

Dans ce document, après avoir défini ce que l'on entend par le processus culturel, est abordée la question de l'existence d'hérédité culturelle dans le monde vivant, au-delà de la seule espèce humaine. En effet, le processus culturel est trop souvent considéré comme l'apanage des humains, plus éventuellement quelques espèces de primates, voire quelques espèces d'oiseaux. Des découvertes récentes suggèrent que ce processus existe en fait chez un très grand nombre d'espèces, y compris chez des invertébrés. Ainsi, le processus culturel qui génère une autre forme d'hérédité apparaît-il comme étant un processus très général d'hérédité qui est donc susceptible d'avoir participé à l'évolution d'un très grand nombre d'espèces probablement depuis des millions d'années. Le processus culturel constitue donc un formidable processus de création et de maintien de la diversité biologique qu'il est nécessaire de prendre en compte en biologie en général et en biologie de la conservation en particulier.
Un des enjeux majeurs en biologie, est de comprendre pourquoi les individus d'une population, ou même au sein des fratries, diffèrent autant les uns des autres. Quels sont les mécanismes qui produisent et maintiennent cette diversité biologique ? Le débat scientifique sur ces sujets n’a jamais vraiment cessé et reste au cœur de la biologie moderne. Ce débat est fortement lié à la question de l'hérédité, c'est-à-dire au fait que les enfants ressemblent à leurs parents. À l'échelle d'une population, cette ressemblance se traduit par le fait que les enfants héritent des caractéristiques de leurs parents. Par exemple, les parents de grande taille ont en moyenne des enfants plus grands que ceux des parents de petite taille. C'est l'hérédité des différences, que j'appelle l'héritabilité inclusive1 qui est une mesure statistique de la ressemblance parent-enfant au sein d'une population. Cette mesure de la ressemblance ne peut, elle, s'effectuer qu'à l'échelle d'une population. La vision classique selon laquelle cette ressemblance résulte essentiellement, si ce n'est exclusivement, de la transmission de la séquence de l'ADN des parents vers leurs enfants, demande à être élargie du fait de très nombreuses découvertes faites depuis le début du troisième millénaire. Un des domaines concerné est celui de l'existence de l'hérédité culturelle. Pour bien comprendre l'importance de la question de l'évolution culturelle, il nous faut non seulement comprendre l'hérédité, mais aussi son rôle dans le vivant et l'évolution. Comme le concept même d'hérédité est en train de changer rapidement, nous commencerons par le discuter.
L'hérédité est au centre de la biologie
L'étude de l'hérédité est au centre de la biologie pour plusieurs raisons fondamentales. Tout d'abord, étudier la ressemblance parent-enfant c'est étudier les sources de variation phénotypique. D'autre part, la ressemblance parent-enfant est au cœur même de l'évolution que ce soit par sélection naturelle ou par dérive. En effet, depuis Darwin [3] la sélection naturelle est définie comme un processus qui s'enclenche inévitablement dès lors que deux conditions sont remplies (a) il existe de la variation dans les populations, et (b) il existe une relation persistante au cours des générations entre la valeur des traits et la capacité à avoir des descendants (ce que l'on appelle la fitness, concept que je traduis par le mot aptitude2). Par exemple pour la taille, cette relation peut être que dans une population, ce sont les individus de grande taille qui survivent et se reproduisent mieux (ils ont donc une plus forte aptitude) que les moyens ou les petits. Cette deuxième clause est ce que l'on dénomme la pression de sélection. Cependant, ces deux conditions ne conduiront à évolution que si une troisième condition est remplie, à savoir que (c) le trait soit héritable, c’est-à-dire que la variation sur le trait soit transmise à la descendance (par exemple dans le cas de la taille, les grands donnent naissance à des grands et les petits à des petits). C'est pour cette raison que la question de l'hérédité (la transmission des caractères) est au centre de toute la biologie en général et de la biologie évolutive en particulier.
Divers mécanismes d'hérédité
Depuis la découverte de l'ADN et de ses fantastiques capacités de stockage d'information, la vision dominante de l'hérédité est qu'elle repose essentiellement, si ce n'est exclusivement, sur la transmission de la séquence de l'ADN. C'est ce que j'appelle la vision « séquencique » de l'hérédité qui consiste à réduire l'hérédité à la seule transmission de la séquence nucléotidique de l'ADN3. C'est la vision la plus largement admise encore aujourd'hui.
