En génétique, l'épistasie désigne l'interaction existant entre deux ou plusieurs gènes, l'un d'entre eux (ou plusieurs) masquant ou empêchant l'expression des autres. Cet article fait le point, en utilisant différents exemples réels et cas de figures théoriques, sur les proportions des divers phénotypes observables dans les descendances, selon le nombre de gènes impliqués et le type d'épistasie ; dominante ou récessive.
Introduction
En génétique mendélienne classique, on commence par décrire le cas dans lequel un caractère est contrôlé par un gène unique. On peut prendre comme exemple le canal chlore de type CFTR (Cystic Fibrosis Transmembrane Conductance Regulator) qui est codé par un gène unique (le gène CFTR), localisé en 7q31 (bras long du chromosome 7 humain), et dont certaines mutations provoquent la mucoviscidose ; la maladie génétique autosomique récessive grave la plus fréquente dans la population européenne (environ 1 enfant touché pour 2500 naissances). Cette situation a l’avantage d’être simple mais ne correspond pas à la majorité des cas : le plus souvent, plusieurs gènes interviennent dans le contrôle d’un caractère. L’épistasie correspond à un niveau de complexité supplémentaire puisqu’il s’agit d’une situation dans laquelle il existe une interaction entre l’expression de deux gènes (ou plus), qui contrôlent un unique caractère.
Monohybridisme et dihybridisme
Dans le cas classique du monohybridisme, on étudie les résultats de croisements concernant l’expression d’un seul gène. Pour reprendre l’exemple du paragraphe précédent, il existe un seul gène codant pour le canal CFTR. Chaque individu peut donc être soit homozygote (deux allèles identiques) soit hétérozygote (deux allèles différents). L’étude de la transmission d’un tel caractère permet de mettre facilement en évidence les notions de dominance, récessivité et codominance des allèles existants.
La dominance entraîne un phénotype identique à celui de l’un des parents pour tous les individus de F1. La codominance entraîne un phénotype différent de celui des deux parents pour tous les individus de F1. La récessivité entraîne la réapparition du phénotype de l’autre parent en F2 (dans un rapport de 1/4).
Dans le cas du dihybridisme on étudie les résultats de croisements concernant l’expression de deux gènes. Pour illustration, prenons l’exemple historique des petits pois étudiés par Mendel. La surface de la graine peut être lisse (dominant) ou ridée, et le cotylédon peut être jaune (dominant) ou vert. Chacun de ces deux caractères est contrôlé par un gène. Si on croise un plant de race pure lisse-jaune avec un plant de race pure ridé-vert, on obtient une F1 homogène lisse-jaune (effet de la dominance). Si on croise des plants F1 entre eux pour obtenir une F2, on obtient 4 phénotypes différents dans les proportions de 9-3-3-1, correspondant à 9 génotypes différents (voir Fig. 1).
| J L | J l | j L | j l | |
| J L | JJ LL [Jaune Lisse] |
JJ Ll [Jaune Lisse] |
Jj LL [Jaune Lisse] |
Jj Ll [Jaune Lisse] |
| J l | JJ Ll [Jaune Lisse] |
JJ ll [Jaune ridé] |
Jj Ll [Jaune Lisse] |
Jj ll [Jaune ridé] |
| j L | Jj LL [Jaune Lisse] |
Jj Ll [Jaune ridé] |
jj LL [vert Lisse] |
jj Ll [vert Lisse] |
| j l | Jj Ll [Jaune Lisse] |
Jj ll [Jaune ridé] |
jj Ll [vert Lisse] |
jj ll [vert ridé] |
| J correspond à l'allèle dominant du gène contrôlant la couleur du grain. j correspond à l'allèle récessif du gène contrôlant la couleur du grain. L correspond à l'allèle dominant du gène contrôlant l'aspect du grain. l correspond à l'allèle récessif du gène contrôlant l'aspect du grain. Les phénotypes sont donnés entre crochets []. |
||||
Plusieurs phénomènes peuvent entraîner des variations dans les résultats statistiques cités.
