Cet article présente les mécanismes généraux par lesquels une plante acquiert sa forme (phénotype) par expression de son programme génétique (génotype). Cette expression est contrôlée par un grand nombre de facteurs de l'environnement : les influences de la lumière et de la gravité sont ici mises en relation avec le phénotype végétal, à partir d'observations des ramifications de différents arbres et d'études expérimentales sur des germinations.
Le port des végétaux ou comment s’établit la forme d’un végétal ?
De simples observations dans la nature montrent à l’évidence que l’on peut reconnaître une espèce à sa silhouette, à son port. Ceci est particulièrement évident pour les arbres. L’acquisition du port est dictée par des phénomènes physiologiques précis, déterminés génétiquement pour chaque espèce, avec des variations parfois notables entre les individus.
Cependant, les facteurs de l’environnement peuvent agir profondément sur ces processus morphogènes. C’est naturellement au cours de son développement que la plante acquiert sa forme définitive. Quelques observations concernant diverses modalités de ramification permettent de poser quelques jalons pour comprendre l’établissement de la morphogenèse.
Observations
La simple observation de deux arbres d’espèces différentes, un chêne et un pin, permet de constater des différences fondamentales dans leurs silhouettes. Ces différences sont assez importantes pour permettre de les reconnaître. Pourtant, il existe aussi des ressemblances puisque l’on affecte sans ambiguïté à ces deux espèces, le type « arbre ».

Silhouette d’un chêne (à gauche) et de pins (à droite)
C’est au niveau de la ramification que les différences les plus fondamentales apparaissent.
Acrotonie
Le port des arbres se distingue de celui des buissons par la croissance privilégiée des bourgeons les plus terminaux. C’est ce que l’on appelle l’acrotonie.
L’observation de deux pins, à des stades de développement différents, montre bien ce phénomène. Il est commun, au moins pendant une partie du développement, à tous les arbres.

L’acrotonie correspond au développement privilégié des bourgeons les plus terminaux,
ici sur l’exemple d’un pin à deux stades de son développement.
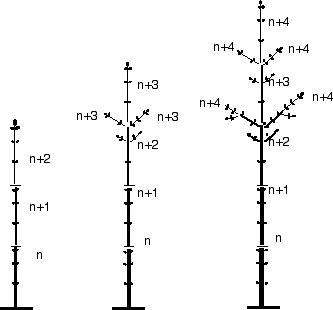
Ce schéma représente le développement d’un jeune arbre par acrotonie, sur 5 ans.
Dans l’exemple schématisé ci- contre, pendant les premières années de développement, les ramifications n’ont pas subsisté. Seul l’axe principal s’est développé (établissement d’un tronc).
À partir de l’année n+2 (dans ce cas de figure), à la fin de l’hiver, le bourgeon terminal et ses voisins latéraux les plus proches, sont les mieux développés. Ce sont ces bourgeons qui donneront des rameaux l’année suivante.
Le bourgeon terminal permettra de poursuivre la croissance du tronc (axe principal) et les bourgeons les plus proches donneront des rameaux latéraux. Le même phénomène se reproduisant chaque année, on observera des étages successifs de rameaux latéraux. Ceci est particulièrement remarquable dans le cas des conifères où chaque étage peut constituer une couronne très régulière.
L’orientation de l’axe vertical et des branches latérales dépend, de plus, d’autres facteurs comme le gravitropisme.
Hypotonie et épitonie – le port des branches
Les branches latérales d’un arbre n’ont pas seulement une croissance plus ou moins plagiotrope (plus ou moins horizontale). Elles grandissent selon une règle proche de l’acrotonie : le bourgeon terminal et ses plus proches voisins se développent et permettent une ramification secondaire. Cependant cette ramification est souvent dissymétrique.

