l’article « Comment grandit une cellule végétale ?»), entre le côté exposé à la lumière et celui exposé à l’obscurité.
Le gravitropisme
Mise en évidence
La gravité est certainement le facteur de l’environnement qui influence le plus la morphogenèse de toutes les plantes. Ce facteur étant omniprésent dans la biosphère et, ne pouvant être modulé, il peut ne pas être perçu par l’observateur. Il est un lieu commun de constater que les plantes poussent selon un axe vertical mais cela apparaît « normal » et on a tendance à ne pas réfléchir à l’origine de ce phénomène.
Les deux images suivantes montrent 2 états d’une plantule de soja que l’on a couchée et solidement fixée à la base de l’hypocotyle, 5 heures séparant les deux photographies.

La plantule de soja est maintenue couchée, fixée par la base de l'hypocotyle.
À gauche: début de l'expérience
À droite: fin de l'expérience, 5 heures plus tard
Vidéo accélérée montrant l'effet du gravitropisme sur une plantule de soja
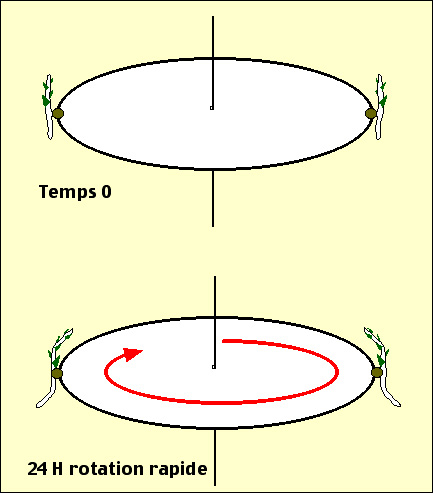
Pour étudier le gravitropisme on peut utiliser le dispositif expérimental ci-contre. Après 24 heures de ce traitement, les plantules sont courbées. Les racines s’orientent vers l'extérieur du disque et les tiges en sens inverse. L’examen détaillé montre que la courbure s’est effectuée au niveau des zones de croissance.
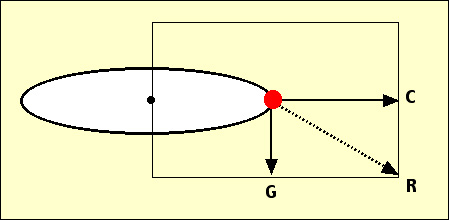
Les deux forces en présence, la gravité (G) et l’accélération centrifuge (C), ont pour résultante une force (R) dirigée vers l'extérieur et vers le bas. L’orientation nouvelle de croissance se réalise selon cette résultante, positivement pour la racine, négativement pour la partie feuillue.
On peut alors se poser la question du lieu de perception de la gravité.
Quel est le lieu de perception de la gravité chez une plante ?
On peut démontrer par des expériences simples (mais difficiles à réaliser hors laboratoire) que la coiffe de la racine est nécessaire à la réaction gravitropique.
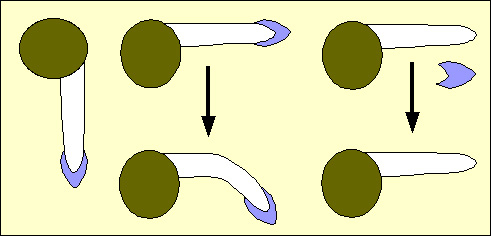
À gauche : graine germée présentant une jeune racine, en position normale, dirigée vers le bas.
Au centre : graine dont la racine (intacte) est placée en position horizontale, on observe une courbure vers le bas
À droite : racine dont la coiffe est ôtée, placée en position horizontale, on n’observe pas de courbure vers le bas.
L’examen de l’extrémité de la racine montre que les cellules de la coiffe, contrairement aux cellules du méristème, sont riches en amyloplastes. Ces organites, contenant de l’amidon, sont les plus denses de la cellule et se répartissent dans celle-ci en fonction de la gravité. Les deux images suivantes donnent, sous une représentation très schématique, la position des amyloplastes dans des racines placées verticalement ou horizontalement.
