Lien entre la taille de certains Vertébrés et les contraintes mécaniques pesant sur leur système musculo-squelettique. Description de différentes solutions permettant de réduire ces contraintes.
Cet article est la traduction de l'article Where Have All the Giants Gone? How Animals Deal with the Problem of Size paru dans la revue PLOS One le 11 janvier 2017 et publié sous licence Creative Commons - Paternité. Traduction réalisée par Pascal Combemorel.
La survie des prédateurs et des proies est souvent une question de vitesse. Cependant, la vitesse à laquelle peut courir un animal est intimement liée à sa taille, de sorte que les animaux les plus rapides ne sont ni les plus gros, ni les plus petits. La capacité à maintenir une vitesse élevée dépend de l’aptitude du corps à supporter les importantes contraintes engendrées par la locomotion. Même en restant immobile, les principes de mise à l’échelle suggèrent que le stress mécanique ressentit par un animal augmente plus rapidement que ce que son corps ne peut supporter. Si les gros animaux veulent être rapides, ils doivent donc trouver une solution pour surmonter ces stress élevés. Cet article explore les manières dont les animaux atténuent les contraintes musculo-squelettiques liées à leur taille, afin d’essayer de comprendre où sont passés les animaux géants.
Introduction
Les plus gros animaux à avoir jamais marché sur Terre ont disparu depuis longtemps. Les fossiles montrent que les plus gros de ces animaux, les dinosaures, atteignaient des masses de plus de 50 000 kg. Compte tenu de leur masse énorme, cela reste un mystère de savoir comment ils pouvaient marcher. Seules les espèces actuelles nous renseignent sur la façon dont la locomotion des animaux évolue avec leur taille. Être gros a des avantages : moins de prédateurs, plus de réserves disponibles, une meilleure rétention de la chaleur et une capacité à voyager loin étant donné le faible coût relatif du transport. Cependant, il y a un désavantage majeur à être grand. La force de gravité qu’un animal subit est proportionnelle à sa masse. Quand celle-ci s’accroît, l’intensité des forces auxquelles son squelette doit résister s’amplifie aussi. Cela ne serait pas un problème si la capacité d’un animal à supporter cette masse augmentait proportionnellement, mais ce n’est pas le cas. De plus, les performances d’un animal, comme la vitesse à laquelle il peut courir, sont liées à cet équilibre entre la taille de son corps et la capacité réduite à supporter ce poids.
Être grand a des avantages, mais être rapide aussi. Pour palier le problème de la taille, différents groupes d’animaux ont adopté diverses stratégies permettant de réduire les contraintes musculo-squelettiques. Cependant, le type de solution retenue peut influencer la relation entre la vitesse d’un animal et sa taille, limitant la vitesse maximale atteinte par l’animal. Être gros et lent est potentiellement un désavantage et pourrait avoir contribué à la chute des animaux géants, maintenant disparus. Mais étaient-ils vraiment grands et lents ? Pour comprendre les solutions permettant de réduire les contraintes mécaniques, et les limitations de vitesse liées à la taille, nous devons d’abord comprendre ce qui cause l’augmentation des contraintes quand les animaux deviennent grands.

Les lignes grises sont des droites de régression montrant une mise à l’échelle géométrique (isométrie) pour la longueur, l’aire et le volume. La ligne bleue, en pointillés, montre que l’aire de section de l’iliofibulaire (muscle) subit une mise à l’échelle à allométrie positive. Chez les Varanidés. Chez les Mammifères, la longueur du fémur se met à l’échelle avec une allométrie négative (ligne jaune en pointillés)
Si les animaux grandissaient en taille mais gardaient la même forme, autrement dit s’ils subissaient une mise à l’échelle géométrique (isométrie), leurs longueurs augmenteraient proportionnellement à M0,33 (avec M la masse de l’animal) tandis que leurs surfaces augmenteraient proportionnellement à M0,67 (Figure 1). Une contrainte est une force exercée sur une certaine surface (σ = F/A) et, comme la force augmente d’un facteur M1 alors que la surface n’augmente que d’un facteur M0,67, une mise à l’échelle géométrique provoquerait une augmentation des contraintes pesant sur les os et les muscles d’un facteur M0,33 (M1/M0,67). Quand la taille des animaux s’accroît, il y a donc une augmentation des contraintes : les changements de dimension des membres ne suffisent pas à compenser l’augmentation de la charge mécanique. Cependant, il existe quand même de grands animaux dans la nature, ce qui amène à se demander comment ils s’en sortent avec l’augmentation des contraintes liées à leur taille. Est-ce que ces animaux fonctionnent simplement dangereusement près du point de rupture de leurs os et de leurs muscles ? Les études réalisées sur un grand nombre d’animaux suggèrent que c’est peu probable. Des animaux allant de 0,04 à 300 kg maintiennent des seuils de sécurité similaires : les contraintes maximales durant leur locomotion sont deux à dix fois inférieures à leur seuil de rupture. Dans cet article, nous décrivons cinq solutions possibles grâce auxquelles les animaux peuvent éviter les problèmes mécaniques liés à la taille, ce qui permettra d’éclairer la façon dont les dinosaures pouvaient se déplacer. Les animaux peuvent devenir plus grands de différentes manières :
- en modifiant les propriétés mécaniques de leurs os et muscles ;
- en changeant la forme de leurs os ;
- en changeant la manière dont les charges sont réparties sur les os ;
- en modifiant l’architecture des muscles squelettiques ;
- en subissant une diminution de leurs performances locomotrices, c’est-à-dire en ralentissant.