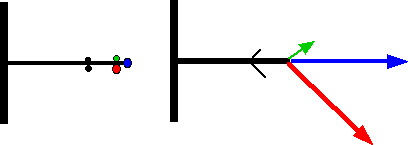
Dans le cas du marronnier et de beaucoup d’autres arbres, le bourgeon secondaire situé au-dessous se développe davantage que le bourgeon situé au-dessus : c’est l’hypotonie.
Le cas inverse (développement maximum des bourgeons situés au-dessus) peut se présenter. L’épitonie est le plus souvent la règle chez les arbustes alors que l’hypotonie est quasiment la règle chez les arbres caducifoliés.
D’autres éléments peuvent influencer le port d’un arbre : prenons comme exemple le peuplier.

On aperçoit deux peupliers, de forme élancée, dans cette photographie d’un bosquet.
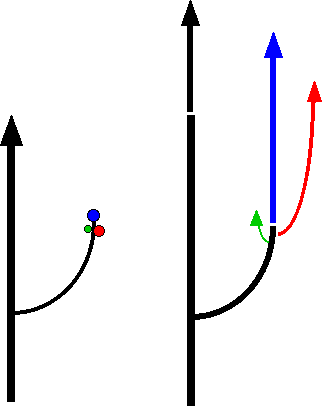
Au niveau de la zone de croissance on observe une courbure vers le haut par gravitropisme négatif.
Dans ce cas, ce n’est pas une épitonie mais un gravitropisme très prononcé qui provoque la croissance verticale des branches, dans le sens opposé à la pesanteur; il s'agit d'un gravitropisme négatif.
Monopode et sympode – exemple de marronniers
Dans le cas de la croissance de type acrotone, un autre système peut
introduire une variation morphologique. Sur l’ensemble des bourgeons
terminaux, certains peuvent exercer une prédominance sur les autres.
Dans le cas le plus général de l’acrotonie chez les arbres, c’est le
bourgeon terminal axial qui se développe prioritairement. Les bourgeons
axillaires latéraux donneront les branches ; c’est la croissance
monopodiale.
Dans certains cas, le bourgeon axial se développe peu ou avorte. C’est
alors un ou plusieurs bourgeons axillaires qui prennent le relai. Lorsqu’un
tel système est régulier, on parle de croissance de type sympodial. Ce type
de croissance est réalisé chez divers arbustes comme le lilas. Ici, pour
comparer, prenons comme exemple deux variétés de marronnier.

A gauche: une plante à croissance monopodiale observée en hiver, le cas typique de l’acrotonie.
A droite: une plante à croissance sympodiale observée en hiver. Le bourgeon terminal a avorté et deux bourgeons axillaires équivalents préparent le relai.
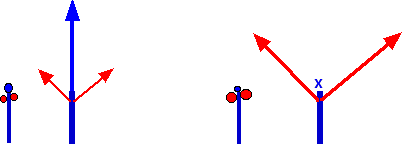
À gauche, le monopode et à droite, le sympode. Dans les deux cas, le premier schéma montre l’importance des bourgeons en hiver et le deuxième schéma, leur développement au printemps suivant.
Ces divers processus, caractéristiques d’une espèce, permettent à celle-ci d’acquérir son port adulte et donc sa silhouette reconnaissable. Les facteurs de l’environnement interagissent avec ces comportements intrinsèques.
Port et environnement
Les facteurs de l’environnement agissent sur le phénotype des plantes. Le facteur le plus important est certainement la gravité. Comme elle s’exerce partout, elle est souvent peu perceptible et difficilement analysable. Les réactions gravitropiques des plantes leur permettent de réguler précisément leur croissance par rapport à la verticalité et ceci correspond à un des éléments les plus essentiels de l’acquisition de leur port. Abordons maintenant quelques autres facteurs facilement observables.
La présence d’autres arbres
Le port d’un arbre d’une même espèce peut varier notablement s’il est isolé, au bord d’une clairière ou au sein d’une futaie. Les facteurs du milieu sont variés (lumière, encombrement spatial, nature du sol, etc.)