Cependant, de longue date, et surtout depuis le début du troisième millénaire, de nombreuses découvertes ont remis en cause cette vision qui réduit l'hérédité à la seule séquencique. En effet, il ressort de ces multiples découvertes que la vision séquencique de l'hérédité et de l'évolution ne permet pas d'expliquer toute la complexité du vivant4. De nombreux arguments permettent aujourd'hui d'affirmer que la ressemblance parent-enfant ne repose pas uniquement sur le transfert de l'information séquencique mais implique aussi toute une série de processus variés de nature non séquencique (ou non génétique). Ces autres modes d'hérédité prennent plusieurs formes extrêmement variées, sophistiquées et surprenantes5. Il s'agit des hérédités épigénétique, cytoplasmique, culturelle et écologique, ou encore de la transmission du microbiote ou celle des prions et de ce que l'on appelle les molécules chaperons qui maintiennent les protéines dans une configuration fonctionnelle. Alors que toutes ces formes d'hérédité se transmettent uniquement de parent à enfant (ce que l'on appelle la transmission verticale), l'hérédité culturelle est très originale en ce sens qu'elle est la seule à pouvoir se transmettre verticalement mais aussi horizontalement (entre individus de la même cohorte) ou obliquement (entre individus non apparentés de générations différentes). Ainsi, notre conception de l'hérédité doit être élargie pour inclure dans les raisonnements de biologie toutes les composantes de l'hérédité, au-delà de la seule transmission séquencique. Cet élargissement de notre conception de l'hérédité des différences (appelée héritabilité inclusive) a le potentiel de fortement modifier notre vision du fonctionnement du vivant et de son évolution car, comme nous l'avons vu plus haut, l'hérédité des différences est la condition centrale pour que la sélection naturelle conduise à l'évolution.
La mise en évidence du fait que la ressemblance parent-enfant ne repose pas uniquement sur une variation de la séquence de l'ADN a émergé comme un des domaines majeurs de la biologie depuis le début du troisième millénaire. Le lecteur intéressé par la question de l'importance de l'hérédité non séquencique pourra par exemple lire [2, 6-13].
La culture animale
Fort de ces importants éléments de contexte, et armé de ces notions très générales de source de variation et d'hérédité, nous pouvons nous focaliser sur les variants culturels rendus héritables par l'apprentissage social.
Un peu d'histoire
Pendant très longtemps nous avons considéré que la culture était l'apanage de l'humanité. De ce fait, toutes les définitions étaient conçues pour capturer toutes les particularités de la culture humaine. Il n'y avait donc pas d'intérêt à rechercher l'existence de phénomènes culturels en dehors de notre espèce, et comme on ne trouve que ce que l'on recherche, nous n'avions pendant très longtemps aucun exemple de transmission culturelle en dehors de notre espèce.

Cependant, des faits ont été rapportés dès le début du XXe siècle qui donnaient à penser que le phénomène culturel avait une portée taxonomique s'étendant bien au-delà de la seule espèce humaine. Le premier exemple concernait des oiseaux. En effet, en plusieurs endroits de Grande-Bretagne dans les années 1900, des mésanges ont commencé à percer l'opercule des bouteilles de lait déposées chaque matin sur le perron des habitations pour se nourrir de la crème qui surnage sur le lait (Figure 1). Cette innovation comportementale s'est ensuite étendue rapidement à quasiment toute la Grande-Bretagne (Figure 2). Pour plus de détails sur les localités impliquées et sur la dynamique d'invasion de ce comportement, on peut consulter [14] qui fournit des cartes accessibles même aux non anglophones1. Ce phénomène a donné lieu à de nombreuses discussions avant que ne fasse consensus l'idée que cette rapide diffusion du comportement relevait bien d'un processus culturel2.
