

Dans une optique d’amélioration du troupeau, sélectionner les reproducteurs en se fondant sur leurs performances propres n’est pas forcément la meilleure méthode. Une estimation de la valeur génétique des candidats à la reproduction peut permettre d’obtenir de meilleurs résultats.
Cet article traite de la sélection génétique « classique », qui repose sur l’estimation de valeurs génétiques à partir des phénotypes des candidats ou de leurs apparentés. Depuis plusieurs années, la sélection génomique offre une approche différente de l’amélioration génétique des animaux d’élevage. Voir l’article de Didier Boichard, La sélection génomique chez la vache laitière.
Dans un troupeau, le phénotype des animaux vis-à-vis de différents traits (taille, fertilité…) dépend à la fois de facteurs génétiques et de facteurs environnementaux (quantité de nourriture disponible, aléas météorologiques, exposition à des maladies…). Pour augmenter la productivité d’un élevage, il est bien sûr possible d’améliorer les conditions environnementales : prévention des maladies, augmentation de la ration alimentaire, etc. La réponse est alors rapide mais limitée à la période d’application de ces conditions… À environnement constant, la sélection génétique permet d’améliorer les qualités d’un troupeau, et cette amélioration obtenue à un moment donné est permanente et cumulative. La sélection génétique consiste à choisir parmi les individus composant le troupeau (appelés candidats), ceux qui répondent le mieux aux objectifs des éleveurs et éleveuses (augmentation de la fertilité, diminution des maladies…), et qui seront donc utilisés en tant que reproducteurs pour procréer la génération suivante. Comment procéder en pratique ?
Les caractères d’intérêt
Les caractères d’intérêt varient selon les espèces. En élevage bovin laitier, qui servira de fil rouge à cet article, il s’agit en particulier de la quantité de lait produite, de sa composition (taux butyreux1, taux protéique), de la résistance aux mammites2, de la fertilité, de la longévité…
Ces caractères ainsi que, de manière générale, l’ensemble des traits d’intérêt agronomique sont des caractères quantitatifs polygéniques. Cela signifie qu’il s’agit de caractères déterminés par un grand nombre de gènes et qu’ils sont aussi affectés par le milieu. Souvent, ces caractères présentent dans la population une distribution se rapprochant d’une loi normale (Figure 1), même si ce n’est pas toujours le cas (cas de la fertilité par exemple, avec une distribution binaire succès/échec).
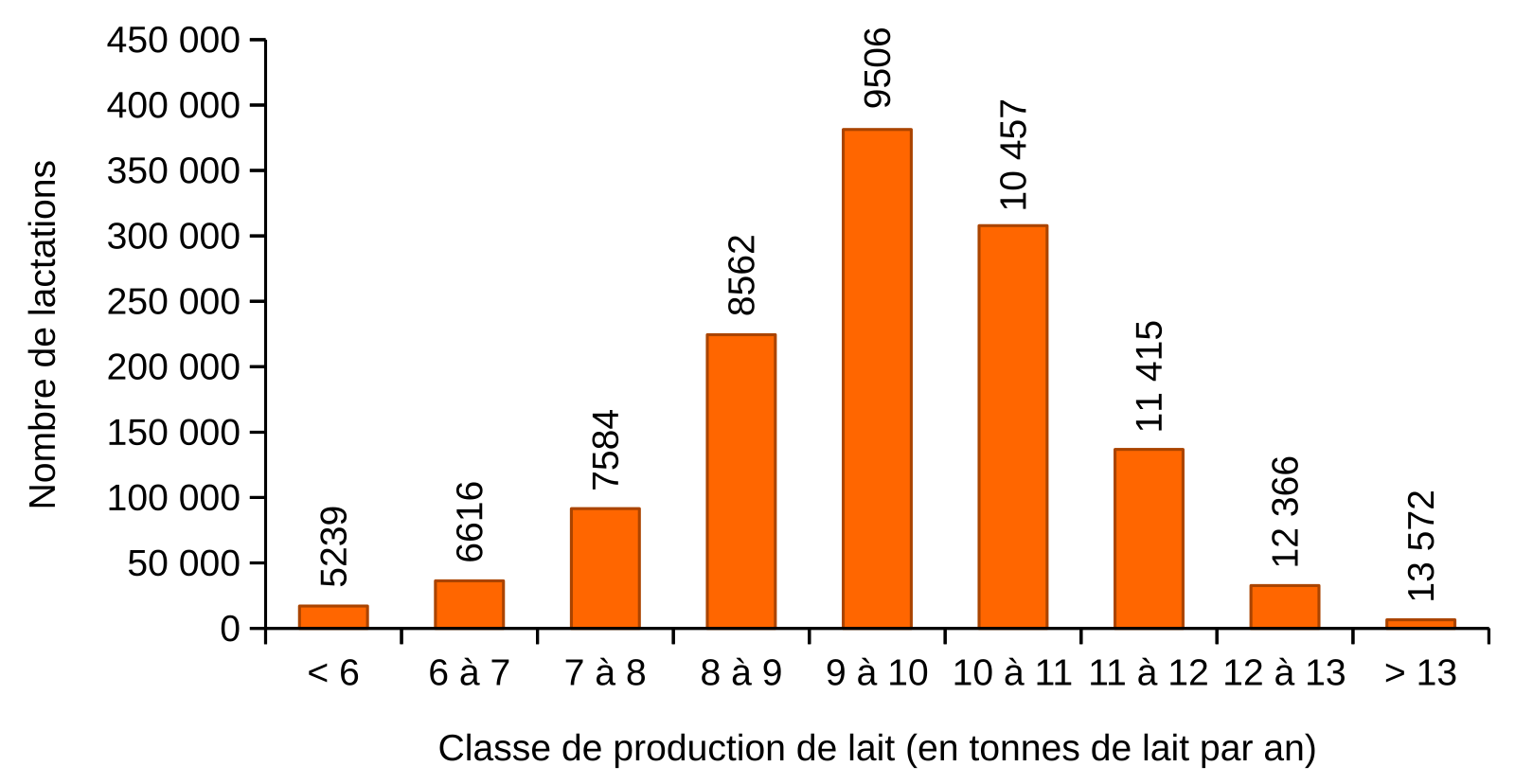