Les pins isolés présentent un port élargi au sommet (branches horizontales, développement plagiotrope) par rapport aux pins regroupés.
Le vent continu
Le vent continu peut engendrer un port en drapeau mais attention à l’interprétation de cette morphologie, qui peut avoir plusieurs causes.
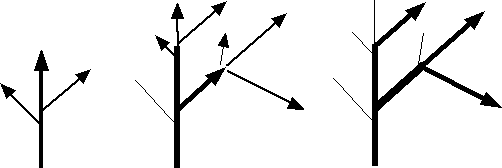
Schéma du développement de ramifications sous l’action d’un vent permanent venant de gauche.
L’anémomorphose (acquisition d’une forme particulière sous l’effet du vent) est un accomodat qui consiste en la modification de la forme des plantes et des paysages végétaux sous l’effet de forts vents dominants. Il constitue un cas particulier de la thigmomorphogenèse (morphogenèse sous l’influence d’un contact, différent de l’effet de la gravité) qui aboutit à des ports « en drapeau », voire « couchés ».
L’action de l’homme
La taille des arbres à des fins économiques ou esthétiques peut modifier considérablement la silhouette d’un arbre. Cependant, après coup, ce sont les caractéristiques de croissance de l’espèce qui reprennent le dessus. Les jardiniers et arboriculteurs se servent de la connaissance de ces caractéristiques pour obtenir des proportions harmonieuses.

Exemple d’arbres taillés par l’homme, de façon drastique et régulière.
Conclusion
Nous avons consacré ce chapitre aux arbres en en choisissant deux parmi les plus connus, le chêne et le pin. Chacun de nous a la possibilité de faire les mêmes observations, et bien d’autres, à partir des arbres disponibles dans son environnement. Les arbres sont intéressants car l’acquisition de leur port, très complexe, dépend en grande partie des règles qui régissent la ramification de leurs branches et il est possible, à partir de leur observation, de tirer ces quelques règles simples.
Naturellement, n’importe quel type de plante peut se prêter à la même
étude. Notons que la morphologie finale d’une plante (plante adulte) dépend
des importances relatives des bourgeons terminaux et axillaires mais
également de la disposition ainsi que de la forme et de la couleur de ses
feuilles.
Les caractéristiques de l’inflorescence, de la fleur et du fruit jouent
également un rôle majeur dans la morphologie générale de la plante et sont
utilisées pour la détermination systématique des plantes. Enfin, des
caractères temporels sont également importants. Une plante atteint sa
maturité de développement parfois à une date précise (date de floraison par
exemple).
La morphogenèse (ou acquisition de la forme = phénotype) dépend donc de très nombreux événements qui se déroulent dans la plante de manière intrinsèque (=génotype). Le déroulement spatio-temporel de ces événements est continuellement réglé par les facteurs de l’environnement. Certains, comme la gravité, s’exercent en permanence et jouent un rôle fondamental, mais peuvent passer inaperçus. Certains se traduisent, comme la lumière ou le contact, par des changements de position et peuvent être provoqués expérimentalement. Certains autres, comme la longueur du jour, peuvent influencer le déroulement morphogène de manière temporelle.
L’expérimentation chez des plantes de grande taille, comme les arbres, est difficile. Aussi, de nombreuses expériences sont réalisées chez de jeunes plantes herbacées (des plantules issues de la germination).
La morphogenèse de jeunes plantules
Les jeunes plantules constituent un matériel expérimental facile à obtenir en un temps court (quelques jours) de manière très reproductible. Pour bien analyser les réponses expérimentales de ces organismes, il est important de bien connaître leur morphologie et les caractéristiques de leur morphogenèse naturelle. Quatre espèces bien caractéristiques sont étudiées ici, deux monocotylédones (le blé et le maïs) et deux dicotylédones (le pois et le soja).