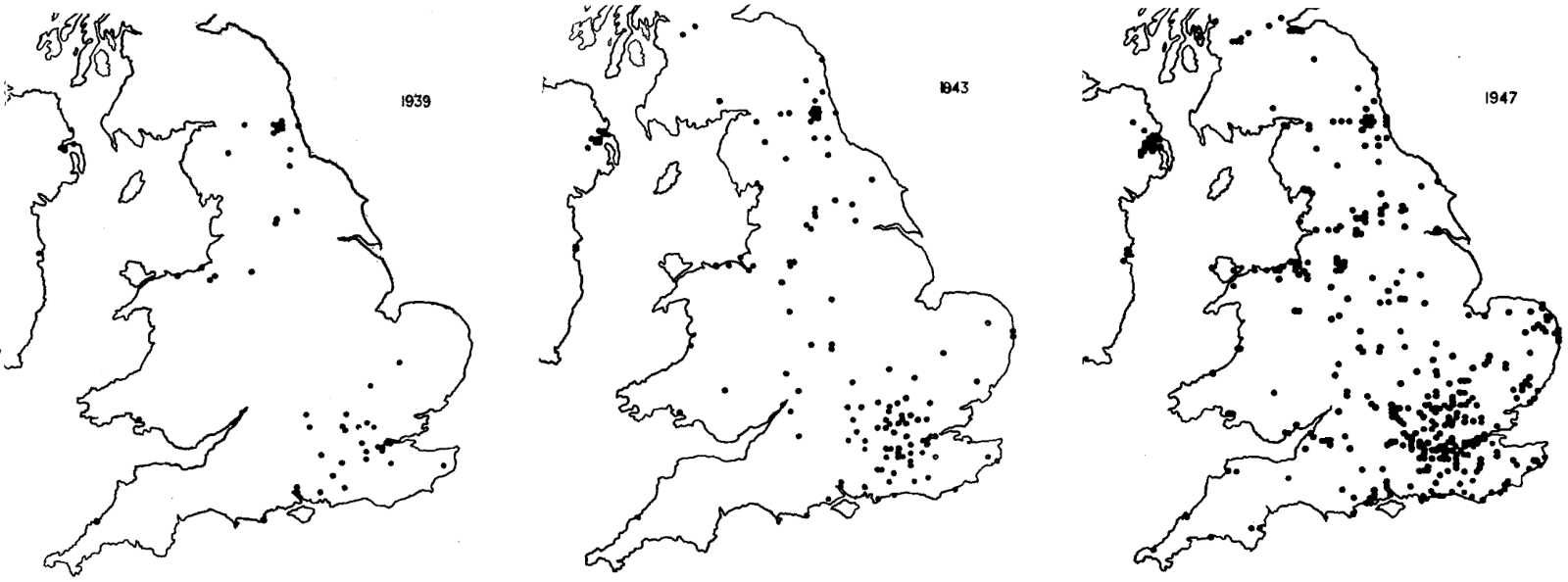
Chaque carte présente la totalité des cas recensés jusqu’à l’année indiquée. Le terme « mésanges » regroupe les espèces Parus major (mésange charbonnière), Periparus ater (mésange noire) et Cyanistes caeruleus (mésange bleue).
Le deuxième exemple classique est celui des macaques de l'île de Koshima, au Japon [16]. Au milieu du XXe siècle des primatologues déposèrent régulièrement dans des endroits faciles à observer des patates douces dont se nourrissent les singes. Une femelle appelée Imo s'est alors mise à laver les tubercules pour enlever le sable qui était dessus, ce qui en facilitait grandement la consommation. Cette innovation s'est alors propagée à tous les membres de son groupe de macaques (Figure 3). Il s'en est suivi toute une série d'autres innovations qui se sont accumulées au cours du temps et qui ont profondément changé le mode de vie de ces singes et donc les pressions de sélection agissant sur eux [16, 17]. De nouveau, ce cas a été discuté pour savoir si l'on pouvait là aussi parler de culture animale, soulevant encore une fois la question de ce qu'est la culture.
Arbre généalogique de macaques japonais (Macaca fuscata) de l’île de Koshima (Japon). Parmi ces individus, certains vont au bord de l’eau pour débarrasser les patates douces du sable qui les couvre. Un autre comportement observé dans cette population de macaques est l’« orpaillage » du blé : au lieu de manger un par un les grains de blé qu’ils trouvent dans le sable, certains individus jettent le mélange sable/blé dans l’eau. Le sable coule tandis que le blé flotte et est alors récupéré pour être consommé.
