L'amélioration génétique des animaux de rente peut se faire en race pure ou bien par croisement. Cet article présente des résultats d’études sur le croisement entre races en bovins lait, espèce pour laquelle le croisement est encore très peu utilisé.
« La sélection animale consiste à favoriser la reproduction d’animaux qui possèdent des attributs supérieurs de façon à en propager les qualités au plus grand nombre. »
La sélection animale consiste à identifier les meilleurs reproducteurs et à diffuser leurs allèles dans la population dans un but d’amélioration génétique. Cette diffusion peut se faire en race pure, le plus souvent pour des espèces peu prolifiques et à cycle long (bovins lait, bovins viande, caprins et ovins viande). Elle peut également avoir lieu par croisement, plutôt pour des espèces prolifiques à cycle court et à vocation de produire de la viande (porcs, volailles, lapins et ovins viande). Après avoir défini la notion de race et donné les principes de la sélection génétique intrarace en bovins lait, cet article présente des résultats d’études sur le croisement entre races en bovins lait, espèce pour laquelle le croisement est encore très peu utilisé.
Notions de races en productions animales
La notion de race est assez récente et correspond à une subdivision de l’espèce (ou d’une sous-espèce), regroupant des animaux qui présentent des caractéristiques héréditaires communes. Jusqu’au milieu du XVIIIe siècle, les éleveurs ont constitué leurs troupeaux par croisement et brassage via l’importation d’animaux ayant les caractéristiques souhaitées. Puis, à partir du milieu du XIXe siècle, les éleveurs ont orienté les populations animales vers un idéal, la race pure, en constituant des populations locales dites fermées, c’est-à-dire non croisées avec des individus extérieurs. Cette notion de race se définissait par un standard correspondant principalement à des caractéristiques visuelles à déterminisme génétique simple (couleur, cornage, etc.). À la fin du XIXe et au début du XXe siècle, apparaissent les livres généalogiques par races (herdbooks). Dans la première moitié du XXe siècle se met en place les contrôles de performances (suivi des généalogies, de la production laitière…) qui vont fournir une base objective permettant une sélection rationnelle. Après la Seconde Guerre mondiale, avec la modernisation et l’intensification de l’agriculture, les races deviennent de plus en plus spécialisées. En France, on voit donc émerger quelques races dominantes qui sont particulièrement bien adaptées au marché. À partir de 1980, la sélection génétique s’intensifie, notamment grâce à la maîtrise des méthodes de reproduction artificielles et d’évaluations génétiques de plus en plus complexes permises par le progrès du numérique et par la maîtrise des outils de biologie moléculaire. À l’heure actuelle, en élevage bovin laitier, la race Prim’Holstein est la première race française en effectif et produit en moyenne 9 241 kg de lait par lactation (Institut de l’élevage et France conseil élevage, 2017).