De gauche à droite: on peut observer une plantule de pois, une plantule de blé, une plantule de maïs et une de soja.
Pour chacune de ces plantules, on essaiera de soulever quelques problèmes intéressant l’expérimentateur
Voir aussi sur ce sujet l'article Les plantules modèles : pois, blé, maïs et soja (partie 1 et partie 2).
La germination du pois
Vous trouverez ci-dessous trois étapes de la germination d’une graine de petit pois.
La germination au sens strict débute par une imbibition et se termine par la sortie de la radicule.
La suite (germination au sens large) peut être considérée comme de la croissance.
On observe successivement la croissance de la jeune racine, puis celle de la jeune tige. Celle-ci s’élève au-dessus des cotylédons de la graine : il s’agit d’une tige épicotylée. Le reste de la graine, et en particulier les cotylédons, ne se soulève pas. Les cotylédons restent à la surface ou sous la surface de la terre : il s’agit d’une germination hypogée.

Sur une graine de pois en début de germination on peut clairement observer que les cotylédons sont en relation directe avec l'axe embryonnaire
L'ouverture de la graine, au début de la germination, montre des cotylédons en relation directe avec l’axe embryonnaire. Ils font partie intégrante de l’embryon. Les réserves sont situées, dans ce cas, dans les cotylédons, donc dans l’embryon lui-même. Il n’y a pas d’albumen chez la graine mature : la graine est dite exalbuminée.

Sur cette graine de pois en germination, les cotylédons ont été écartés, laissant apparaître leur relation avec l'axe embryonnaire.

Sur cette jeune racine se développent des racines secondaires.
L’observation du système racinaire d’une plantule plus âgée montre que la radicule est devenue une racine principale qui grandit verticalement vers le bas (gravitropisme positif, voir figure ci-contre).
Le long de cette racine se développent des racines secondaires (ou de deuxième ordre) qui grandissent horizontalement ou selon un certain angle avec la verticale.
On verra que ce n’est pas toujours le cas (monocotylédones). Ces racines apparaissent à une distance assez précise et constante de l’extrémité puis grandissent au fur et à mesure que la racine principale grandit.
La germination du blé
Le grain de blé est un caryopse, c’est-à-dire un akène (fruit sec indéhiscent contenant une graine) particulier, caractéristique des graminées. Son enveloppe protectrice est constituée de deux parties, le péricarpe du fruit et le tégument de la graine. Ces deux parties sont plus ou moins soudées. Cependant, la graine constituant la majeure partie du caryopse, il est possible d’expérimenter sur cet organe comme si c’était une graine.

Le "grain" de blé est en réalité un fruit dont le péricarpe est soudé au tégument de la graine (caryopse). On peut étudier sa germination comme s'il s'agissait d'une graine.
Sur les trois étapes de la germination d’un grain de blé, ci-dessus, on observe la sortie d’une première racine et d’un organe aérien (en vert), le coléoptile. Très rapidement le coléoptile grandit verticalement et de nouvelles racines apparaissent. Ces nouvelles racines sont ici au nombre de trois.
Des agrandissements (ci-dessous) permettent de mieux comprendre la formation de ces racines. La première racine était protégée par un étui, le coléorhize, qu’elle a rapidement percé. Les deux autres racines ne sont pas des racines secondaires apparues sur la racine principale, mais des racines adventives initiées dans le corps de l’axe embryonnaire.