Pendant longtemps ce furent les deux seuls exemples illustrant la possibilité de culture animale. Depuis cette époque, toute une série d'exemples de transmission culturelle ont été décrits chez les chimpanzés [18-22], chez les orang-outans [23], le singe vervet [24], plusieurs cétacés [25-28], les suricates [29], diverses espèces d'oiseaux [30, 31] et d'une manière encore plus surprenante chez les insectes. Les études sur les oiseaux sont remarquables car elles sont expérimentales et étudient la dynamique de transmission intrapopulation, l'une sur le chant [30], l'autre sur l'exploitation d'une source artificielle de nourriture [31]. Pour les insectes les arguments expérimentaux portent sur la transmission à l'intérieur d'une ruche d'un nouveau comportement d'alimentation chez un bourdon [32], et la transmission des préférences sexuelles chez la mouche du vinaigre [33]. Voir aussi [17, 34].
Une définition centrée sur les invariants du processus culturel
Toutes ces discussions posent en fait la question de la définition de la culture. Il fallait pour cela se dégager de notre anthropocentrisme de base pour nous concentrer sur ce qu'il y a d'essentiel dans le processus culturel. Le point sur lequel s'accorderait la majorité des chercheurs est que, dans sa version princeps, la culture est quelque chose qui est appris depuis les autres. Ainsi, le phénomène culturel est intimement lié à la question de l'apprentissage social, domaine pour lequel il existe une vaste littérature. Un des premiers exemples d'apprentissage social décrit dans la littérature est celui de la danse des abeilles [35]. Depuis cette date, de très nombreux autres exemples ont été publiés (voir par exemple [17,36-47]).
Les premières définitions de la culture animale se focalisaient sur les patrons de variation comportementale entre populations. Par exemple, pour une tâche donnée, les individus d'une population se comportent d'une certaine manière (utilisation de pierres pour casser des noix) alors que dans d'autres populations leurs congénères se comportent différemment (utilisation de bâtons pour casser des noix) alors que les conditions écologiques et génétiques sont très semblables. Comme ces différences comportementales persistaient au cours des générations, il semblait qu'il s'agissait de véritables traditions culturelles. Cependant, la seule présence de patrons de variation ne permet pas d'inférer les mécanismes responsables de cette variation. Il fallait bien entendu aussi démontrer que ces variations sont effectivement transmises entre les individus par apprentissage social, c'est-à-dire par apprentissage à partir des autres.
Une définition de la culture transposable à toutes les espèces
Nous avons donc proposé une nouvelle définition de la culture qui associe à cette approche centrée sur les patrons de variation (les traditions) une étude des propriétés de l'apprentissage social sous-jacent afin de vérifier que cet apprentissage social génère effectivement l'émergence de comportements communs au sein d'une même population, mais variables entre populations [33].
Avec mes coauteurs de cet article, nous proposons donc que « la culture est la part de la variation entre individus qui est transmise par (1) un apprentissage social qui (2) se fait régulièrement des plus anciens vers les plus jeunes, (3) est mémorisée assez longtemps pour être de nouveau copiée, (4) qui est basée sur les traits plutôt que sur les individus1 et qui (5) incorpore un mécanisme de réparation comme le conformisme2. Enfin, il faut aussi vérifier que ces cinq conditions réunies suffisent à faire émerger des comportements collectifs similaires localement et persistants sur de très longues périodes, bien au-delà de la durée d'une génération. Il faut donc que ces caractéristiques cognitives aient le potentiel de faire émerger des traditions culturelles persistantes. » Cette définition est résumée dans la Figure 4.
Ainsi caractérisée de manière rigoureuse et complète, il devient maintenant possible d'étudier la culture animale chez n'importe quel organisme, cette définition constituant une boîte à outils conceptuelle et pratique pour étudier la culture dans le monde animal.
Des drosophiles culturelles
Pour tester cette définition, nous avions choisi un organisme tout à fait improbable a priori, à savoir la drosophile (Drosophila melanogaster) [33]1. Cette petite mouche mesure en effet de l'ordre du millimètre et a un cerveau qui est environ dix millions de fois plus petit que le nôtre. De manière très surprenante, nous avons observé que les préférences sexuelles des drosophiles femelles sont transmises socialement entre femelles et que cet apprentissage social vérifie tous les critères de notre définition (Figure 5). De plus, dans une expérience de chaîne de transmission dans laquelle les observatrices (élèves) d'un pas de transmission devenaient les démonstratrices (professeurs) du pas suivant, nous avons constaté que de telles chaines duraient significativement plus longtemps qu'attendu en l'absence d'apprentissage social. Enfin, des simulations statistiques nous ont permis de montrer que les caractéristiques de l'apprentissage social sont effectivement susceptibles de faire émerger une préférence collective pour un type de mâle donné, certaines de ces traditions pouvant persister sur plus de 100 000 pas de transmission, ce qui pouvait représenter près de 10 000 générations de drosophiles (pour tous les détails, voir [33]).