Sans viser à l’exhaustivité, citons parmi les plus classiques :
- le cas d’un allèle létal,
- le cas d’un gène porté par un chromosome sexuel (hérédité liée au sexe),
- ou encore, pour le diybridisme, la proximité physique des deux gènes sur un même chromosome (liaison génétique).
Différents types d’épistasie
Dans le cas d’une hérédité faisant intervenir l’épistasie, les croisements donnent des résultats qui ne peuvent pas être interprétés par le monohybridisme ou le dihybridisme « classique ».
Toujours pour prendre un exemple historique, Emerson, un généticien américain, avait croisé des plants de maïs de race pure à graines jaunes, pourtant la F1 obtenue était 100 % pourpre du fait de la présence d’un pigment, l’anthocyane. La F2 était tout aussi inattendue avec un rapport de 9/16e de pourpre et de 7/16e de jaune.
Le monohybridisme ne permettant pas d’expliquer ces résultats, il fallut envisager que le caractère était contrôlé par deux gènes, mais, plus encore, que ceux-ci interagissaient l’un avec l’autre.
A partir du moment où il y a interaction entre deux gènes (ou plus) on parle d’épistasie.
Plusieurs possibilités d’interaction existent.
L’exemple de la couleur des grains de maïs : effet cumulatif de deux gènes
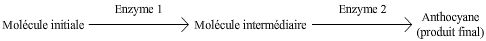
Le pigment pourpre est le produit d’une chaîne de biosynthèse faisant intervenir deux enzymes, chacune codée par un gène différent (voir Fig. 2). Le produit final est coloré mais les molécules en amont sont toutes incolores. Si l’une ou l’autre des enzymes est non-fonctionnelle, le résultat phénotypique est le même, à savoir une absence d’anthocyane, donc de pigmentation, et ceci même si l’autre enzyme est présente et fonctionnelle. L’expression de chacun de ces gènes est donc dépendante de celle de l’autre gène : on parle d’effet cumulatif de deux gènes.
L’exemple des groupes sanguins ABO : épistasie récessive
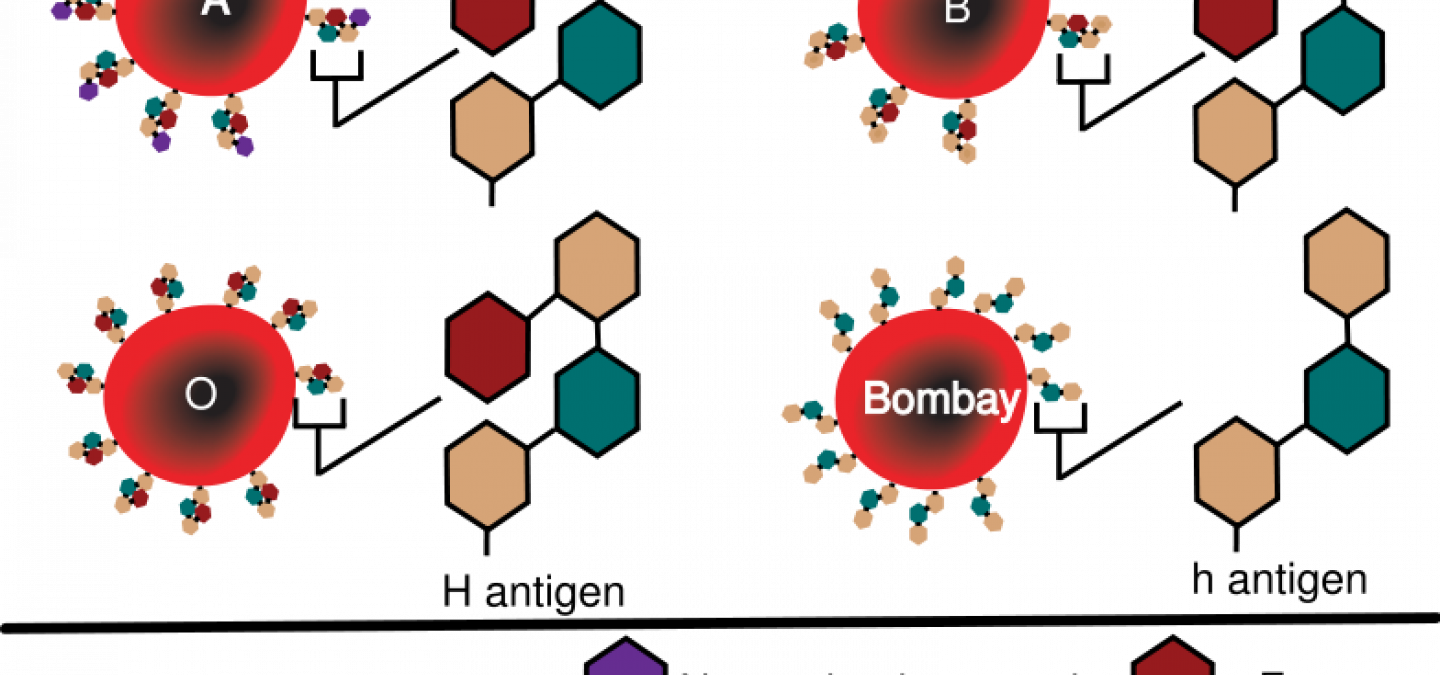