Le coléoptile est un étui qui protège l’apex caulinaire et les jeunes feuilles. À partir d’un certain stade, sa croissance résulte essentiellement d’une croissance par élongation des cellules (auxèse). C’est cette propriété qui en fait un matériel de choix pour l’expérimentation sur la croissance cellulaire (de Darwin à Went et Thimann). Lorsque sa croissance est terminée, les jeunes feuilles le percent au niveau d’une ouverture préexistante.
La germination du maïs
Le grain de maïs (graminées) est un caryopse, comme le grain de blé.
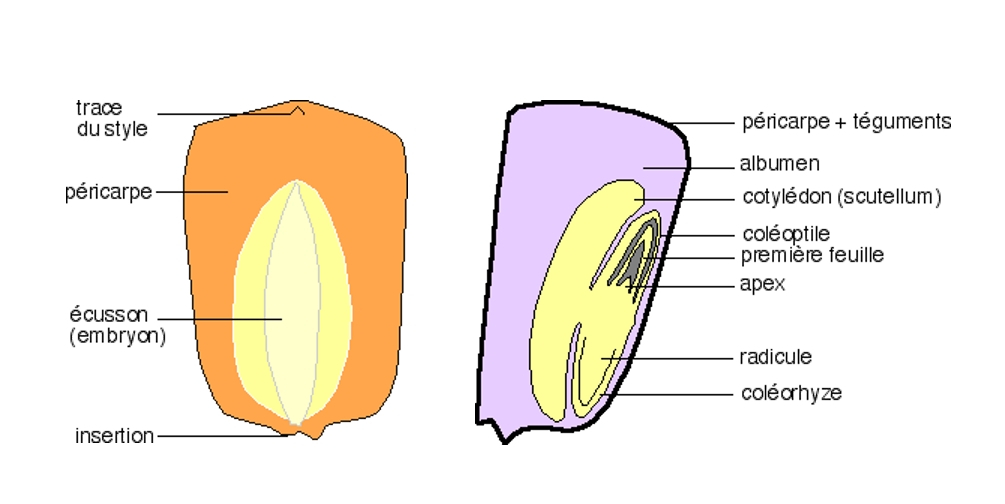
Le schéma de gauche présente l'aspect externe du grain du maïs et celui de droite une coupe sagittale longitudinale montrant l'organisation générale de ce caryopse

Sur ce caryopse de maïs, ouvert en deux longitudinalement, une moitié a été colorée au lugol
Une coupe longitudinale sagittale montre l’organisation générale d’un caryopse. Une coloration par le lugol montre que les réserves amylacées sont localisées dans un tissu spécial, indépendant de l’embryon, l’albumen. La graine de ce caryopse est dite albuminée.
Sa germination suit les mêmes règles. Comme il est plus gros, il se prête mieux à certaines expérimentations. Sa morphogenèse présente quelques particularités.


La germination à la lumière ressemble à celle du grain de blé. La germination à l’obscurité fait apparaître une particularité ; à savoir l’existence d’un organe tubulaire entre le corps de l’embryon et l’insertion du coléoptile : le mésocotyle.
Sur les agrandissement ci-dessous, on constate que de nouvelles racines adventives apparaissent de manière assez irrégulière à partir de l’axe embryonnaire, mais non à partir de la racine, comme dans le cas du pois.
Chez le maïs, il n’y a donc pas un système racinaire pivotant constitué d’une racine principale et de racines de deuxième ordre, mais un système racinaire fasciculé, formé de racines adventives de même ordre.

À gauche, on observe très nettement le coléorhize percé par la radicule.
À droite, le mésocotyle, qui soulève le coléoptile (et donc l’apex et les jeunes feuilles contenues dans le coléoptile), est bien délimité.
Des sections longitudinales sagittales à deux stades de la germination permettent de bien montrer les relations entre les différentes parties.

Sur ces deux sections, on visualise l'albumen en blanc, le scutellum (cotylédon unique) translucide, la plantule (sur le côté gauche). Sur l'image de droite on distingue deux vaisseaux grisâtres dans le mésocotyle de la plantule.
La germination du soja
Le soja vert, utilisé dans l’alimentation sous forme de graines ou de germinations étiolées, représente un excellent matériel expérimental pour l’étude de la croissance cellulaire.
En effet, comme les coléoptiles, à partir d’un certain stade, la croissance de son hypocotyle est principalement due à de l’auxèse.
Comme on le voit sur la série d’images représentant des plantules d’âges différents, les cotylédons sont soulevés au-dessus du sol (germination épigée) grâce à la croissance d’un organe intermédiaire entre la racine et l’apex, l’hypocotyle.

Les deux images suivantes montrent deux états des plantules de soja dans une barquette de germination, 12 heures séparent les deux photographies.

La culture de soja de droite est plus âgée de 12 heures que celle de gauche
Il est relativement aisé avec ce matériel d’établir une courbe de croissance de manière qualitative et quantitative. Toutes les germinations épigées de type « haricot » se réalisent selon le même processus.
Comme chez le pois, les réserves sont contenues dans les cotylédons qui appartiennent à l’embryon. Ces graines sont donc exalbuminées. Chez le soja, une grande partie des réserves est de nature protéique. Chez le haricot, les réserves sont en majorité amylacées. En ouvrant une graine de haricot entre les deux cotylédons, puis en la colorant par le lugol, on met bien en évidence son anatomie et la localisation de ses réserves.