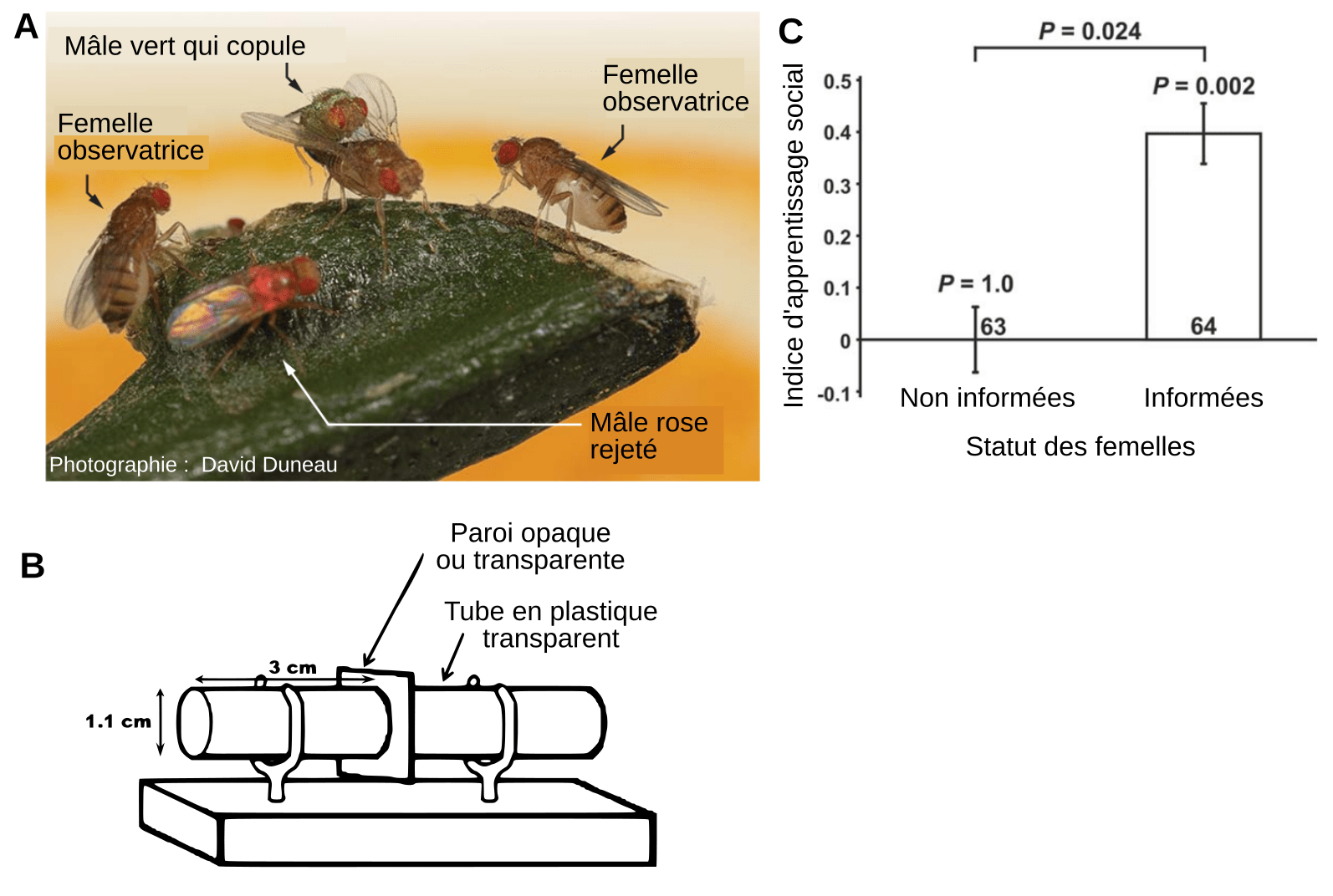
A. Le mâle poudré de vert copule, mais pas le mâle poudré de rose. Deux femelles semblent observer le couple en train de copuler. B. Dispositif expérimental permettant de reproduire la situation présentée en A. Une femelle observatrice est placée dans un tube en plastique d'où elle peut observer (paroi transparente) ou non (paroi opaque) la démonstration (impliquant une femelle et un mâle poudré de rose ou de vert) se déroulant dans l'autre tube. Ensuite les individus impliqués dans la démonstration sont enlevés et remplacés par deux nouveaux mâles, un de chaque couleur, la paroi est enlevée et on enregistre la couleur du mâle avec lequel la femelle observatrice copule. C. Indice d’apprentissage sociale de femelles n’ayant pas observé (paroi opaque utilisée en B) ou ayant observé (paroi transparente utilisée en B) une femelle démonstratrice s'accoupler avec un mâle d’une couleur donnée. Un indice valant zéro témoigne du choix aléatoire du partenaire tandis qu’un indice positif traduit une préférence pour la couleur du mâle observé durant la démonstration. Les valeurs P des tests au-dessus des barres sont celles de tests binomiaux par rapport à un choix aléatoire. Les barres d’erreur représentent les erreurs types de la moyenne.
D'autre part, nous nous attendions à ce que la transmission des préférences sexuelles se fasse culturellement car, dans ce contexte, le conformisme apporte un avantage chaque fois qu'un individu change de population (ou de groupe culturel). En effet, le conformisme permet à un individu qui change de groupe culturel de toujours adopter le comportement majoritaire localement, indépendamment du comportement majoritaire de son groupe d’origine. De ce fait, les femelles conformistes produiront toujours des fils du phénotype qui est préféré dans le groupe culturel de la reproduction en cours, ce qui augmentera fortement leur aptitude. Nos expériences ont confirmé magistralement le caractère conformiste des drosophiles femelles dans le choix de leurs partenaires.