Les groupes sanguins sont un autre cas de figure (pour des détails sur les groupes sanguins, voir l’article Groupes sanguins et conséquences médicales). La voie de biosynthèse se fait en deux étapes, chacune d’elle impliquant une enzyme codée par un gène spécifique (voir Fig. 3), le gène H (deux allèles H et h) et le gène I (trois allèles ; IA, IB et IO). Si le génotype du gène H est HH ou Hh, alors le groupe sanguin dépendra du génotype du gène I : groupe O pour le génotype oo, groupe A pour les génotypes AA ou Ao, groupe B pour les génotypes BB ou Bo, et groupe AB pour le génotype AB. Cependant, si le génotype du gène H est hh, il n’y a pas de substance H à la surface des globules rouges donc il n’y a pas de substrat pour l’action des enzymes codées par le second gène. Quel que soit le génotype de ce gène, l’individu apparaîtra de groupe O .
Dans ce cas, et contrairement au cas de la couleur du maïs, les interactions entre les deux gènes ne sont pas équivalentes : le gène H influence l’expression du gène I mais pas l’inverse. Le gène H est dit épistatique, le gène I hypostatique.
Par ailleurs, c’est l’allèle récessif du gène épistatique qui empêche l’expression du gène hypostatique, puisque si un allèle dominant du gène H est présent, il y a présence de substance H donc possibilité d’expression du gène I. C’est pourquoi on parle d’épistasie récessive.

Le système ABO correspond à la présence, sur la membrane des hématies, de polysaccharides spécifiques. Ces polysaccharides sont composés d'un résidu, qui n'est pas antigénique, et de quelques sucres terminaux qui diffèrent selon les antigènes. Ces sucres sont associées par l'action successive des produits de deux gènes (H puis I). Les allèles de ces gènes, présents chez un individu, déterminent les types de réaction qui peuvent avoir lieu et donc le ou les types d'antigène présents sur ses hématies.
Remarque : Le groupe O étant défini comme une absence d’antigène A et d’antigène B, un individu de génotype HH oo ou Hh oo et un individu de génotype hh apparaissent tout deux comme appartenant au groupe O. Il y a pourtant une différence. Dans le premier cas, le globule rouge porte à sa surface la substance H, dans l’autre non : on parle de phénotype Bombay.