Principe de la sélection génétique intrarace
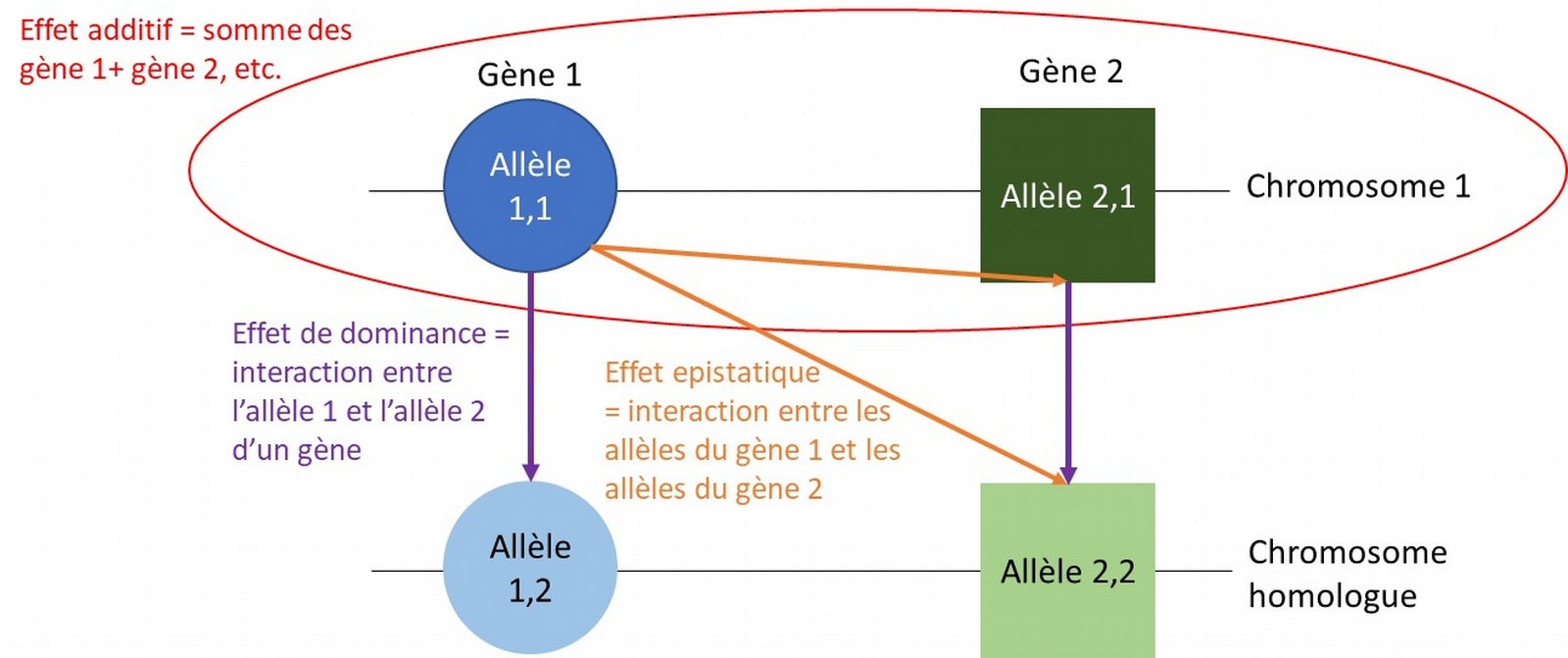
L’amélioration génétique repose sur deux grands types d’effets génétiques : les effets additifs et les effets d’interaction. La figure 1 illustre ces différents effets génétiques. Un individu ne transmet que la moitié de son patrimoine génétique et donc qu’un chromosome sur les deux chromosomes homologues de chaque paire. La somme des effets des gènes transmis à la descendance s’appelle la valeur génétique additive d’un individu. En revanche, les effets d’interaction sont propres à chaque individu car ils dépendent des allèles transmis par les deux parents. Ces effets d’interaction peuvent être positifs et augmenter la performance de l’individu ou négatifs et pénaliser la performance de l’individu. En sélectionnant les meilleurs individus en race pure, l’espèce humaine a sélectionné les meilleurs effets additifs mais également sélectionné les interactions génétiques favorables à l’expression de la performance.
En productions animales, les caractères d’intérêt pour la sélection génétique sont des caractères quantitatifs polygéniques. C’est-à-dire qu’ils sont mesurables chez les animaux qui les expriment (c’est le cas par exemple de la quantité de lait produite par la vache, de sa capacité à être gestante après une insémination, etc.) et qu’ils sont déterminés par un très grand nombre de gènes (probablement des milliers). L’expression de tels caractères est également influencée par l’environnement et notamment par le milieu d’élevage. Mesuré sur un très grand nombre d’individus, on constate que la variation d’un caractère quantitatif est continue et suit généralement une loi normale.
Si l’on considère un caractère quantitatif polygénique chez un individu en particulier, on peut, pour cet individu, déterminer la différence entre sa performance et la performance moyenne de la population à laquelle il appartient. Cette différence s’explique par des différences de valeur génétique et par des différences de milieu d’élevage. À partir de modèles mathématiques, on peut estimer les effets du milieu et la valeur génétique additive de l’individu. Plus l’effet du milieu sur l’expression du caractère est important moins la différence de performance s’explique par une différence de valeur génétique. On définit ainsi un paramètre génétique important appelé héritabilité. L’héritabilité, notée h2, a été définie pour décrire ce qui est transmissible intrarace dans la variabilité d’une performance ou d’un phénomène biologique. L’héritabilité correspond donc à la fraction de la variance phénotypique qui s’explique par une variation d’origine génétique. Plus un caractère est héritable, plus la probabilité qu’un individu avec des performances supérieures à la moyenne de la population transmette une valeur génétique additive positive est importante. L’héritabilité est donc un paramètre clé de l’amélioration génétique, les caractères fortement héritables étant plus facilement améliorables que les caractères faiblement héritables.