Les os ou les muscles deviennent plus résistants
Les gros animaux pourraient régler les problèmes d’augmentation des contraintes grâce à des os ou à des muscles plus résistants. Les mesures des contraintes pour lesquelles les os cassent ne montrent pas de différences dans le seuil de rupture de ceux-ci, pour des animaux allant de 0,04 à 700 kg. Tout comme celles des os, les propriétés des muscles squelettiques sont hautement conservées. En fait, l’arrangement des protéines d’actine et de myosine des muscles ne varie pas selon les taxons. De plus, Seow et Ford ont mesuré les propriétés mécaniques des muscles squelettiques sur une large gamme de taille et n’ont pas trouvé de différence dans le diamètre des fibres ou dans la force de contraction isométrique maximale, suggérant que la contrainte maximale à laquelle les muscles peuvent résister est la même chez les petits et les grands animaux. Cependant, une modélisation récente laisse penser que l’architecture des os trabéculaires des Mammifères et Oiseaux allant de 0,003 à 3,400 kg varierait avec la taille du corps. Cela pourrait réduire la pression sur les os au niveau cellulaire, mais il reste à savoir si cela diminuerait les contraintes à l’échelle du squelette. D’autres solutions peuvent donc être nécessaires.
Les os deviennent plus épais
Un autre mécanisme pouvant régler le problème de l’augmentation des contraintes est un changement dans la forme des os. Des os plus épais ou des membres plus courts réduiraient les contraintes pesant sur les os. McMahon a proposé un modèle de mise à l’échelle appelé « similarité élastique » dans lequel les animaux optimisent la forme de leurs os pour devenir plus robuste et plus à même de faire face à l’augmentation des contraintes. Ce modèle s’accorde bien avec ce que l’on observe pour les os des membres des ongulés et pour l’humérus des reptiles et des Mammifères, mais d’autres études montrent que lorsque l’on considère un vaste panel de Mammifères, la dimension des membres augmente de façon presque isométrique. Des études ont également estimé la mise à l’échelle des os dans des taxons éteints et ont révélé des similarités entre les dinosaures et les Mammifères, avec des os devenant légèrement plus épais dans les espèces de grande taille, comparativement à celles de petite taille. L’augmentation de la taille des membres montre donc d’importantes variations : il n’y a pas de règle générale permettant de décrire tous les Vertébrés terrestres.