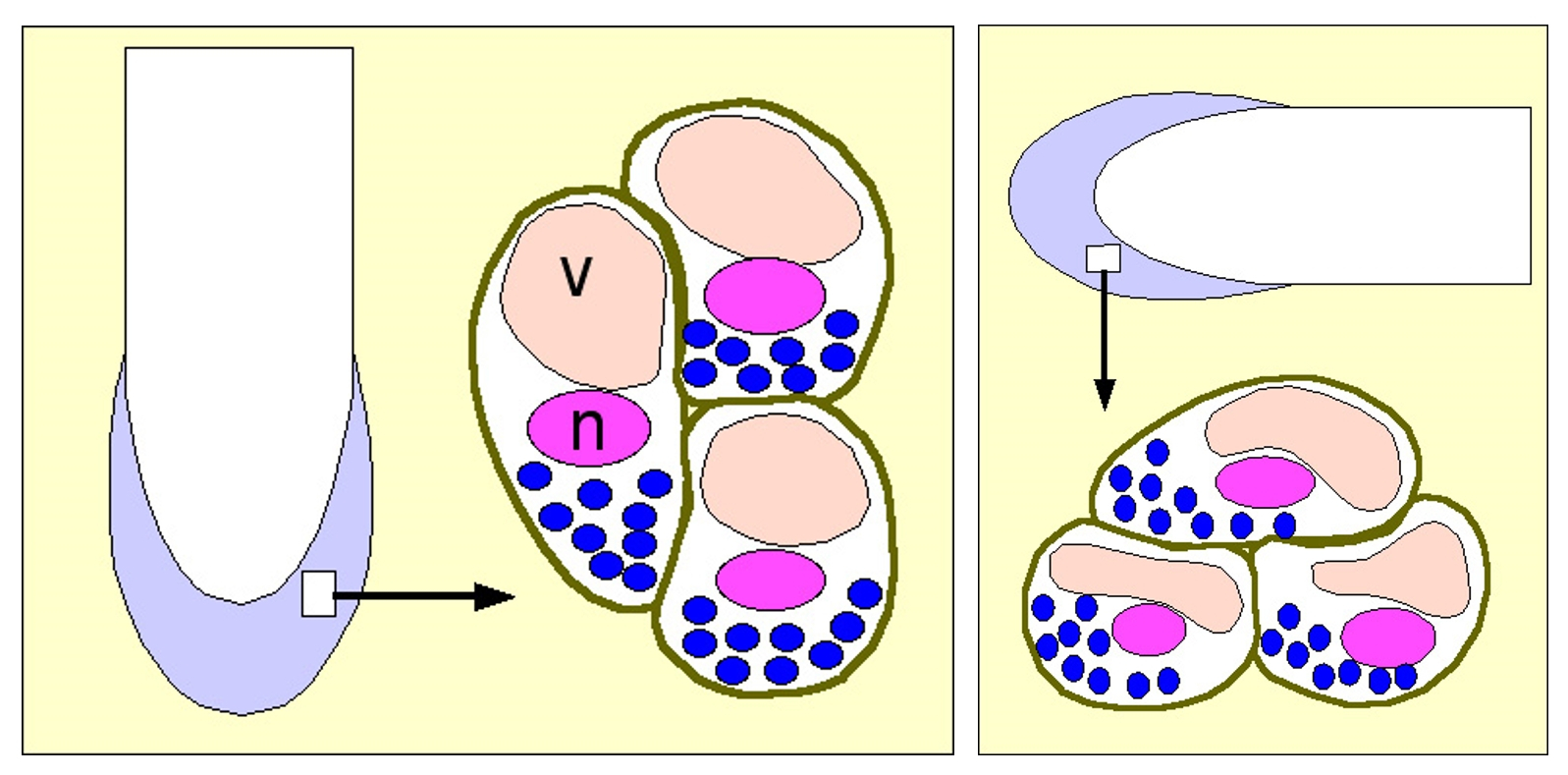
Les grains d’amidon (ici en bleu foncé) se positionnent dans la cellule en fonction de la gravité et jouent le rôle de statolithes. Les statocystes sont ces cellules renfermant des statolithes.
v : vacuole ; n : noyau.
