Cet article explique par quel processus l'inactivation de l'un de deux chromosomes X est réalisée et transmise.
Introduction
Chez la plupart des Mammifères, les chromosomes sexuels des mâles, XY, sont très peu homologues contrairement à ceux des femelles, XX (Figure 1).
L’absence d’un second X chez les mâles les désavantage-ils vis-à-vis des femelles dans le cas où leur seul X serait déficient pour un ou plusieurs gènes ? Non, car les cellules des femelles fonctionnent aussi avec un X unique, l’autre X étant inactivé.
Quelles sont les signatures de cette inactivation et quel en est le mécanisme ?
Notons d’emblée que l’inactivation du chromosome X a été, jusqu’ici, bien plus étudiée chez les Mammifères placentaires (Euthériens) que chez les Mammifères marsupiaux (Métathériens) ou les Mammifères pondeurs d’œufs (Protothériens) [1].
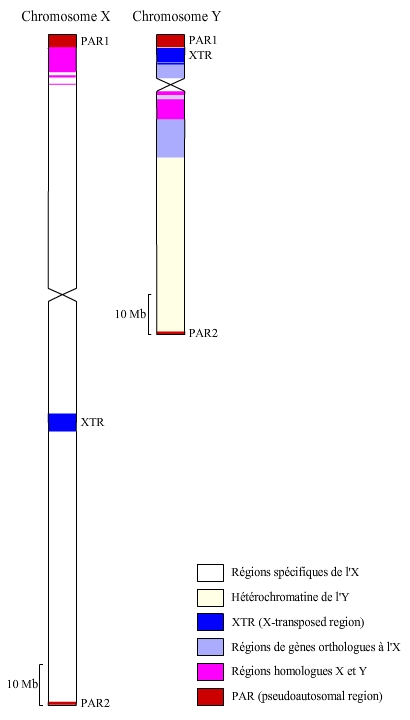
La comparaison des séquences du chromosome X et du chromosome Y permet de reconstituer l'histoire évolutive de ces deux chromosomes depuis la paire d'autosomes ancestrale.
Les signatures de l’inactivation
Une signature cytologique, le corpuscule de Barr
Entre deux divisions cellulaires et en dehors du ou des nucléoles, dont la chromatine est suffisamment condensée, la chromatine est trop diffuse pour être visible au microscope optique. À l’exception aussi, chez les femelles de Mammifères placentaires et marsupiaux, d’un autre corpuscule, généralement localisé en bordure de la membrane nucléaire, qui est appelé corpuscule de Barr (cb), du nom du chercheur canadien qui le mit en évidence en 1949.
Ce corpuscule n’existe pas chez les Mammifères pondeurs d’œufs dont les chromosomes X et Y, par ailleurs quintuplés, sont de taille presque égale (le chromosome Y a raccourci au cours de l’évolution des Mammifères, voir article « Le chromosome Y humain, portaits croisés »).
Les corpuscules de Barr ont été dénombrés dans des cellules au caryotype atypique car contenant un nombre variable de X, ce qui donne les relations suivantes dans l’espèce humaine (voir Fig. 2) :
- Individus A : 46 chromosomes dont XY, 0 corpuscule de Barr ;
- Individus B : 46 chromosomes dont XX, 1 corpuscule de Barr ;
- Individus C : 47 chromosomes dont XXX, 2 corpuscules de Barr ;
- Individus D : 48 chromosomes dont XXXX, 3 corpuscules de Barr ;
- Individus E : 49 chromosomes dont XXXXX, 4 corpuscules de Barr.