En bovin laitier, les caractères les plus héritables sont les taux de matière utile (taux butyreux et taux protéique). La quantité de lait a une héritabilité moyenne tandis que la fertilité et la santé ont des héritabilités très faibles. De plus, il existe entre les caractères d’intérêt des corrélations génétiques qui peuvent être soit favorables (c’est-à-dire qu’en ne sélectionnant qu’un des deux caractères, l’autre caractère va quand même être amélioré) ou défavorables (c’est-à-dire qu’en ne sélectionnant qu’un des deux caractères, l’autre caractère va subir une contre-sélection). En bovin laitier, c’est le cas notamment de la quantité de lait et de la fertilité qui sont des caractères corrélés génétiquement négativement. Sélectionner les meilleurs reproducteurs sur la quantité de lait entraîne donc une diminution de la fertilité dans la race. Par conséquent, les sélectionneurs doivent définir des objectifs de sélection qui déterminent les caractères d’intérêt et les compromis à faire pour les améliorer conjointement. Ces objectifs sont définis indépendamment pour chaque race.
En résumé, l’amélioration génétique doit prendre en compte trois paramètres : la variabilité génétique, c’est-à-dire qu’il existe des individus plus performants que d’autres, et que cet écart est en partie d’origine génétique ; l’héritabilité du caractère, c’est-à-dire la capacité de cet écart de performance à se transmettre à la descendance ; et les corrélations génétiques entre les caractères pour éviter de détériorer certains caractères d’intérêt. L’exploitation de la variabilité génétique additive par la sélection a conduit à sa réduction au cours du temps et à l’augmentation de la consanguinité au sein d’une même race pure puisqu’un nombre réduit de taureaux transmettent largement leurs allèles dans leurs populations raciales (Danchin et coll., 2012). Cette augmentation de la consanguinité, si elle est non maîtrisée, entraîne une diminution des performances de reproduction et augmente la probabilité d’apparition de tares génétiques.
Valeur génétique additive et variabilité génétique
Il ne faut pas confondre valeur génétique additive et variabilité génétique. La valeur génétique additive est la valeur d’un individu en comparaison à une population de référence, c’est une variable individuelle. La variabilité génétique est une variable de population. Elle se traduit généralement par un écart-type génétique autour de la moyenne des valeurs génétiques additives de la population.
Des différences d’orientation génétique entre races bovines laitières
En France, trois grandes races se détachent dans la production bovine laitière (Institut de l’élevage et Confédération nationale de l’élevage, 2018) : la Prim’Holstein (64 % des effectifs), la Montbéliarde (17 %) et la Normande (8,5 %). Dans ces trois races, une sélection sur la production laitière produite est réalisée, ce qui se traduit par une augmentation continue de la quantité de lait produite par vache et par lactation (Figure 2).
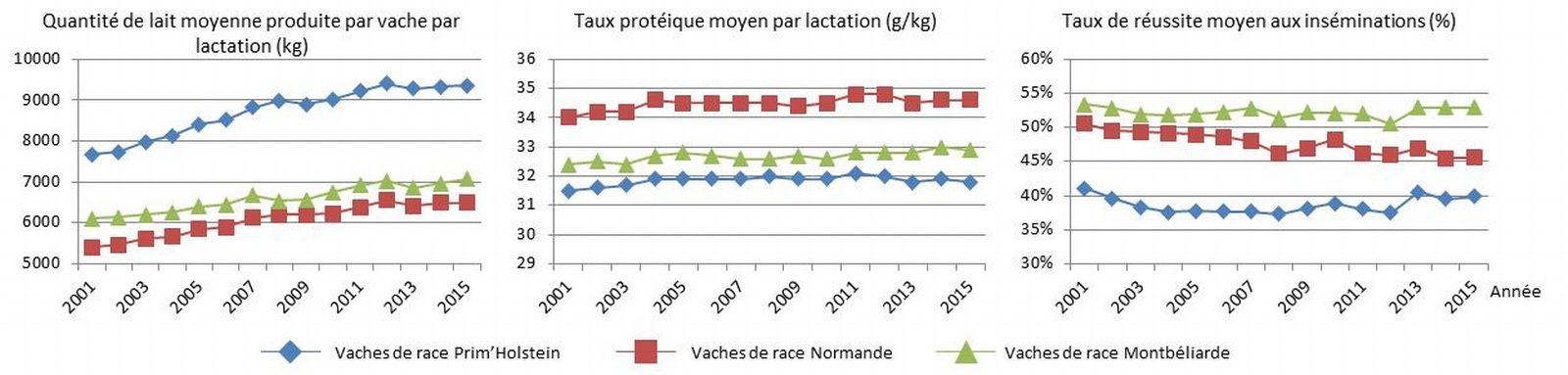
Sources : Le Mézec, P, & Launay, A. (2017) et Le Mezec, P., Barbat-Leterrier, A., & Barbier, S. (2017).