Pour la seule race Prim’Holstein, plus de 1,43 million de lactations ont été recensées en France métropolitaine pour l’année 2020. La distribution de ces lactations en fonction de la classe de production de lait est présentée. Les valeurs au-dessus des barres de l’histogramme représentent la moyenne de production de lait pour chaque classe (en L/an). La production moyenne d’une vache Prim’Holstein est de 9 594 L/an. À noter, la race Prim’Holstein est l’une des plus productives. Toutes races confondues, la production moyenne de lait d’une vache en France est de 7 060 L/an.
Source des données : Résultats de contrôle laitier, Espèce bovine, France 2020, Institut de l’élevage.
Sélectionner les meilleurs reproducteurs à partir de leurs phénotypes : la sélection massale
La solution la plus intuitive pour améliorer les caractéristiques d’un troupeau au cours du temps consiste à sélectionner comme reproducteurs, à chaque génération, les individus présentant les meilleures performances pour le caractère d’intérêt. Par exemple, si l’objectif est d’augmenter la production de lait, il semble logique de sélectionner comme reproductrices les vaches produisant le plus de lait. Ce type de sélection, fondée sur les performances propres des individus, est appelée sélection massale. Elle est probablement utilisée depuis les débuts de la domestication.
Cependant, cette option n’est pas forcément la solution optimale pour assurer l’amélioration génétique du troupeau et cela pour plusieurs raisons :
-
Le phénotype d’un individu ne dépend pas exclusivement de son génotype, il est également influencé par de nombreux facteurs non génétiques. Par exemple, une vache adulte produit plus qu’une vache en première lactation. Des facteurs comme l’âge à la mise bas ou la saison de mise bas ont également des effets importants. Les différences non génétiques les plus importantes sont entre élevages. Il convient donc de comparer des performances réalisées dans des conditions comparables ou, à défaut, corrigées pour les effets non génétiques. En effet, sélectionner des individus sur la base de leurs performances brutes (non corrigées) revient à sélectionner partiellement pour des effets de milieu qui ne sont pas transmis aux descendants.
-
Le caractère phénotypique d’intérêt peut ne pas être observable sur tous les animaux, ou pas encore observé sur les candidats à l’âge de leur sélection. Par exemple, les taureaux n’expriment pas la plupart des caractères sélectionnés, comme la quantité de lait ou la résistance aux mammites.
-
Enfin, seule une partie de la performance est d’origine génétique et se transmet à la descendance. Si cette partie, dite héritabilité, est réduite, la supériorité de la descendance sur les parents sera faible et il existe alors des méthodes de sélection plus efficaces que la sélection massale.