Ainsi les drosophiles ont toutes les capacités cognitives nécessaires à l'émergence de véritables traditions locales de préférence sexuelle pour un type de mâle donné, ce qui suggère que la transmission culturelle est beaucoup plus ancienne et répandue que nous ne le pensions, ou qu’elle résulte de phénomènes de convergence entre les différents groupes étudiés. Dans la première hypothèse, la culture pourrait donc avoir participé à l'hérédité et l'évolution depuis des millions d'années et ceci chez un panel d'animaux bien plus varié que nous ne pouvions l'imaginer.
Conséquences
Une première conséquence de la probable omniprésence d'hérédité culturelle dans le monde animal est qu'elle a le potentiel de fortement affecter la sélection sexuelle et la spéciation chez les organismes où les préférences sexuelles sont héritées culturellement. Une autre conséquence de l'existence d'une composante culturelle dans la ressemblance parent-enfant est qu'il est nécessaire de revoir la définition de l'évolution. Alors que classiquement l'évolution est définie comme « le processus par lequel la fréquence des variants génétiques change au cours du temps », il nous faut maintenant adopter une définition plus inclusive qui incorpore l'ensemble des processus participant à la ressemblance parent-enfant. Pour cela il suffit d'enlever un seul mot: « l'évolution est le processus par lequel la fréquence des variants change au cours du temps »1 [49].
Dans cette nouvelle définition de l'évolution, le mot variant inclus tous les variant possibles, dont bien entendu les variants séquenciques (c'est-à-dire dans la séquence nucléotidique de l'ADN). Mais il inclut aussi le rôle de toute une série de variants phénotypiques non génétiques mais néanmoins transmis entre générations, et dont le rôle majeur (et largement sous-estimé) dans l'hérédité et l'évolution est actuellement mis au jour. Ces variants peuvent être épigénétiques, cytoplasmiques, microbiotiques, culturels, et écologiques, ou dus à des prions ou des molécules chaperons. Comme toutes ces variations ont des composantes transmises des parents vers leurs enfants (c'est-à-dire qu'elles sont en partie inclusivement héritables), ces variants sont susceptibles d'évoluer dans le temps par l'effet de la sélection naturelle ou de la dérive, et donc de participer à l'évolution.