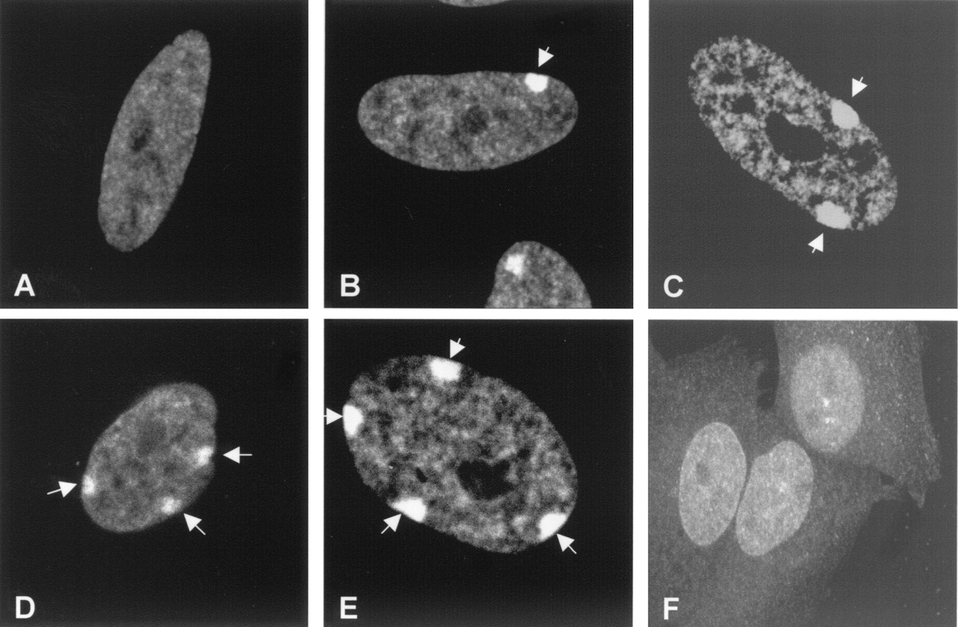
Les cellules utilisées sont des fibroblastes humains diploïdes qui se différencient par le nombre de leurs chromosomes X (respectivement 1, 2, 3, 4 et 5 pour les photos A, B, C, D et E).
La photo F correspond à une photo d'une lignée cellulaire hybride humain-hamster contenant un chromosome X humain inactivé.
Les flèches pointent sur les corpuscules de Barr.
Partant du fait que :
- d’une part, il n’y a pas de corpuscule de Barr chez les cellules mâles XY, dont on sait par ailleurs que l’X est activé ;
- d’autre part, chez les cellules femelles il y a systématiquement un corpuscule de Barr de moins que de X totaux,
ce double constat incite fortement à penser que corpuscules de Barr et inactivité des X sont liés, c’est du moins l’hypothèse la plus économe.
Une signature fonctionnelle, l’exemple des chattes bicolores
De nombreux gènes gouvernent la couleur du pelage. Chez le chat, l’un d’entre eux localisé sur le chromosome X existe sous deux variantes (ou allèles), l’une codant pour la couleur noire, l’autre pour la couleur orangée. Les mâles porteurs de ce gène sont toujours uniformément colorés, tout noirs ou tout orangés : leur pelage exprime l’allèle du seul X dont ils ont hérité, qui ne peut être que celui de la mère (Xm).
Les femelles hétérozygotes pour ce gène, c’est-à-dire qui ont reçu un allèle de chaque type – l’un d’origine maternelle (Xm), l’autre d’origine paternelle (Xp) – ne sont pas uniformément colorées noires, oranges ou orange foncé comme on pourrait s’y attendre en se basant sur la dominance éventuelle d’un allèle sur l’autre, ou leur codominance. Ces chattes, dites calico, sont bicolores, tachetées de noir et d’orange ; leur pelage mosaïque exprime selon les territoires, tantôt l’allèle porté par un X, tantôt l’allèle porté par l’autre X, et ce de façon permanente puisque ni la forme ni la couleur des taches ne varient au cours de leur vie.
Cette inactivation ne concerne pas un seul gène mais la presque totalité des gènes de l’X. En effet, il a été montré qu’au début de la vie embryonnaire des femelles placentaires, un X sur deux (Xp ou Xm au hasard) est presque entièrement inactivé dans chacune des cellules de leur masse cellulaire interne (à l’origine de tous les tissus de l’individu, voir Fig. 3 et 4).
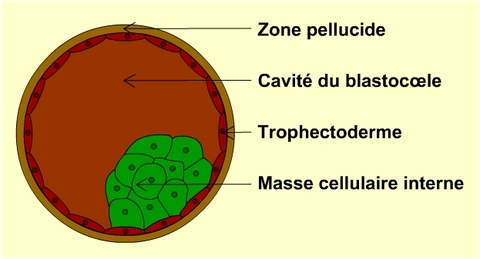
Cette inactivation au hasard de l’X concerne donc aussi bien les parties externes que les parties internes du corps : cerveau, foie, cœur, poumons…
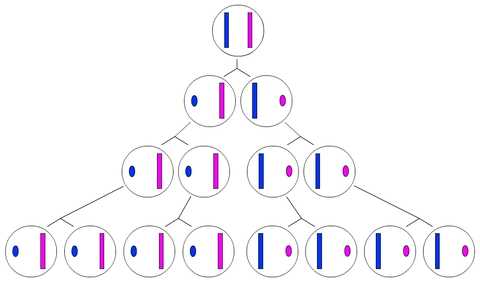
Très précocement au cours du développement, l'un des deux chromosome X est inactivé en donnant naissance au corpuscule de Barr.