Cependant, même si les os suivaient la théorie élastique de McMahon, cela ne serait peut-être pas suffisant. Biewener a montré que même avec des os légèrement plus épais, les stress maximaux augmenteraient quand même chez les Mammifères d’un facteur M0,28. D’autres groupes d’animaux montrent des caractéristiques semblables. Parmi les Varanidés par exemple, le diamètre des os des membres augmente avec une allométrie positive (NdT : cela signifie que la croissance des os des membres est comparativement plus importante que la croissance globale de l’organisme) et, de même que chez les Mammifères, les comparaisons avec les modèles théoriques montrent que ces changements dans la géométrie des os des membres ne suffisent pas à compenser l’augmentation des contraintes pesant sur ceux-ci. Il apparaît donc que les changements de propriétés des os, ou ceux de la forme des os, ne peuvent à eux seuls compenser l’augmentation des contraintes. […]
Les changements de posture
Depuis des siècles, les scientifiques ont remarqué que les petits Mammifères se déplacent en ayant une posture accroupie dans laquelle leurs membres sont fléchis sous leurs corps, alors que les animaux plus gros se déplacent en se tenant droits, avec les membres en extension. En se tenant droits, les gros animaux augmentent l’avantage mécanique efficace (AME, ou EMA en anglais, effective mechanical advantage) de leurs articulations et diminuent donc fortement les stress musculo-squelettiques durant leurs déplacements (Figure 2). L’AME est proportionnel à M0,25, ce qui correspond à l’augmentation des contraintes qui se produirait si la posture ne changeait pas (Figure 2). Cependant, il y a probablement un maximum à l’AME : chez les gros animaux terrestres (d’une masse supérieure à 300 kg environ), l’AME n’augmente plus linéairement avec la masse de l’animal car les membres sont presque étendus au maximum. Par conséquent, la posture semble être une explication au fait que les contraintes pesant sur les os et les muscles restent inchangées chez des Mammifères et des Oiseaux phylogénétiquement éloignés, au moins pour les masses inférieures à 300 kg. Mais même dans cette gamme de petites tailles, cette stratégie de réduction des contraintes pourrait ne pas être employées par tous les taxons.

(A) L’avantage mécanique efficace (AME) des membres postérieurs des Mammifères augmente proportionnellement à M0,26. (B) Effet de la posture du membre postérieur sur l’AME (ratio entre le bras de levier du muscle extenseur (r) et le bras de levier de la force de réaction du sol (R)). L’AME du membre postérieur des animaux à posture accroupie est inférieur à celui des animaux se tenant droits. Les flèches pointillées représentent le vecteur force de la réaction du sol.
Les Félidés, ou Félins, forment un groupe d’animaux dans lequel la posture ne varie pas pour des masses allant de 4 à presque 200 kg (Figure 3). Plutôt que de se tenir droits, les Félidés maintiennent une posture accroupie, même pour des tailles importantes ; cette posture pourrait leur permettre d’accélérer et de changer de direction plus facilement ce qui, comme la vitesse, peut augmenter la valeur sélective. Les Varanidés sont un autre groupe dans lequel le lien entre la taille des animaux et leur posture est ténu. Ils ne se redressent pas pour des masses allant de 0,04 à 8 kg mais maintiennent au contraire une posture accroupie dans laquelle les fémurs sont en abduction (NdT : c’est-à-dire écartés du plan médian du corps), un peu comme s’ils faisaient des pompes. La flexion des membres chez les Félidés et leur abduction chez les Varanidés réduit l’AME s’opposant à la force de gravité. […] Cependant, certains Varanidés atteignent tout de même des masses importantes.

Les angles des articulations pour les Félidés (cheville : points rouges, ligne pointillée ; genou : points oranges, ligne continue) et pour les Varanidés (cheville : pojnts bleus foncés, ligne pointillé ; genou : points bleus clairs, ligne continue) montre que la posture varie peu avec la masse dans les deux groupes, et que les Varanidés ont une posture plus accroupie que les Félidés. Les angles des articulations des Félidés sont représentés dans le plan sagittal alors que pour les Varanidés, les angles sont montrés en trois dimensions.
Par exemple, le dragon de Komodo semble défier les relations existant entre la posture d’un animal et sa taille, avec sa masse dépassant 100 kg et sa posture très accroupie. Une simple mise à l’échelle géométrique prédirait que les contraintes pesant sur les muscles de ces lézards géants les amèneraient près de leur point de rupture, causant des fractures ou des déchirures lors d’efforts quotidiens. Cependant, les dragons de Komodo semblent bien s’en sortir, en atteignant des vitesses de 20 km.h-1 tout en gardant leur système musculo-squelettique intact. Comment des animaux aux postures accroupies comme les lézards et les Félidés arrivent-ils à gérer l’augmentation des contraintes liées à leur taille ? Un mécanisme possible pourrait être un changement dans l’architecture de leurs muscles squelettiques.