La couleur des pétales : épistasie dominante
Un autre exemple nous est donné avec la couleur des pétales d’une fleur. Ce caractère est sous la dépendance de deux gènes, l’un codant une enzyme permettant la fabrication du pigment (qu’on appellera le gène A) en transformant un précurseur rose pâle en produit rose foncé, l’autre contrôlant le dépôt du piment dans les pétales (qu’on appellera le gène B). Le dépôt ne peut se faire que si les deux allèles portés par le gène B sont récessifs bb. Dans ce cas, en présence des deux allèles aa (pas de transformation du précurseur) les pétales seront roses pâles, et en présence d’au moins un allèle A (transformation du précurseur), les pétales seront roses foncées. Par contre, en présence d’une ou deux copie(s) de l’allèle dominant B, il n’y a pas de dépôt, de sorte que les pétales restent blancs quels que soient les allèles portés par le gène A.
Le gène hypostatique (le A) ne peut donc s’exprimer en présence de l’allèle dominant du gène épistatique (le B) : on parle d’épistasie dominante.
Remarque : Lorsque l’on parle « d’épistasie récessive » ou « d’épistasie dominante », il faut faire attention à ne pas se tromper dans les relations de dominance et de récessivité existantes.
Le nom « épistasie » implique la dominance ou la récessivité d’un gène par rapport à un autre, et non d’un allèle par rapport à un autre (ce qui n’empêche pas la présence d’allèles dominants et récessifs pour ces gènes).
L’adjectif « récessive » ou « dominante » implique que l’inhibition de l’expression du gène dominé (par le gène dominant) soit le fait de l’allèle récessif ou de l’allèle dominant, respectivement, du gène dominant.
Épistasie faisant intervenir deux gènes et deux allèles par gène : toutes les possibilités
En cas d’épistasie, il existe six cas de figure mettant en jeu deux gènes comportant chacun deux allèles différents. Chacun de ces cas de figure entraîne des résultats spécifiques en F2. Dans la description des six cas de figures, les deux gènes seront appelés G1 et G2, l’allèle dominant sera noté « G » et l’allèle récessif « g », et en cas de besoin le gène épistatique sera le G1 et l’hypostatique le G2. Le phénotype est donné entre crochets.
Cas numéro 1 : l’épistasie dominante
Le gène G1 est épistatique au gène G2 par son allèle dominant G1.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1] |
G1G1 G2g2 [G1] |
G1g1 G2G2 [G1] |
G1g1 G2g2 [G1] |
| G1 g2 | G1G1 G2g2 [G1] |
G1G1 g2g2 [G1] |
G1g1 G2g2 [G1] |
G1g1 g2g2 [G1] |
| g1 G2 | G1g1 G2G2 [G1] |
G1g1 G2g2 [G1] |
g1g1 G2G2 [g1G2] |
g1g1 G2g2 [g1G2] |
| g1 g2 | G1g1 G2g2 [G1] |
G1g1 g2g2 [G1] |
g1g1 G2g2 [g1G2] |
g1g1 g2g2 [g1g2] |
On obtient 12/16e de phénotype [G1], 3/16e de phénotype [g1G2] et 1/16e de phénotype [g1g2], d’où un rapport de 12 : 3 : 1.
Cas numéro 2 : l’épistasie récessive
Le gène G1 est épistatique au gène G2, par son allèle récessif g1.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1G2] |
G1G1 G2g2 [G1G2] |
G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
| G1 g2 | G1G1 G2g2 [G1G2] |
G1G1 g2g2 [G1g2] |
G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1g2] |
| g1 G2 | G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
g1g1 G2G2 [g1] |
g1g1 G2g2 [g1] |
| g1 g2 | G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1g2] |
g1g1 G2g2 [g1] |
g1g1 g2g2 [g1] |
On obtient 9/16e de phénotype [G1G2], 3/16e de phénotype [G1g2] et 4/16e de phénotype [g1], d’où un rapport de 9 : 3 : 4.