À gauche, les deux cotylédons de la graine de haricot sont écartés, sur l’un d’eux est attachée la jeune plantule.
À droite, cette même structure a subi une coloration au lugol, mettant en évidence des réserves amylacées dans les cotylédons (mais pas dans la plantule).
Action de la lumière et étiolement
La plupart des expériences concernant la croissance sont réalisées sur des plantules étiolées, c’est-à-dire dans des conditions peu naturelles. Pourquoi ?
La lumière inhibe sensiblement la croissance. À l’obscurité, la croissance par auxèse des coléoptiles, épicotyles ou hypocotyles est plus forte et les réponses hormonales (à l’auxine en particulier) plus nettes. On peut ainsi se poser la question de l’environnement lumineux sur la morphogenèse d’une plantule.
Observons deux exemples, le pois (dicotylédones) et le maïs (monocotylédones)

À gauche : germination de pois cultivé à la lumière.
À droite : germination de pois cultivé à l’obscurité.

À gauche : germination de maïs cultivé à la lumière.
À droite : germination de maïs cultivé à l’obscurité.
Le pois cultivé à l’obscurité présente les symptômes suivants, comme la plupart des dicotylédones :
-
croissance en longueur des entre-nœuds beaucoup plus forte qu’à la lumière
-
croissance des feuilles plus faible
-
absence de chlorophylle (la tige est blanche et les feuilles jaunes).
Le maïs réagit de manière légèrement différente, comme beaucoup de monocotylédones.
-
croissance en longueur du coléoptile plus forte qu’à la lumière
-
croissance des feuilles peu modifiée
-
absence de chlorophylle (le coléoptile est blanc et les feuilles jaunes).
Les modifications touchent donc :
-
la croissance en longueur des tiges
-
la différenciation de l’appareil photosynthétique (développement des feuilles, différenciation des chloroplastes, synthèse de chlorophylle).
Quelle peut être la signification biologique de la croissance exacerbée observée à l’obscurité ? Réalisons une expérience très simple : dans un pot recouvert de papier aluminium, plaçons des grains de maïs à différentes profondeurs. Lorsque les jeunes feuilles sortent à la surface, déterrons les germinations.