Les changements de l’architecture des muscles
L’architecture des muscles squelettiques des gros animaux pourrait être adaptée pour supporter l’augmentation de masse. Les muscles squelettiques servent à la fois au support et à la locomotion. Toutes choses étant égales par ailleurs, de plus gros muscles (en masse ou en surface) seraient capables de supporter et de produire des forces plus importantes. Cependant, l’arrangement des fibres dans un muscle affecte aussi son fonctionnement. Par exemples, les muscles pennés sont caractérisés par des fibres placées obliquement par rapport à l’axe du muscle. […] Cela permet (i) de grouper plus de fibres musculaires dans un volume donné, augmentant ainsi l’aire de section physiologique du muscle (NdT : l’aire de section physiologique (physiological cross-sectional area) correspond à l’aire de la section d’un muscle, perpendiculairement à la disposition des fibres musculaires) et (ii) aux fibres musculaires de pivoter avec un angle élevé durant la contraction, ce qui découple la vitesse de raccourcissement des fibres de la vitesse du mouvement. Pour de gros animaux, une augmentation de l’épaisseur relative des muscles permet de répartir les forces sur une plus grande surface et permet donc de diminuer les contraintes que subissent les muscles.
Certains groupes d’animaux altèrent les propriétés mécaniques de leurs muscles afin d’atténuer l’augmentation des contraintes liées à la taille. Les Mammifères allant de 0,009 à 2,5 kg augmentent la force générée par leurs muscles en augmentant leur masse ou leur section, ou en diminuant la longueur des fibres musculaires. Nos travaux sur les Varanidés, et ceux de Cuff et de ses collègues sur les Félidés, étaient parmi les premiers à montrer que l’architecture des muscles squelettiques variait avec la taille, pour les groupes dans lesquels la posture est indépendante de la taille du corps. Chez les lézards allant de 0,008 kg (Varanus brevicauda) à 40 kg (Varanus komodoensis), nous avons montré une augmentation de la force relative générée par les adducteurs du fémur, les fléchisseurs du genou et les fléchisseurs de la cheville, avec une allométrie positive pour la masse et la section de ces muscles et, dans une moindre mesure, pour l’angle des fibres des muscles pennés (Figure 1). L’absence de preuve solide de changements géométriques suggère que les gros animaux gardent la même vitesse de raccourcissement relatif et utilisent la masse musculaire supplémentaire pour augmenter la force générée par leurs muscles. Ce changement dans l’architecture des muscles squelettiques, qui offrent un plus grand soutien, peut expliquer l’importante taille des dragons de Komodo, qui maintiennent pourtant une posture accroupie.
Chez les Félidés, la situation est différente. Seulement l’un des 38 membres postérieurs étudiés se met à l’échelle avec une allométrie positive pour la masse musculaire, laissant penser que les grands Félidés ont tout simplement des membres postérieurs relativement faibles par rapport aux petits félins. Ces différences de propriétés des muscles chez les Varanidés et chez les Félidés pourrait être expliqué par les différences de posture entre ces deux groupes (Figure 3). Si l’on retrouve dans les deux cas une position accroupie, les membres des Félidés sont en position parasagittale (dans un plan parallèle au plan de symétrie bilatérale) alors que ceux des Varanidés sont en position transversale. Une comparaison des angles des articulations des membres postérieurs montre que les Varanidés sont plus accroupis […]. Pour surmonter les problèmes liés à la taille, les gros animaux à posture accroupie peuvent donc soit augmenter la taille de leurs muscles (masse ou surface) pour favoriser le soutien, comme chez les Varanidés, ou simplement faire avec des muscles comparativement plus faibles, comme chez les Félidés. Ces deux situations diminuent toutefois probablement les performances locomotrices.
Les vitesses diminuent
Il n’y a pas une stratégie universelle pour diminuer les contraintes liées à la taille. Différents groupes d’animaux utilisent différentes stratégies biomécaniques pour réduire les contraintes pesant sur leur système musculo-squelettique. Même dans des groupes proches, la capacité de ces techniques à réduire les contraintes dans une vaste gamme de taille pourrait être limitée. Une fois la limite atteinte, la seule façon de réduire efficacement les contraintes est de diminuer la vitesse de locomotion. En ralentissant, les animaux peuvent mieux distribuer les forces grâce à un temps de contact avec le sol plus long, et réduire efficacement les contraintes maximales. En observant la relation entre la taille du corps et la vitesse, on remarque une augmentation initiale de la vitesse, grâce à l’allongement des foulées. Mais par la suite, les contraintes deviennent telles que les stratégies de réduction de celles-ci deviennent inefficaces. Au-dessus de cette masse corporelle, nous nous attendrions à une diminution de la vitesse pour soulager les contraintes, et c’est en effet ce qui se passe (Figure 4). Mais la masse à partir de laquelle la vitesse décroît semble dépendre de la stratégie de réduction des contraintes utilisée – ce qui pourrait être utile pour estimer les vitesses des animaux géants disparus.