La sélection massale est donc particulièrement intéressante lorsque le phénotype est disponible sur tous les candidats, suffisamment tôt pour permettre une sélection, et lorsque l’héritabilité est élevée, c’est-à-dire lorsque le phénotype des parents reflète bien le potentiel génétique transmissible aux descendants. Lorsque ces conditions sont réunies, la sélection massale présente l’avantage d’être facile à mettre en œuvre puisqu’elle se fonde sur les phénotypes des candidats et qu’elle ne nécessite pas d’informations généaologiques.
L’héritabilité des caractères
Pour certains caractères, alors que les parents possèdent un phénotype supérieur à la moyenne du troupeau, les descendants présentent des valeurs dans la moyenne du troupeau, loin des performances de leurs parents. À l’inverse, pour d’autres traits, les performances des descendants sont bien supérieures à celles de la moyenne du troupeau, se rapprochant alors de celles des parents.
Ces observations mettent en évidence que la ressemblance entre parents et descendants varie selon les caractères, qui ne possèdent donc pas la même héritabilité. L’héritabilité, notée h2, se définit comme le rapport de la variance génétique additive sur la variance phénotypique : h2 = VA / VP. En théorie, la valeur de l’héritabilité peut varier de 0 (le caractère ne dépend que de l’environnement) à 1 (le caractère est uniquement dû à des effets génétiques additifs ; voir section suivante pour la définition de la variabilité génétique additive). En pratique, les valeurs d’héritabilité constatées chez les animaux d’élevage vont généralement de 0 à 0,6. Dans les élevages de bovins laitiers, les caractères liés à la fertilité sont peu héritables, tandis que ceux liés à la production de lait ont une héritabilité plus élevée (Tableau 1).
| Groupes de caractères | Héritabilité |
|---|---|
| Croissance | 0,3 |
| Format (taille des animaux) | 0,5 |
| Conformation (morphologie) | 0,15 – 0,5 |
| Production laitière | 0,3 |
| Composition du lait | 0,5 |
| Reproduction | 0,01 à 0,10 |
| Résistance aux maladies | 0,02 à 0,15 |
Sélectionner les meilleurs reproducteurs à partir de leur valeur génétique
Comme nous l’avons vu, la sélection massale n’est pas toujours possible ni efficace. D’autres modes de sélection existent, comme la sélection se fondant sur les valeurs génétiques.
La notion de valeur génétique
La performance d’un individu pour un caractère donné n’est pas simplement égale à la moyenne des performances de ses parents. En effet, en plus du rôle de l’environnement déjà évoqué, deux autres facteurs conditionnent la transmission des caractères :
-
Un reproducteur ne transmet que la moitié de son génome à sa descendance. L’autre moitié étant transmise par l’autre parent, des effets de dominance entre allèles paternels et maternels peuvent influencer positivement ou négativement le phénotype du descendant.
-
La production des gamètes fait intervenir les phénomènes de brassages interchromosomique et intrachromosomique, qui rompent, chez les descendants, les associations entre allèles qui existaient chez le parent. Chez les descendants, l’interaction entre allèles transmis (épistasie) contribue positivement ou négativement au phénotype.
Aussi, à l’échelle du troupeau, la variabilité phénotypique VP constatée entre les individus ne dépend pas que de la variabilité des allèles possédés, chacun ayant indépendamment un petit effet positif ou négatif sur le phénotype (cette part de variabilité dans l’explication de la variabilité phénotypique est appelée variabilité génétique additive, VA). Il faut également tenir compte de la variabilité associée aux phénomènes de dominance et d’épistasie, ainsi que de la variabilité de l’environnement.
Au final, on a donc : VP = VA + VD + VI + VE
avec :
- VP, la variabilité phénotypique
- VA, la variabilité génétique additive
- VD, la variabilité génétique due aux effets de dominance
- VI, la variabilité génétique due aux effets épistatiques
- VE, la variabilité due aux effets de l’environnement
C'est parce que le phénotype ne dépend pas que des effets génétiques additifs, mais qu'il est également sous l'influence des effets de dominance, d’épistasie et surtout de l’environnement que les valeurs d'héritabilité ne sont jamais égales à 1 (voir Tableau 1, section précédente).
La sélection génétique « classique » repose sur l'estimation de la valeur génétique des candidats à la reproduction. Cette valeur génétique représente la part de la performance d'un individu qui sera en moyenne transmise à ses descendants.