Les amyloplastes sont les seuls organites à être affectés physiquement par la gravité. On peut penser qu’ils jouent un rôle essentiel dans la perception de la gravité. On les considère comme des statolithes et les cellules de la coiffe qui les contiennent comme des statocystes.
Quel est le mécanisme qui permet à la racine d’orienter sa croissance au cours de la stimulation gravitropique ?
Mécanisme de la réponse à la gravité
Trois prérequis sont nécessaires :
-
La croissance de la racine est stimulée par des doses très faibles d’auxine. Les doses qui stimulent la croissance des tiges sont inhibitrices pour les racines.
-
Il semble que l’auxine synthétisée au niveau des apex caulinaires et des jeunes feuilles descende dans les racines par le cylindre central. Elle serait redistribuée au niveau de la coiffe et remonterait dans les racines, avec des concentrations faibles, par la zone corticale.
-
Les cellules sensibles à l’auxine (pour la croissance) seraient les cellules de la zone corticale.
Les schémas ci-dessous tentent d’expliquer comment une stimulation gravitropique perçue par la coiffe peut se traduire par une croissance différentielle de la zone de croissance de la racine.
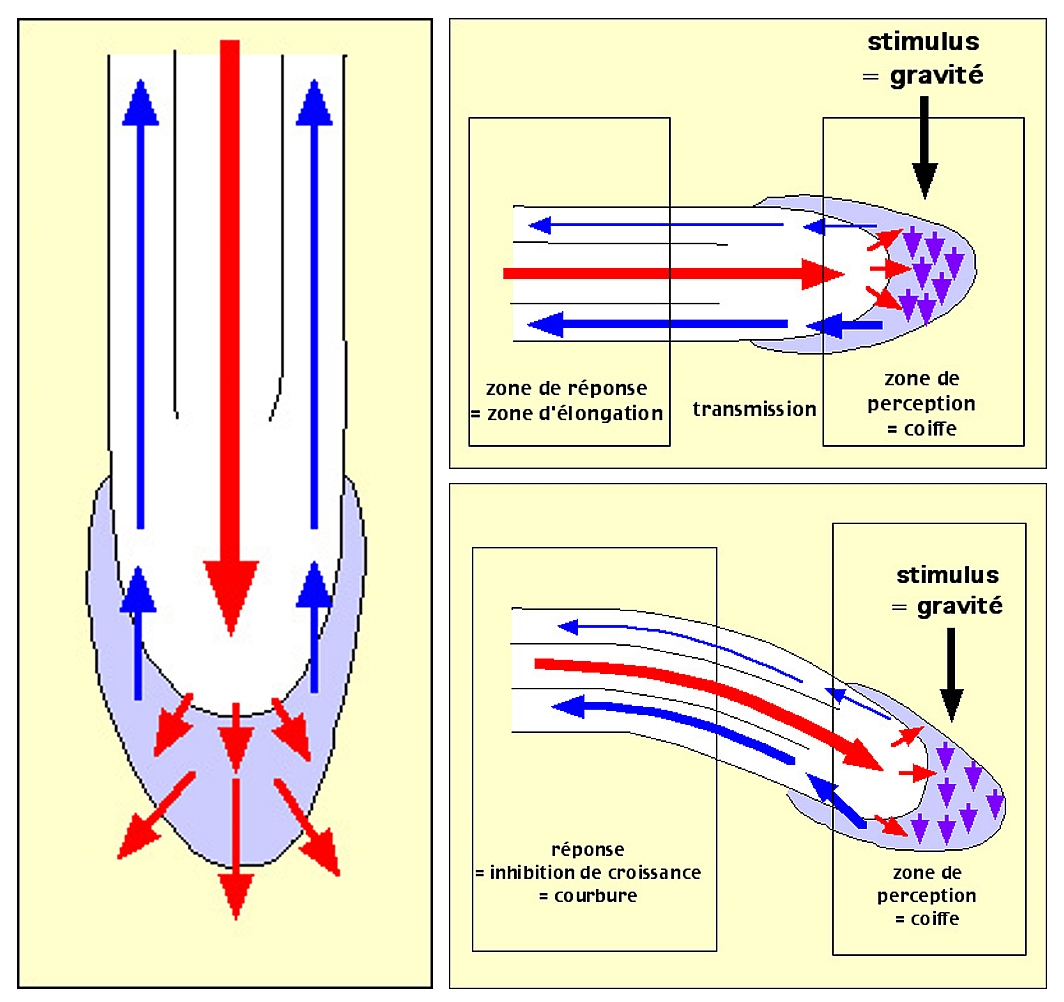
Coiffe de la racine en bleu-violet, trajets de l'auxine matérialisés par des flèches rouges ou bleues, deux cadres localisant la zone de perception de la gravité et la zone de croissance, ou élongation, de la racine.