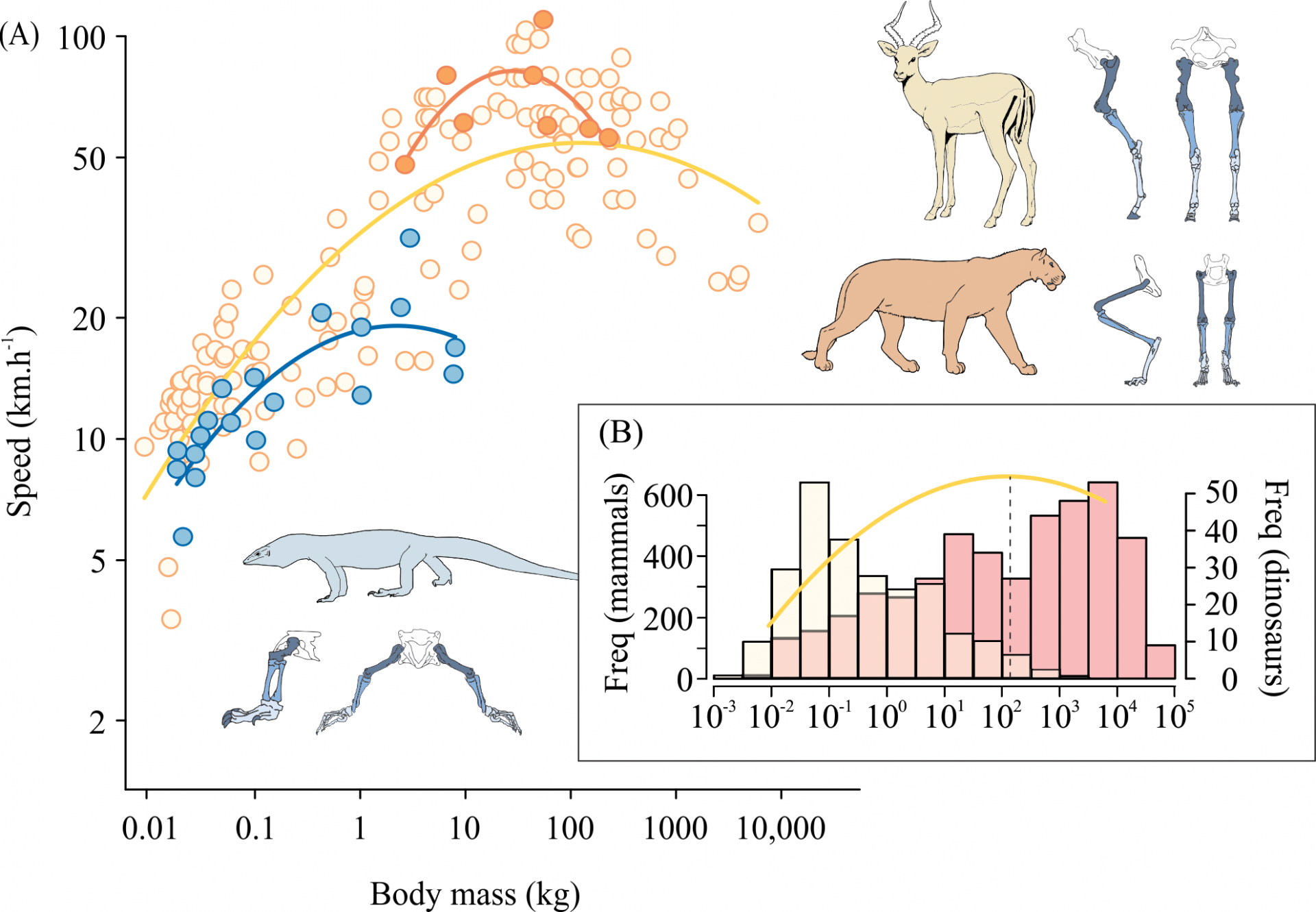
(A) Vitesses de pointe atteintes par des Mammifères non Félidés (n = 142, en jaune), des Félidés (n = 8, en orange) et des Varanidés (n = 19, en bleu) de différentes tailles. La masse pour laquelle la vitesse est optimale correspond au sommet de chaque courbe. (B) Distributions des masses chez les Mammifères actuels (n = 2919, barres jaunes) et chez les espèces de dinosaures éteints (n = 406, barres rouges) mises en relation avec la courbe de relation masse-vitesse des Mammifères non Félidés.