Estimer la valeur génétique d’un individu par testage sur descendance
Pour chaque trait d’intérêt (quantité de lait, taux butyreux, fertilité…), il est possible d’estimer pour un individu donné sa valeur génétique. Cette valeur génétique est estimée à partir de la valeur du trait mesuré chez sa descendance. La valeur génétique d’un individu est exprimée relativement à la performance moyenne du troupeau et peut donc être positive ou négative.
En système bovin laitier, du fait que les vaches ne peuvent produire que très peu de descendants au cours de leur vie, c’est en grande partie sur les taureaux que repose le potentiel d’amélioration génétique d’un troupeau, car ils ont un fort potentiel de diffusion. S’il est vrai qu’un taureau n’exprime pas la plupart des caractères que l’on souhaite sélectionner (quantité de lait, taux butyreux…), il est tout de même possible de lui attribuer des valeurs génétiques pour ces caractères. Ces valeurs génétiques peuvent en effet être estimées à partir des performances mesurées chez des apparentés. En pratique, l’estimation de la valeur génétique d’un taureau sur la base de son ascendance n’est pas suffisamment précise pour le sélectionner et le diffuser largement. En sélection génétique « classique », l’estimation de la valeur génétique d’un taureau se fait donc à partir des phénotypes mesurés chez ses filles, en mettant en place un testage sur descendance (Figure 2).
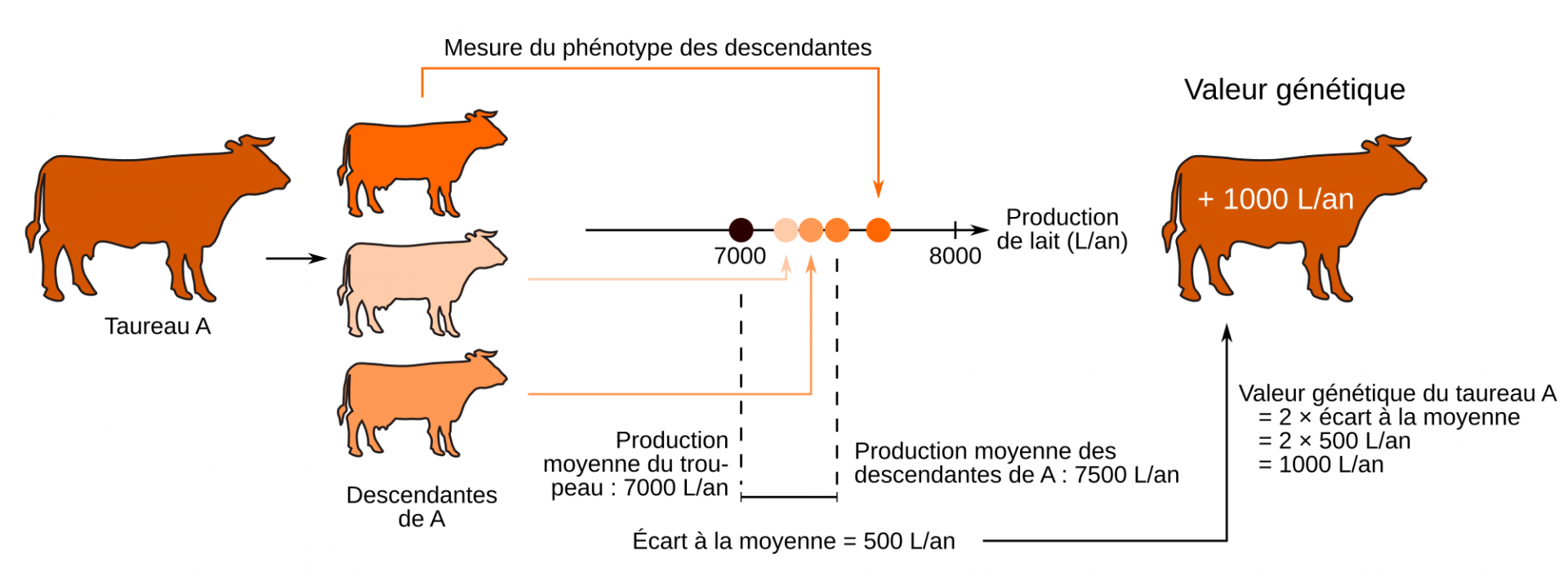
Pour estimer la valeur génétique d’un individu donné (ici le taureau A) pour un caractère donné (ici le caractère « production de lait »), on fait se reproduire cet individu. Le phénotype des descendants est ensuite mesuré. L’écart à la moyenne entre les performances des descendants et celles du troupeau, permet de calculer la valeur génétique de l’individu d’intérêt (ici le taureau A). En fonction du résultat obtenu, comparé à ceux des autres candidats, l’éleveur ou l’éleveuse décidera d’utiliser ou non l’animal comme reproducteur.