Cas numéro 3 : l’effet cumulatif de deux gènes
La présence des allèles G1 OU G2 donne le même phénotype, différent de la présence de G1 ET G2.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1G2] |
G1G1 G2g2 [G1G2] |
G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
| G1 g2 | G1G1 G2g2 [G1G2] |
G1G1 g2g2 [G1,2] |
G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1,2] |
| g1 G2 | G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
g1g1 G2G2 [G1,2] |
g1g1 G2g2 [G1,2] |
| g1 g2 | G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1,2] |
g1g1 G2g2 [G1,2] |
g1g1 g2g2 [g1g2] |
| [G1,2] correspond au phénotype commun à la présence de l'allèle G1 OU de l'allèle G2. L'homozygotie ou l'hétérozygotie n'intervient pas dans le résultat. | ||||
On obtient 9/16e de phénotype [G1G2], 6/16e de phénotype [G1,2] et 1/16e de phénotype [g1g2], d’où un rapport de 9 : 6 : 1.
Cas numéro 4 : pas d’effet cumulatif (premier cas de figure)
La présence des allèles G1 ET/OU G2 donne le même phénotype.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1G2] |
G1G1 G2g2 [G1G2] |
G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
| G1 g2 | G1G1 G2g2 [G1G2] |
G1G1 g2g2 [G1G2] |
G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1G2] |
| g1 G2 | G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
g1g1 G2G2 [G1G2] |
g1g1 G2g2 [G1G2] |
| g1 g2 | G1g1 G2g2 [G1G2] |
G1g1 g2g2 [G1G2] |
g1g1 G2g2 [G1G2] |
g1g1 g2g2 [g1g2] |
On obtient 15/16e de phénotype [G1G2], et 1/16e de phénotype [g1g2], d’où un rapport de 15 : 1.
Cas numéro 5 : pas d’effet cumulatif (deuxième cas de figure)
La présence de g1g1 ET/OU g2g2 donne le même phénotype.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1G2] |
G1G1 G2g2 [G1G2] |
G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
| G1 g2 | G1G1 G2g2 [G1G2] |
G1G1 g2g2 [g1g2] |
G1g1 G2g2 [G1G2] |
G1g1 g2g2 [g1g2] |
| g1 G2 | G1g1 G2G2 [G1G2] |
G1g1 G2g2 [G1G2] |
g1g1 G2G2 [g1g2] |
g1g1 G2g2 [g1g2] |
| g1 g2 | G1g1 G2g2 [G1G2] |
G1g1 g2g2 [g1g2] |
g1g1 G2g2 [g1g2] |
g1g1 g2g2 [g1g2] |
On obtient 9/16e de phénotype [G1G2], et 7/16e de phénotype [g1g2], d’où un rapport de 9 : 7.
Cas numéro 6 : pas d’effet cumulatif (troisième cas de figure)
La présence de l’allèle G1 OU le double récessif g1g1 g2g2 donne le même phénotype.
| G1 G2 | G1 g2 | g1 G2 | g1 g2 | |
| G1 G2 | G1G1 G2G2 [G1] |
G1G1 G2g2 [G1] |
G1g1 G2G2 [G1] |
G1g1 G2g2 [G1] |
| G1 g2 | G1G1 G2g2 [G1] |
G1G1 g2g2 [G1] |
G1g1 G2g2 [G1] |
G1g1 g2g2 [G1] |
| g1 G2 | G1g1 G2G2 [G1] |
G1g1 G2g2 [G1] |
g1g1 G2G2 [G2] |
g1g1 G2g2 [G2] |
| g1 g2 | G1g1 G2g2 [G1] |
G1g1 g2g2 [G1] |
g1g1 G2g2 [G2] |
g1g1 g2g2 [G1] |
On obtient 13/16e de phénotype [G1], et 3/16e de phénotype [G2], d’où un rapport de 13 : 3.