Cependant, les vaches de race Prim’Holstein se distinguent par une productivité laitière très supérieure à celle des vaches des deux autres races. En revanche, les vaches de race Normande produisent un lait dont le taux protéique est supérieur aux deux autres races tandis que les vaches de race Montbéliarde sont plus fertiles. Ces écarts de performances traduisent en partie les corrélations négatives existant entre ces différents caractères. En effet, la quantité de lait est génétiquement corrélée négativement avec le taux protéique et la fertilité. De plus la fertilité est génétiquement corrélée négativement avec le taux protéique (V. Ducrocq et D. Boichard, Inra de Jouy-en-Josas ; communication personnelle). Sélectionner conjointement ces caractères suppose donc de faire des compromis sur l’amélioration génétique de chaque caractère pris indépendamment. Les orientations génétiques décidées par les organismes de sélection de chacune des races ont été différentes dans le temps (Institut de l’élevage, 2012), ce qui conduit aux écarts de performances observables en Figure 2. Il apparaît ainsi difficile d’obtenir en race pure des améliorations sur la quantité de lait, la qualité du lait et la fertilité. En revanche en croisant ces différentes races, on pourrait a priori bénéficier des efforts de sélection réalisés dans chacune des races impliquées. Les parties suivantes ont pour objectif d’illustrer et de quantifier cet intérêt.
Réaliser des croisements entre races bovines laitières c’est bénéficier de la complémentarité entre races
Tout comme en race pure, le principe génétique du croisement repose sur les effets génétiques additifs et les effets d’interaction. La partie génétique de la performance d’un animal issu d’un croisement dépend de trois composantes distinctes : la contribution additive moyenne de chacune des races constitutives ; la valeur génétique additive individuelle intrarace de chacun des deux parents ; l’effet spécifique du croisement. Prédire la valeur d’un animal revient donc à prédire ces trois composantes. Il convient de noter que si les valeurs additives raciales et individuelles se transmettent des parents aux descendants de façon simple, l’effet spécifique du croisement dépend la composition raciale des parents. Les effets génétiques du croisement vont donc dépendre de la valeur intrinsèque de chaque parent et du type de croisement réalisé.
Dans notre étude (Dezetter et coll., 2015), à partir des données du contrôle de performance des vaches de races pures et issues de croisement, nous avons estimé les paramètres génétiques du croisement entre les trois grandes races laitières française (Prim’Holstein, Montbéliarde et Normande). Nous avons montré que la race Prim’Holstein est génétiquement supérieure pour le caractère de quantité de lait aux races Montbéliarde et Normande (Figure 3). Ces races sont quant à elles génétiquement supérieures pour les caractères de fertilité. Ces écarts entre races laissent présager d’une bonne complémentarité entre celles-ci. En effet, si on prend l’exemple des caractères quantité de lait et taux de conception, nous avons estimé un écart génétique additif de 1150 kg de lait sur 305 jours en équivalent adulte et un écart génétique additif de − 12 points de pourcentage de réussite à l’insémination entre les races Prim’Holstein et Montbéliarde. En prenant pour référence la Montbéliarde, la valeur génétique additive moyenne des vaches F1 issues Montbéliarde × Prim’Holstein est donc de 575 kg de lait sur 305 jours en équivalent adulte et de − 6 points de taux de conception. Les vaches F1 maintiennent donc un niveau de production de lait supérieur à la Montbéliarde tout en ayant un potentiel génétique de reproduction nettement supérieur à la Prim’Holstein.