Lorsque la racine est placée verticalement, l’auxine remonte de manière égale tout autour de la racine et les réactions de la zone de croissance sont équivalentes sur tout le pourtour : la racine grandit verticalement.
Lorsque la racine est placée horizontalement, la réaction initiée par le déplacement des statolithes provoque un déplacement de l’auxine selon la position relative à la gravité. L’auxine remonte dans la partie corticale de manière inégale et provoque au niveau de la zone d’élongation une croissance différentielle par inhibition de croissance du côté où se trouvaient les statolithes (l’auxine ayant une action inhibitrice sur la croissance des racines, voir plus haut).
Deux remarques :
-
Le mot « perception » est employé ici avec un sens qui pourrait faire sursauter nos collègues neurologistes puisqu’en effet, la réception du stimulus « gravité » ne fait pas l’objet d’une analyse sensorielle.
-
L’analyse de la réaction gravitropique est bien comprise pour les racines. On sait peu de chose de ces mécanismes chez les tiges.
Circumnutation et thigmotropisme
Outre des facteurs comme la lumière et la gravité, d’autres facteurs de l’environnement peuvent intervenir pour modifier le comportement des plantes. Il en est ainsi du simple contact d’un objet étranger. Ceci peut être observé de manière très spectaculaire chez les plantes volubiles.
Chacun a déjà pu observer des plantes volubiles s’enroulant autour d’un tuteur. Le mécanisme est complexe et fait appel à au moins deux mécanismes unitaires indépendants : la circumnutation et le thigmotropisme.
La nutation correspond au fait que tous les apex de plantes (tiges ou racines) décrivent, en croissant, une spirale. Celle-ci a en général une très faible amplitude et seules des méthodes cinématographiques accélérées peuvent la mettre en évidence. C’est un phénomène intrinsèque indépendant de l’environnement.
Chez les plantes volubiles, ce mouvement peut être de très grande amplitude. Il est appelé circumnutation. On dit communément, avec finalité, que la plante « recherche » un tuteur. C’est pourquoi ce type de mouvement est souvent appelé mouvement d’exploration. En fait, sans rechercher une finalité que la plante ne peut imaginer, ce mouvement permet de balayer une grande partie de l’espace et augmente ainsi la possibilité de rencontrer un support. Ce type de mouvement fait partie du phénotype normal de l’individu et il est fixé génétiquement.
Les images suivantes montrent 3 états d’un plan de haricot, 30 minutes séparant chaque image.

Trois images de l'extrémité d'une tige de haricot prises à t = 0 à gauche, t = 30 mn au centre et t = 1 h à droite
Vidéo accélérée de la circumnutation d'un plan de haricot
Le thigmotropisme (ou haptotropisme) est un tropisme dont le stimulus est le contact. Le contact d’un objet au niveau d’une zone de croissance peut provoquer une inhibition locale de croissance. Ceci se traduit alors par une courbure dirigée vers l’objet. Cette réaction est particulièrement remarquable chez les tiges de plantes volubiles et chez les vrilles.
Le mouvement intrinsèque de circumnutation a permis de rencontrer un tuteur. Le thigmotropisme développé au contact du tuteur permet de s’enrouler précisément autour de lui.
Les images suivantes montrent deux états d’une jeune tige de liseron s’enroulant autour d’un tuteur, cinq heures séparant les deux photographies.

À gauche : début de l'observation à t = 0 et à droite : fin de l'observation à t = 5 h
L’association de ces deux phénomènes (et de bien d’autres) permet à certaines plantes de développer des morphologies particulières pour lesquelles l’impact des conditions extérieures modifie ou prolonge les capacités intrinsèques de l’individu.