Source de la silhouette de vache : Kingroyos, CC BY-SA, Wikimedia
Alors que la figure 2 présente une estimation de la valeur génétique d’un taureau à partir de trois descendantes, c’est en réalité un échantillon de 50 à 100 filles qui est utilisé, chacune étant phénotypée pour tous les caractères d’intérêt (quantité de lait, taux protéique, fertilité, résistance aux mammites, conformation…). Une fois ces informations disponibles, il est possible d’établir un classement des taureaux et de sélectionner les meilleurs pour les diffuser largement.
En pratique, l’estimation de la valeur génétique d’un individu ne repose pas que sur les performances de sa descendance, mais plus généralement sur toutes les informations disponibles (performances propres, ascendance, frères et sœurs…), qui sont combinées pour déduire l’estimation de valeur génétique la plus précise possible à un moment donné. La fiabilité de l’estimation d’une valeur génétique, notée R2, augmente avec la quantité d’informations disponibles pour l’animal. La valeur de R2 traduit le carré de la corrélation entre la valeur génétique estimée (appelée index) et la valeur génétique « vraie » de l’animal. Plus R2 est proche de 1 et plus l’estimation est précise.
L’estimation de la valeur génétique d’un individu par testage sur descendance est une méthode qui a été utilisée pendant plus de 50 ans avec beaucoup d’efficacité. Elle présente cependant plusieurs inconvénients, dont sa lenteur (5 ans environ pour tester un taureau) et son coût (environ 40 000 euros pour chaque taureau, de sorte qu’avec un taux de sélection de 1 taureau sur 10, le taureau diffusé vaut 400 000 euros et doit être largement diffusé pour financer la sélection).
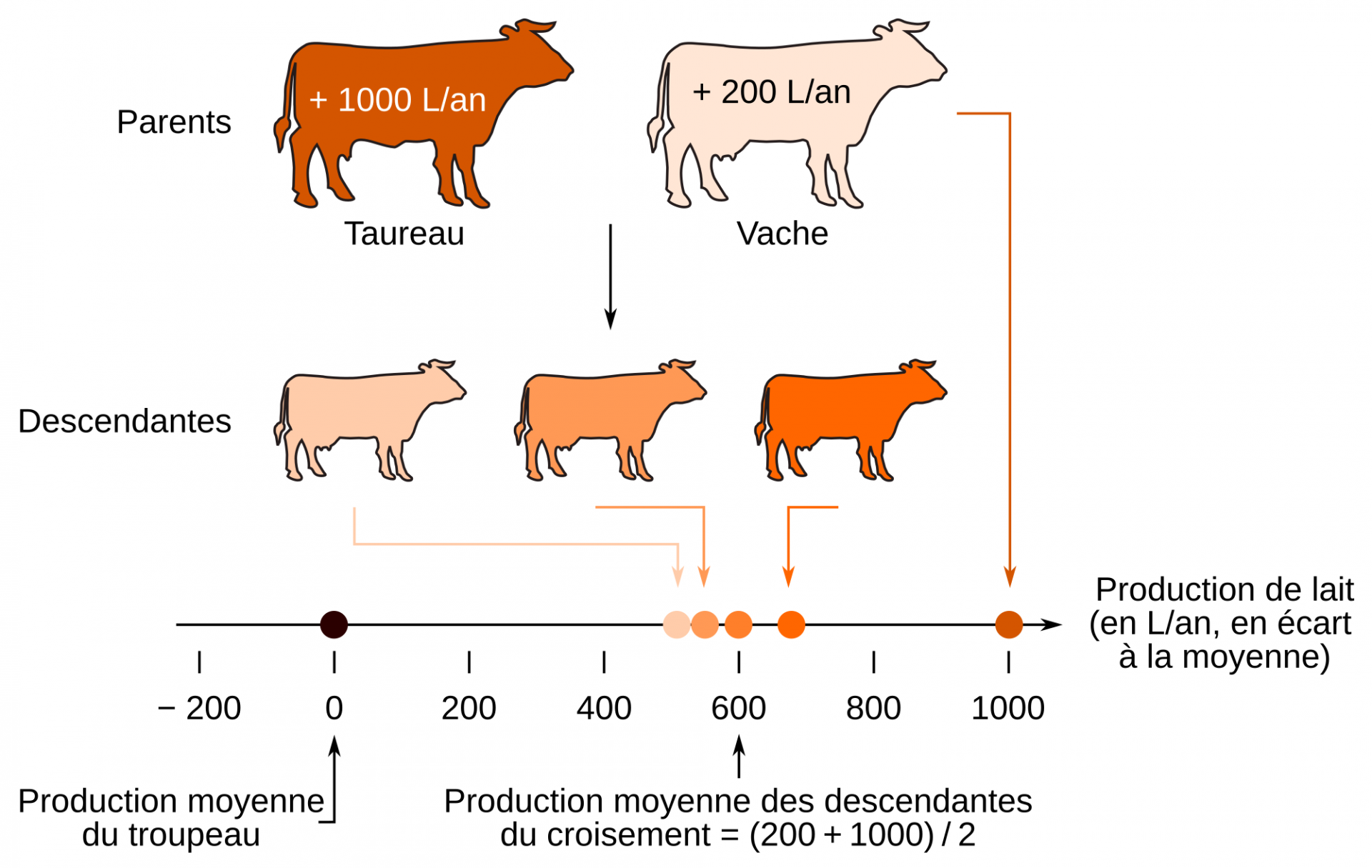
Valeurs génétiques estimées des parents et phénotypes attendus des descendants
Pour un croisement entre un mâle et une femelle donnés, la valeur attendue du phénotype chez les descendants est égale à la moyenne des valeurs génétiques estimées des deux parents. Pour reprendre l’exemple du caractère « production de lait », les descendants d’un croisement entre un père de valeur génétique + 1 000 L/an et d’une mère + 200 L/an produiront en moyenne 600 L/an de plus que la moyenne du troupeau (Figure 3). Ainsi, la valeur génétique représente la partie de la performance de l’animal qui est héréditaire et qui sera donc transmise à sa descendance.