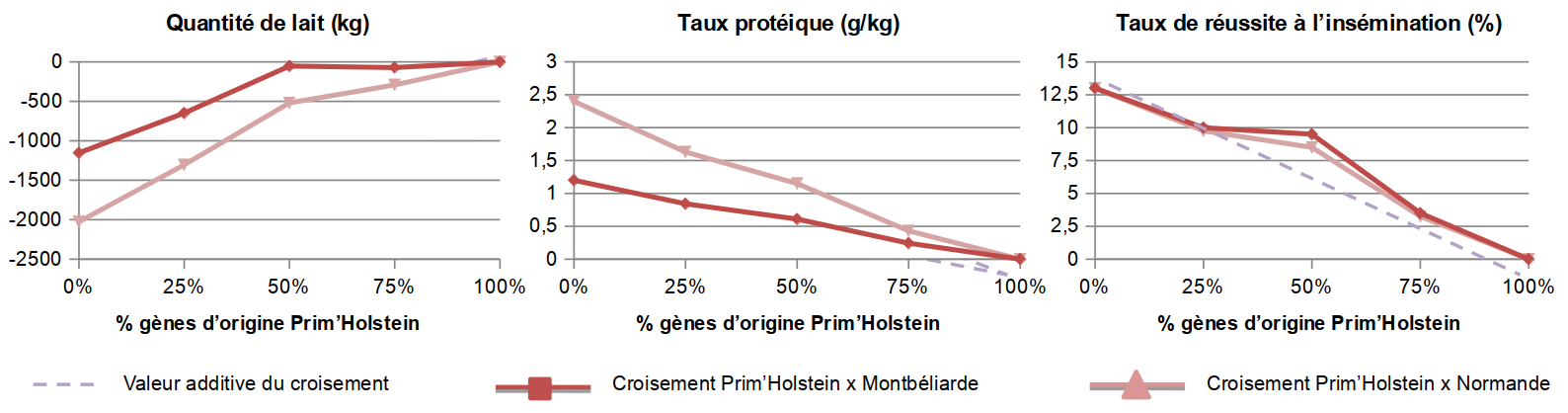
On mesure la quantité de lait produite sur 305 jours, le taux protéique du lait et le taux de réussite à l’insémination chez différents types vaches, par rapport aux vaches Prim’Holstein prises comme référence (valeur 0). Source : Dezetter et coll., 2015.
Expression théorique des effets génétique non additif chez les individus croisés
On définit l’effet d’hétérosis, ou de vigueur hybride, comme la différence entre la performance moyenne de la population issue du croisement de première génération et la performance moyenne des deux populations parentales (Shull 1948, Bidanel 1992). Cet écart de performances, souvent favorable, s’explique essentiellement par des effets non additifs de dominance et/ou d’épistasie. Les effets de dominance sont générés par des interactions génétiques intraloci (c’est-à-dire entre les deux allèles d’un même gène), tandis que les effets épistatiques sont causés par des interactions entre loci (entre gènes différents). De manière générale, les effets de dominance représentent la part la plus importante des interactions. En race pure, la sélection génétique, en se basant sur les effets additifs (et donc héritables) et en augmentant la fréquence de certains allèles dans la race, augmente le taux d’homozygotie. Cela réduit donc les effets de dominance dans l’expression des caractères. En revanche, le croisement entre races augmente l’hétérozygotie et donc l’importance des effets de dominance dans l’expression des caractères. En règle générale cet effet est favorable et s’ajoute aux effets additifs, c’est ce qu’on appelle l’effet d’hétérosis.