Cette inactivation est alors transmise à la descendance de la cellule.
Le choix de l'inactivation du chromosome X paternel (en bleu) ou maternel (en rose), est aléatoire.
C’est la britannique Mary Lyon qui en 1961, mettant en relation pelage mosaïque des souris et corpuscules de Barr, émit pour la première fois l’hypothèse de l’inactivation du chromosome X.
Quelques aspects du mécanisme de l’inactivation
Ce mécanisme d’inactivation est encore loin d’être élucidé, mais quelques aspects de sa mise en place et de sa maintenance peuvent être indiqués [3,4].
Sa mise en place
L’intervention d’un centre d’inactivation de l’X (Xic), long d’un million de bases (1Mb), fut mise en évidence, d’abord chez la souris dans les années 70-80, puis chez d’autres placentaires :
- des expériences supprimant ce centre (en quelque sorte supprimant ce suppresseur) rendaient l’X actif,
- des expériences de translocation de ce centre, sur un autosome, avaient pour effet, à l’inverse, de rendre cet autosome partiellement inactif.
L’identification, en 1991, de l’un des gènes dont ce centre est porteur, le gène Xist (effecteur majeur de l’inactivation), fit avancer de façon décisive la compréhension du mécanisme de l’inactivation. On a pu établir que le gène Xist s’exprime, soit sur l’Xp, soit sur l’Xm, de façon apparemment aléatoire, dans les premiers moments de la vie embryonnaire, au sein des cellules de la masse cellulaire interne de l’embryon. Il gouverne la synthèse d’un ARN Xist de 19 kilobases (X inhibitory specific transcript). Ce transcrit est non codant, c’est-à-dire non traduit en protéine. Produit en grande quantité, il finit par tapisser entièrement, et uniquement, le chromosome Xp ou Xm incomparablement plus grand (160 millions de bases) dont il est lui-même issu (action en cis), le rendant ainsi presque entièrement inactif, et produisant donc le corpuscule de Barr (Figure 5). Cet ARN Xist agit en interdisant l’accès de la machinerie transcriptionnelle au chromosome X qu’il recouvre. Il apparaît également impliqué dans le transfert du corpuscule de Barr au contact de la membrane nucléaire [4].
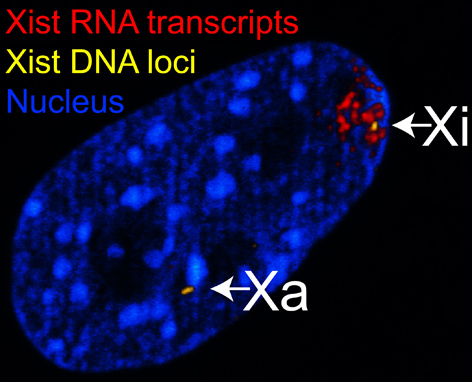
Noyau d’un fibroblaste de souris femelle observé en microscopie confocale. L’ADN est révélé par la fluorescence bleue du DAPI. Le locus du gène Xist est mis en évidence par hybridation fluorescente in situ (FISH, en jaune). Ce locus est présent en deux exemplaires correspondant au chromosome X actif, Xa, et au chromosome X inactif, Xi. Le transcrit Xist, en rouge, n’est présent qu’au niveau du chromosome X inactif, Xi.
Le chromosome compagnon, Xm ou Xp, n’exprime pas cet ARN Xist inhibiteur et devient, par là-même, l’X actif de la cellule.
On a pu montrer qu’en amont de ce processus, c’est à dire de l’expression monoallélique de Xist, se trouve un facteur autosomal, présent en quantité limitante, qui, au hasard, soit sur le Xp soit sur le Xm, méthyle le promoteur du gène Xist et ce faisant l’éteint.
Chez les placentaires, l’inactivation de l’X ne se limite pas aux seules cellules de la masse cellulaire interne de l’embryon. Elle se produit aussi dans les cellules dérivées du trophectoderme (voir Fig. 3) génératrices des annexes embryonnaires, soit qu’elle ait lieu au hasard sur l’Xm ou l’Xp, c’est le cas de l’espèce humaine, soit qu’elle se porte spécifiquement sur l’Xp comme chez les souris.
L’inactivation de l’X emprunte donc des chemins différents chez les placentaires. Et c’est encore plus vrai des marsupiaux dont l’X inactivé est toujours le Xp, dans la masse cellulaire interne comme dans le trophectoderme, et chez lesquels l’homologue du gène Xist, appelé Ln 3, n’a rien à voir avec l’inactivation de l’X puisqu’il code en fait pour une protéine participant à des voies de signalisation intracellulaire. Ce sont donc un ou d’autres gènes qui gouvernent l’inactivation de l’X chez les femelles de Marsupiaux, gène(s) qu’il reste à identifier.