L’amélioration génétique du troupeau
Plusieurs facteurs contraignent le rythme d’amélioration d’un caractère quantitatif dans un troupeau. Ainsi, le progrès génétique $\Delta G$, également appelé réponse à la sélection, dépend :
-
de l’intensité de sélection $i$. Celle-ci dépend de la proportion d’animaux sélectionnés pour produire la génération suivante. Plus cette proportion est faible (seuls les meilleurs animaux sont sélectionnés), plus l’intensité de sélection est élevée et donc plus le rythme d’amélioration du caractère est important. En élevage bovin, l’intensité de la sélection peut être très importante sur les taureaux (un seul taureau peut avoir des dizaines ou des centaines de descendants) mais est bien plus faible sur les vaches (qui n’ont en moyenne que trois lactations, donc trois descendants).
-
de l’écart type génétique $σ_{g} = \sqrt{h^{2}} σ_{p}$ avec $σ_{p}$ l’écart-type phénotypique qui est une mesure de la variabilité des performances. Plus la variabilité entre les animaux du troupeau est grande pour le caractère d’intérêt, et plus il est possible de sélectionner des animaux présentant des valeurs extrêmes pour le caractère d’intérêt.
-
de $R$, c’est-à-dire de la racine carrée de $R^{2}$, qui correspond à la corrélation entre la valeur génétique estimée (appelée index) et la valeur génétique « vraie » de l’animal.
-
de l’intervalle entre générations, noté $T$. Plus celui-ci est court, et plus le rythme d’amélioration du troupeau pourra être élevé. En élevage bovin laitier, l’intervalle entre générations est long, en particulier à cause du faible potentiel reproductif des vaches et du testage sur descendance des taureaux.
Au final, la réponse annuelle à la sélection est donnée par :
$$\Delta G = \frac{i σ_{g} R}{T} = \frac{ i \sqrt{h^{2}} σ_{p} R}{T}$$
La connaissance de ces paramètres permet aux éleveurs et aux éleveuses de prévoir l’évolution des performances de leur troupeau. En effet, si $σ_{g}$ est une constante du caractère, les paramètres $i$, $R$ et $T$ varient en fonction des méthodes de sélection.
Pour aller plus loin
Les éditions Quae proposent gratuitement au téléchargement le livre L'amélioration génétique animale de Gerald Wiener et Roger Rouvier, 2009 (EAN13 : 9782759203710).





_crossing.svg?uselang=fr){kind=link}