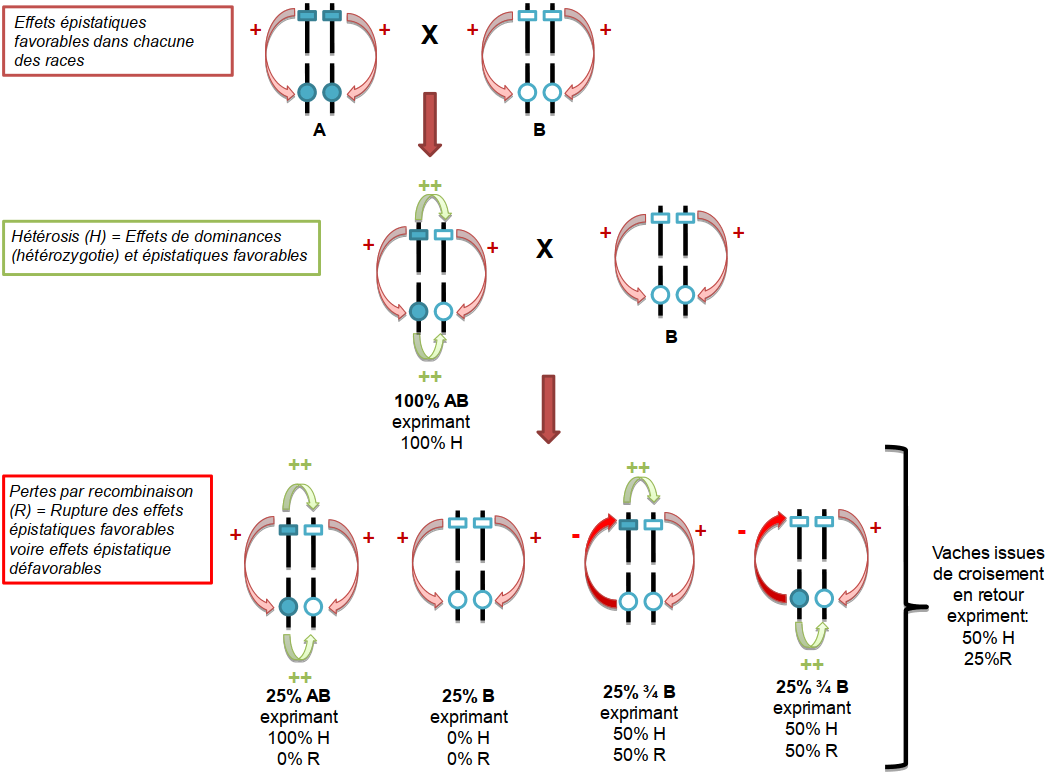
On considère l’exemple de deux parents issus de deux races homozygotes au niveau de deux loci indépendants. Aucun des deux parents n’exprime d’effet de dominance (puisqu’ils sont homozygotes) au niveau de ces deux loci. En revanche ils expriment des effets épistatiques favorables issus de la sélection (flèche rouge). Lorsque ces deux parents sont accouplés, les vaches F1 issues de ce croisement vont exprimer un effet d’hétérosis maximal. En effet, les vaches F1 possèdent les deux copies de chaque locus et donc expriment 100 % des effets de dominance (flèches vertes). Puis ces vaches F1 sont accouplées avec un taureau de la race B (c’est ce qu’on appelle un croisement en retour). En moyenne, les vaches issues de cet accouplement n’expriment plus que 50 % de l’effet d’hétérosis observé chez les vaches F1 (4 flèches vertes sur 8 interactions de dominance possible), et vont exprimer 25 % de pertes par recombinaison suite à la rupture d’interactions favorables entre les deux loci (flèche rouge accompagnée d’un −).
Parce que les races pures ont été sélectionnées pendant un temps long, elles ont accumulé et maintenu des interactions favorables entre gènes. Les effets épistatiques en race pure sont donc favorables à l’expression du caractère dans le milieu de sélection. Comme il s’agit d’une composante raciale, les effets épistatiques détruits lors de la recombinaison méiotique sont immédiatement reconstitués intrarace. Au contraire, lors d’un accouplement impliquant un individu croisé, ces interactions positives détruites par la recombinaison méiotique ne sont que partiellement reconstituées (uniquement entre gènes reçus de la même race). Cette perte, dite effet de recombinaison épistatique, se traduit par une diminution de la performance observée par rapport à la performance attendue en ne tenant compte que des effets additifs et de dominance.
Race pure et lignée pure
En production animale, la notion de race pure désigne la situation où les reproducteurs s’unissent au sein d’une même population. Elle s’oppose au croisement, où les reproducteurs viennent de populations différentes de la même espèce. En race pure, le taux d’homozygotie est plus élevé qu’en croisement, mais il n’est jamais de 100 %. La notion de race pure est donc à distinguer de celle de lignée pure, utilisé pour des animaux de laboratoire, où l’accouplement d’animaux consanguins permet d’obtenir à terme une homozygotie pour tous les locus.