Enfin, alors que nous savons depuis longtemps que le comportement participe à l'évolution en permettant aux organismes de s'adapter rapidement aux changements dans leur environnement, il apparaît aujourd'hui que le comportement constitue aussi un important acteur d'hérédité ayant des propriétés de transmission très originales (intra mais aussi inter lignées familiales) qui sont susceptibles d'affecter le fonctionnement même de l'évolution. De plus, plusieurs auteurs ([50-53] ; pour une revue, voir [54]) ont insisté sur la nécessité de prendre en compte les variants culturels dans toutes nos politiques de conservation afin de conserver au maximum le potentiel évolutif des espèces et espérer ainsi tamponner autant que possible les effets délétères des changements globaux sur la diversité biologique2.
Références
1. Danchin, É., and Wagner, R.H. (2010) Inclusive heritability: combining genetic and nongenetic information to study animal behavior and culture. Oikos 119, 210-218.
2. Danchin, É., et al. (2011) Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nature Rev Genet 12, 475-486.
3. Darwin, C. (1859) On the origin of species by means of natural selection. John Murray.
4. Danchin, É., et al. (2005) Ecologie comportementale. Dunod.
5. Danchin, É., et al.(2008) Behavioural Ecology. Oxford University Press.
6. Danchin, É., et al. (2019) Early in life effects and heredity: reconciling neo-Darwinism with neo-Lamarckism under the banner of the inclusive evolutionary synthesis. Phil. Trans. R. Soc. B 374, 20180113.
7. Bonduriansky, R., and Day, T. (2018) Extended Heredity: A New Understanding of Inheritance and Evolution. Princeton University Press.
8. Pigliucci, M., and Müller, G.B. (2010) Evolution, the extended synthesis. MIT Press.
9. Bonduriansky, R. (2012) Rethinking heredity, again. Trends Ecol Evol 27, 330-336.
10. Wang, Y., et al. (2017) Lamarck rises from his grave: parental environment-induced epigenetic inheritance in model organisms and humans. Biol Rev 92, 2084-2111.
11. Danchin, É., et al. (2019) Epigenetically-Facilitated mutational assimilation: Epigenetics as a hub within the Inclusive Evolutionary Synthesis. Biol Rev 94, 259-282.
12. Lu, Q., and Bourrat, P. (2017) The evolutionary gene and the extended evolutionary synthesis. British J Phil Sci 69, 775-800.
13. Huneman, P., and Whalsh, D.M. (2017) Challenging the Modern Synthesis. Adaptation, Development, and Inheritance. Oxford University Press.
14. Fisher, J., and Hinde, R.A. (1949) The opening of milk bottles by birds. Brit Birds 42, 347.
15. Sherry, D.F., and Galef, B.G. (1984) Cultural transmision without imitation - Milk bottle opening by birds. Anim Behav 32, 937-938.
16. Hirata, S., et al. (2008) "Sweet-potato washing" revisited. In Primate origins of human cognition and behavior (T., M., ed), pp. 487-508, Springer.
17. Avital, E., and Jablonka, E. (2000) Animal Traditions. Behavioural Inheritance in Evolution. Cambridge University Press.
18. Whiten, A., et al. (1999) Culture in chimpanzees. Nature 399, 682-685.
19. Whiten, A. (2017) Culture extends the scope of evolutionary biology in the great apes. Proc Nat Acad Sci USA 114, 7790-7797.
20. Whiten, A. (2011) The scope of culture in chimpanzees, humans and ancestral apes. Philos Trans R Soc Lond B 366, 997-1007.
21. Whiten, A., et al. (2005) Conformity to cultural norms of tool use in chimpanzees. Nature 437, 737-740.
22. Whiten, A. (2007) Pan African culture: Memes and genes in wild chimpanzees. Proc Nat Acad Sci USA 104, 17559–17560.
23. van Schaik, C.P., et al. (2003) Orangutan cultures and the evolution of material culture. Science 299, 102-105.
24. van de Waal, E., et al. (2013) Potent Social Learning and Conformity Shape a Wild Primate's Foraging Decisions. Science 340, 483-485.
25. Whitehead, H. (1998) Cultural selection and genetic diversity in matrilineal whales. Science 282, 1708-1711.
26. Allen, J., et al. (2013) Network-Based Diffusion Analysis Reveals Cultural Transmission of Lobtail Feeding in Humpback Whales. Science 340, 485-488.