Quant aux Mammifères pondeurs d’œufs, sans corpuscules de Barr et aux chromosomes sexuels X et Y quintuplés et presque de même taille, l’inactivation de leurs X est restreinte aux portions de l’X qui n’ont pas leur équivalent sur l’Y.
La mise en place de l’inactivation de l’X chez les femelles de Mammifères a donc connu des rebondissements. Pour les pister, il reste à progresser dans la biologie cellulaire et moléculaire des marsupiaux et des Mammifères pondeurs d’œufs, ainsi que dans la séquence de leur génome [5]…
Sa maintenance
Le gène Xist n’intervient qu’un temps dans l’inactivation de l’X. Rapidement, son expression cesse sur le chromosome inactivé n’alimentant plus la production d’ARN recouvreur. Son action est alors relayée par les modifications épigénétiques qui se produisent sur l’X inactivé, c’est-à-dire par des modifications qui ne touchent pas à la séquence de l’ADN, mais qui ajoutent un groupement méthyle à certaines de ses bases (cytosines) ou des groupements méthyle/acétyle, aux histones auxquelles il est lié.
On a vu que c’est une telle méthylation, pour être plus précis de cytosine, déclenchée par un facteur autosomal, qui inhibe le promoteur du gène Xist de l’X, levant l’inactivation de ce dernier.
De même, ce sont de telles modifications d’histones réalisées spécifiquement sur le Xp du trophectoderme de souris (on parle alors d’empreinte génétique paternelle) qui sont à l’origine de l’inactivation du Xp dans ce tissu murin.
Ces modifications épigénétiques, dont on découvre de plus en plus qu’elles jouent un rôle déterminant dans la façon dont est modulée l’expression des gènes, sont transmises au fil des divisions cellulaires et concernent également les chromosomes autosomaux. Elles s’additionnent au cours de la vie d’un individu ce qui explique en partie, par exemple, que de vrais jumeaux finissent par moins se ressembler au fur et à mesure qu’ils vieillissent. Dans un autre registre, incorrectement apposées, elles donnent naissance à des maladies.
Conclusion
L’inactivation de l’X des femelles de Mammifères s’est mise en place à plusieurs reprises au cours de l’évolution.
Celle des cellules du corps des placentaires (dérivées de leur masse cellulaire interne) se manifeste par l’existence d’un corpuscule de Barr et par l’expression mosaïque de la quasi-totalité des gènes lorsqu’ils sont présents à l’état hétérozygote sur les chromosomes X paternel et maternel.
Dans ce processus d’inactivation/activation de l’X, des facteurs à la fois directement génétiques (enrobage par l’ARN Xist) et épigénétiques (inhibant le processus en amont, ou venant au contraire le relayer en aval) sont à l’œuvre.
Ainsi, mâles et femelles disposent d’un seul X actif. Il est à noter que cet unique X fonctionnel s’exprime en quantité deux fois plus importante que les chromosomes autosomaux, ce qui rétablit la balance avec eux (voir Fig.7 de la référence 3).
Cette inactivation de l’X et son expression mosaïque, Xp ou Xm, rend compte de la moins grande sévérité de certains traits chez les femelles hétérozygotes que chez les mâles porteurs de la version délétère d’un gène : en effet elles possèdent toujours dans leurs tissus une fraction de cellules exprimant la version fonctionnelle du gène.
Mais il faut bien garder à l’esprit qu’un gène n’agit généralement pas seul, et qu’il est influencé aussi bien par son environnement que par les autres gènes du génome. Tout n’est pas aussi simple qu’avec le pelage des chattes calico !
Bibliographie
-
Le chromosome X maître du silence. (2005) La Recherche 385 (avril).
-
Bo Hong, Peter Reeves, Barbara Panning, Maurice S. Swanson et Thomas P. Yang. Identification of an autoimmune serum containing antibodies against the Barr body (2001) Proc. Natl. Acad. Sci. USA 98, 8703-8708.
-
Edith Heard et Christine M. Disteche. Dosage compensation in mammals : fine-tuning the expression of the X chromosome (2006) Genes & Development 20:1848-1867.
-
Julie Chaumeil, Patricia Le Baccon, Anton Wutz et Edith Heard. A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. (2006) ref 5 06/08pubheard Xist RNA role exclus° of RNA pol and transc factors Genes & Development 20:2223-2237.
-
Loïc Mangin. Sexe à rebondissements. (2006) Pour la Science. 346 (aout) : p24.