Des effets d’hétérosis favorables et des pertes par recombinaison limitées chez les vaches laitières issues de croisement
En règle générale, l’effet d’hétérosis pour un caractère est favorable et d’autant plus important que son héritabilité est faible (Lynch et Walsh 1998). En effet, pour les caractères influencés par des gènes à effets essentiellement additifs (et donc très héritables), les effets de dominance sont généralement très faibles et donc l’effet d’hétérosis est quasi nul. C’est plutôt l’inverse pour les caractères faiblement héritables qui sont peu influencés par des gènes à fort effets additifs et dont l’expression est plutôt influencée par des effets d’interaction et des effets du milieu. L’effet d’hétérosis est également plus important lorsque les populations parentales sont génétiquement éloignées (Lynch et Walsh 1998). En effet, on vient de voir que plus un individu est hétérozygote plus l’effet d’hétérosis est important. Or plus les races sont éloignées génétiquement, plus la fréquence des allèles est différente au sein des deux races et donc plus l’individu issu du croisement entre ces deux races aura de chance d’être hétérozygote pour un grand nombre de gènes. Chez les bovins laitiers, on constate que les effets d’hétérosis sont responsables d’une augmentation de 5 à 6 % des caractères de production (quantités de lait, de matière grasse et de matière protéique) par rapport à la moyenne parentale, tandis qu’ils peuvent être de plus de 10 % pour les caractères fonctionnels tels que la fertilité.
Concernant les trois grandes races françaises, la figure 3 illustre l’impact des différents effets additifs et non-additifs sur le potentiel génétique des vaches. Ainsi les vaches Montbéliarde ont un potentiel génétique additif de − 1150 kg de lait par rapport aux vaches Prim’Holstein. C’est-à-dire que dans des conditions d’élevage exactement similaires, les vaches Montbéliarde produisent en moyenne 1150 kg de lait de moins que les vaches Prim’Holstein. On pourrait donc s’attendre à ce que les vaches F1 issues du croisement entre la race Prim’Holstein et la race Montbéliarde aient un potentiel génétique de − 575 kg de lait par rapport aux vaches Prim’Holstein. Or, en réalité, la différence de potentiel génétique entre les F1 et les vaches Prim’Holstein est d’environ − 55 kg. Donc l’hétérosis est « responsable » des 520 kg produits en plus par les F1 par rapport à ce que l’on pourrait attendre en ne regardant que le potentiel des parents. Pour le taux de matière protéique, les effets d’hétérosis sont très faibles, voire négligeables, et seul l’écart génétique entre les races explique le meilleur potentiel des vaches F1 issues de croisement par rapport aux vaches Prim’Holstein. Concernant les caractères de fertilité, en combinant effet additif et effet d’hétérosis, les vaches F1 issues des croisements entre Prim’Holstein et Montbéliarde ou Prim’Holstein et Normande expriment un potentiel supérieur aux vaches Prim’Holstein d’environ + 10 points de taux de conception.
Les pertes par recombinaison sur les animaux de deuxième génération de croisement sont généralement défavorables. Cumulées avec la perte progressive de l’expression des effets d’hétérosis, ces pertes viennent réduire la supériorité des individus issus de croisement. Dans notre étude nous nous sommes intéressés aux individus de deuxième génération issus d’un croisement par retour : vaches nées d’une mère F1 Prim’Holstein × Montbéliarde (ou Normande) et d’un père Prim’Holstein, donc ¾ Prim’Holstein, et vaches nées d’une mère F1 Prim’Holstein × Montbéliarde (ou Normande) et d’un père Montbéliard (ou Normand), donc ¾ Montbéliarde (ou ¾ Normande). Nous avons estimé que ces individus F2 exprimaient un effet de pertes par recombinaison allant − 32 kg de lait pour le croisement avec la Normande à − 45 kg de lait pour le croisement avec la Montbéliarde.
Par conséquent, pour le lait, les individus [¾ Prim’Holstein, ¼ Montbéliarde] ont un potentiel génétique de − 72,5 kg par rapport aux vaches Prim’Holstein (0,25 × (− 1150) kg (soit ¼ de la différence génétique entre les vaches Prim’Holstein et les vaches Montbéliardes) + 0,5 × 520 kg (car elles expriment 50 % de l’effet d’hétérosis) – 45 kg de pertes par recombinaison).
Les individus [¾ Montbéliarde, ¼ Prim’Holstein] ont quant à eux un potentiel génétique de − 647,5 kg par rapport aux vaches Prim’Holtsein (0,75 × (− 1150) kg + 0,5 × 520 kg − 45 kg). La figure 3 illustre l’impact de ces pertes par recombinaison sur le potentiel génétique de ces individus puisque l’on observe une cassure au niveau des animaux ¾.