A gauche le pot contenant les grains de maïs qui ont été placés à différentes profondeurs, à droite les germinations déterrées. On constate que la hauteur des parties aériennes des différentes germinations est la même, quelle que soit la profondeur à laquelle les grains avaient été enterrés.
On constate que, quelle que soit la profondeur du semis, les parties aériennes sont toutes au même niveau. Ceci est dû à la croissance du mésocotyle, très rapide à l’obscurité (en profondeur) et qui se ralentit lorsque les parties aériennes arrivent en surface.
Cette expérience montre que le facteur externe « lumière » (présence ou absence) permet à la plante de régler sa morphogenèse en s’adaptant aux conditions locales et momentanées.
Conclusion
Germinations et jeunes plantules constituent un matériel de choix pour l’expérimentation sur la croissance. Chaque type de germination possède ses règles propres de morphogenèse, dues à son génotype. Cependant le phénotype de la plantule, c’est-à-dire son aspect et sa physiologie (et aussi sa constitution biochimique) dépend des facteurs de l’environnement qui ont pu agir sur son développement.
Le facteur le plus aisé à mettre en évidence est la présence ou l’absence de lumière (étiolement). La lumière agit dans ce cas par son intensité, et plus spécifiquement par sa longueur d’onde comme « signal morphogène ».
Dans certains cas, la lumière peut également agir par sa direction. Elle produit alors des réactions de croissance différentielle qui modifient la position de certains organes. La réaction de la plante à cet éclairement directionnel est appelé le phototropisme.
D’une manière plus générale, les tropismes sont des réactions de courbure
dues à une croissance différentielle provoquée par un facteur dissymétrique
de l’environnement. On peut étudier assez aisément les réactions provoquées
par la lumière (le phototropisme), par la gravité (le gravitropisme) et par
un contact (le thigmotropisme). Ces réactions, mises en évidence
expérimentalement lorsque l’on ne fait varier qu’un seul facteur,
s’intègrent dans les réactions naturelles à l’ensemble des facteurs
internes et externes qui règlent la morphogenèse.
Voir l’article « La morphogenèse végétale : action dirigée des facteurs de
l’environnement ».
Localisation de la croissance, l’expérience de Sachs
La croissance cellulaire n’est pas régulière dans un organe ; il existe des zones de croissance. Celles-ci ne sont pas délimitées précisément et il vaut mieux parler de « gradients de croissance ». L’expérience classique de Sachs a permis de préciser la zone de croissance dans les racines.
La technique consiste à réaliser des marques (encre de Chine par exemple) équidistantes sur une jeune racine, à partir de sa pointe. Après 24 heures, la racine a grandi et les marques se sont déplacées. Les intervalles situés entre deux marques qui ont grandit le plus se situent entre le 2 e et le 4e millimètre au-dessus de l’extrémité. On en déduit que le maximum de croissance s’est réalisé à ce niveau dans la région subterminale.
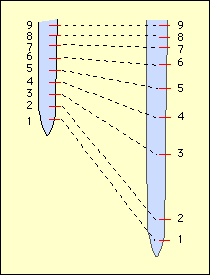
Remarque : la compréhension spatio-temporelle de ce phénomène est en fait très complexe, car la croissance n’est pas linéaire pendant les 24 heures de l’expérience. En effet, si l’intervalle (2-3) en pleine croissance au temps 0 se trouvait bien dans la région 2-3 mm, il se trouve, après 24 heures, dans une région située entre 3 et 7 mm…qui ne grandit plus !
Ce type d’expérience est souvent difficile à réaliser, car les racines sont fragiles et doivent être manipulées en conditions d’humidité importante.
Comment concilier cette observation avec l’étude histologique de la racine ?

La zone de croissance observée par Sachs correspond principalement à la zone de division et à la zone d’élongation, l’ensemble de ces zones se trouvant bien en position subterminale.
Ce type d’expérience peut être appliqué à différents organes comme des hypocotyles. Les images suivantes montrent quatre étapes de la croissance d’un hypocotyle de soja marqué. Une séquence vidéo accélérée montre la cinétique de la croissance.
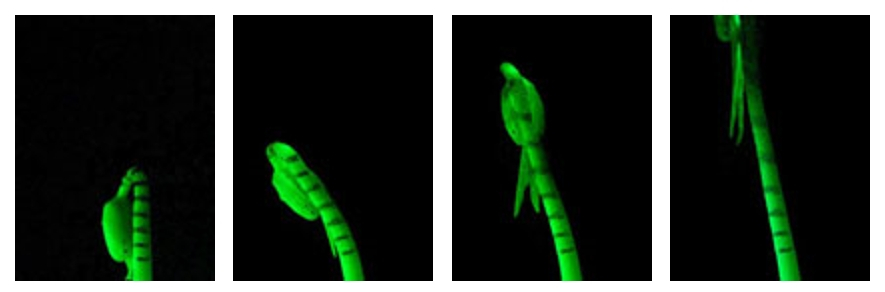
Début de l'observation, à gauche, et fin, à droite. On note l'écartement progressif, mais de façon non linéaire, des marques faites sur l'hypocotyle.
Vidéo accélérée de la croissance d'un hypocotyle de soja
Si dans la racine les zones de mérèse et d’auxèse sont distribuées séquentiellement dans le sens longitudinal et donc assez facilement repérables, il n’en est pas de même dans les méristèmes de la tige. Dans ce cas, la distribution est complexe, d’autant plus que le fonctionnement du méristème contribue à la fois à la croissance en longueur de la tige mais aussi à la formation des jeunes feuilles.