27. Krutzen, M., et al. (2005) Cultural transmission of tool use in bottlenose dolphins. Proc Nat Acad Sci USA 102, 8939-8943.
28. Kopps, A.M., et al. (2014) Cultural transmission of tool use combined with habitat specializations leads to fine-scale genetic structure in bottlenose dolphins. Proc R Soc Lond B Biol Sci 281, 20133245.
29. Thornton, A., et al. (2010) Multi-generational persistence of traditions in neighbouring meerkat groups. Proc R Soc Lond B Biol Sci 277, 3623-3629.
30. Feher, O., et al. (2009) De novo establishment of wild-type song culture in the zebra finch. Nature 459, 564-U594.
31. Aplin, L.M., et al. (2015) Experimentally induced innovations lead to persistent culture via conformity in wild birds. Nature 518, 538-541.
32. Alem, S., et al. (2016) Associative Mechanisms Allow for Social Learning and Cultural Transmission of String Pulling in an Insect. PloS Biol 14, e1002564.
33. Danchin, É., et al. (2018) Cultural flies: conformist social learning in fruit flies predicts long-lasting mate-choice traditions. Science 362, 1025-1030.
34. Danchin, É., et al. (2004) Public information: from nosy neighbors to cultural evolution. Science 305, 487-491.
35. Von Frisch, K. (1974) Decoding the language of the bee. Science 185, 663-668.
36. Heyes, C.M., and Galef, B.G.J. (1996) Social learning and Imitation: the Roots of Culture. Academic Press.
37. Brown, C.R., and Laland, K.N. (2003) Social learning in fishes: a review. 4, 280-288.
38. Griffin, A.S. (2004) Social learning about predators: A review and prospectus. Learnin Behav 32, 131-140.
39. Chittka, L., and Leadbeater, E. (2005) Social learning: Public Information in Insects. Curr Biol 15, R869-R871.
40. Coolen, I., et al. (2005) Social learning in noncolonial insects? Curr Biol 15, 1931-1935.
41. Leadbeater, E., and Chittka, L. (2007) Social learning in insects - From miniature brains to consensus building. Curr Biol 17, R703-R713.
42. Mery, F., et al. (2009) Public versus personal information for mate copying in an invertebrate. Curr Biol 19, 730-734.
43. Slagsvold, T., and Wiebe, K.L. (2011) Social learning in birds and its role in shaping a foraging niche. Philos Trans R Soc Lond B 366, 969-977
44. Battesti, M., et al.(2012) Spread of Social Information and Dynamics of Social Transmission within Drosophila Groups. Curr Biol DOI 10.1016/j.cub.2011.12.050
45. Watson, S.K., et al. (2015) Vocal Learning in the Functionally Referential Food Grunts of Chimpanzees. Curr Biol
46. Dagaeff, A.-C., et al. (2016) Drosophila mate copying correlates with atmospheric pressure in a speed learning situation. Anim Behav 121, 163-174.
47. Whiten, A. (2018) Culture and conformity shape fruitfly mating. Potent social learning sustains the inheritance of mating preferences over generations. Science 362, 998-999.
48. Boyd, R., and Richerson, P.J. (1985) Culture and the evolutionary process. Chicago University Press.
49. Bentley, R.A., et al. (2004) Random drift and culture change. Proc R Soc Lond B Biol Sci 271, S353-S356.
50. Whitehead, H., et al. (2004) Culture and conservation of non-humans with reference to whales and dolphins: review and new directions. Biol Cons 120, 427-437.
51. Laiolo, P., and Tella, J.L. (2005) Habitat fragmentation affects culture transmission: patterns of song matching in Dupont's lark. J Anim Ecol 42, 1183-1193.
52. Laiolo, P., and Tella, J.L. (2007) Erosion of animal cultures in fragmented landscapes. 5, 68-72.
53. Laiolo, P., and Jovani, R. (2007) The emergence of animal culture conservation. Trends Ecol Evol 22, 5-5.
54. Brakes, P., et al. (2019) Animal cultures matter for conservation. Science
55. Jesmer, B.R., et al. (2018) Is ungulate migration culturally transmitted? Evidence of social learning from translocated animals. Science 361, 1023-1025.