En raison du nombre réduit d’individus issus de deuxième génération de croisement dans la population analysée, ces estimations sont statistiquement non significatives. Il faut cependant garder en tête qu’elles existent et qu’elles contribuent sans doute à expliquer pourquoi les performances des individus issus de croisement de deuxième génération et plus sont inférieures à celles espérées.
Une étude plus prospective sur l’impact économique d’introduire du croisement dans un troupeau pour bénéficier de ces effets génétiques a été menée (Dezetter, 2015 ; Dezetter et coll., 2017). Les principaux résultats montrent que dans des troupeaux de vaches Prim’Holstein à très haut niveau de production laitière et sans troubles majeurs de la reproduction ni de la santé, inséminer ces vaches avec des taureaux Montbéliard ou Normand (puis ces vaches F1 avec un taureau Prim’Holstein et ainsi de suite) pour constituer un troupeau de vaches issues de croisement présente peu d’intérêt. En effet, les vaches issues de croisement n’arrivent pas à atteindre le même niveau de production laitière et donc le volume commercialisé diminue fortement, pénalisant le produit de l’exploitation. En revanche sur des troupeaux de vaches Prim’Holstein à plus faible niveau de production laitière ou avec des troubles majeurs de la reproduction et de la santé, inséminer ses vaches en croisement augmente le produit de l’exploitation (en augmentant les taux de matière utile qui sont une composante du prix du lait) et en diminuant les charges de l’exploitation (car le croisement contribue à améliorer la reproduction et la santé des vaches). Il faut cependant noter que le croisement augmente la variabilité des phénotypes (car augmente l’hétérozygotie) et complexifie le suivi des accouplements car dans un troupeau sont présentes à la fois les mères, les filles et les petites filles, donc des vaches de générations de croisement différentes.
Références
- Bidanel, J. P. 1992. Comment exploiter la variabilité génétique entre races : du croisement simple à la souche synthétique. INRA Productions animales, (hs), 249-254.
- Danchin‐Burge, C., Leroy, G., Brochard, M., Moureaux, S., Verrier, E. 2012. Evolution of the genetic variability of eight French dairy cattle breeds assessed by pedigree analysis. J. Anim. Breed. Genet., 129, 206–217.
- Dezetter C., 2015. Evaluation de l’intérêt du croisement entre races bovines laitières. Thèse de l’école doctorale Biologie-santé Nantes Angers, LUNAM, France. Accessible sur : http://www.theses.fr/s163135.
- Dezetter C., Bareille N., Billon D., Côrtes C., Lechartier C., Seegers H., 2017. Changes in animal performance and profitability of Holstein dairy operations after introduction of crossbreeding with Montbéliarde, Normande, and Scandinavian Red. J. Dairy Sci., 100, 8239-8264.
- Dezetter, C., Leclerc, H., Mattalia, S., Barbat, A., Boichard, D., et Ducrocq, V. 2015. Inbreeding and crossbreeding parameters for production and fertility traits in Holstein, Montbéliarde, and Normande cows. J. Dairy Sci., 98, 4904-4913.
- Institut de l’Elevage. 2012. IBL 2012-2 ISU 2012 : les objectifs de sélection évoluent. http://idele.fr/no_cache/recherche/publication/idelesolr/recommends/ibl-2012-2-isu-2012-les-objecti…
- Institut de l’Elevage et Confédération Nationale de l’Elevage. 2018. Les chiffres clés du GEB, bovins 2018, productions lait et viande.
- Institut de l’Elevage et France Conseil Elevage. 2017. Résultats de contrôle laitier des espèces bovine, caprine et ovine, France 2017. Compte rendu n° 0018201022.
- Le Mezec, P., Barbat-Leterrier, A., et Barbier, S. 2017. Fertilité des vaches laitières : c’est mieux. Institut de l’Elevage, Technipel n°L0070.
- Le Mézec, P, et Launay, A. 2017. Le cheptel laitier français : évolution 1996-2016, prévision d’évolution génétique 2016-2022. Institut de l’Élevage, Compte-rendu n° 0017203001.
- Lynch, M., Walsh, B. 1998. Genetics and Analysis of Quantitative Traits. Sinauer Associates, Sunderland, MA.
- Minvielle, F. 1998. La sélection animale. Collection Que sais-je ?. Edition Presses Universitaires de France – PUF.
- Shull, G. H. 1948. What is » heterosis »?. Genetics, 33, 439.