La figure 4 montre la relation entre la masse et la vitesse de course pour trois groupes d’animaux pour lesquels nous disposons de données complètes. Chez les Varanidés, on voit des changements dans les os et les muscles, mais à cause de leur posture accroupie, avec leurs membres en position transversale, il est peu probable qu’ils puissent se redresser. La masse à partir de laquelle leur vitesse commence à décliner est donc relativement faible. Les Mammifères utilisent un plus grand nombre de stratégies pour gérer le problème de la taille : certains (par exemple les ongulés) montrent des changements dans leurs os, leurs muscles et leur posture, en se redressant quand leur taille augmente. L’effet combiné de ces facteurs est probablement responsable de la capacité des Mammifères à atteindre d’importantes masses corporelles avant que l’augmentation des contraintes liées à la taille ne fasse diminuer leur vitesse. Les Félidés apparaissent comme en situation intermédiaire entre ces deux groupes : leurs os changent, mais pas leurs muscles ; ils ont une posture accroupie mais leurs membres sont en position parasagittale, et cette posture ne change pas avec leur masse. Les Félidés contiennent certains des animaux courant le plus vite sur Terre […] mais les mécanismes de réduction des contraintes ne sont pas aussi importants que chez d’autres Mammifères, ce qui explique probablement la masse plus faible à laquelle leur vitesse commence à diminuer.
La masse corporelle est la caractéristique la plus évidente et sans doute la plus fondamentale d’un organisme, ayant des conséquences sur de nombreux attributs de sa vie, de son écologie et de son évolution. Il est étonnant de voir que si peu d’espèces ont une masse supérieure à la masse optimale pour la vitesse (Figure 4A). De plus, ce nombre pourrait être biaisé par la plus grande représentation des gros Mammifères dans la littérature scientifique. En fait, lorsque l’on observe la distribution des masses avec un jeu de données plus complet sur les Mammifères (n = 2 919), on observe qu’il y a un nombre élevé de faibles masses corporelles (Figure 4B). Cela montre qu’un pourcentage très faible des espèces de Mammifères existant (3,8 %) sont au-dessus de la masse autorisant une vitesse optimale, et ce nombre est à peine plus élevé (8 %) si l’on inclut les Mammifères éteints. Chez les Varanidés on retrouve une distibution similaire : seules 21 % des espèces sont au-dessus de l’optimum. Lorsque l’on inclut des données additionnelles sur des Varanidés (n = 45) et d’autres espèces de lézards (n = 185), il y a même encore moins d’espèces au-dessus de l’optimum (5,4 %). Cela suggère que les espèces grandes et lentes ne réussissent pas dans la nature. Par conséquent, si les dinosaures employaient les mêmes stratégies de réduction des contraintes que les Mammifères, nous pourrions nous attendre à ce qu’une part similaire de ces espèces soit au-dessus de la masse permettant une vitesse optimale. Mais la distribution que l’on observe chez ces géants éteints est différente (Figure 4B). En utilisant les estimations sur la masse des dinosaures, on observe un pourcentage beaucoup plus important de dinosaures au-dessus de cet optimum : 81 % montrent une taille supérieure à la masse autorisant la vitesse optimale prédite pour les Mammifères. Même en tenant compte de la plus faible préservation des nombreuses espèces de dinosaures de petite taille, le nombre de dinosaures au-dessus de l’optimum est plus élevé que ce que nous pourrions prédire d’après la distribution des espèces existantes.
Comment ces animaux ont-ils pu être aussi grands ? Il pourrait y avoir deux explications à l’absence de géants parmi les espèces actuelles. Soit les effets combinés des facteurs environnementaux, écologiques et biologiques ont changé du tout au tout (ou continuent à changer trop rapidement) et ne permettent plus à des animaux gros et lents de réussir dans la nature. Seconde hypothèse, les dinosaures pourraient avoir utilisé des stratégies de réduction des contraintes que nous ne connaissons pas chez les espèces actuelles, et qui leur auraient permis d’atteindre des vitesses élevées même avec une masse importante. La reconstruction de la façon dont se déplaçaient ces géants disparus passera par des modélisations biomécaniques et des simulations informatiques rigoureuses, comparées aux données expérimentales chez les animaux actuels. Ces approches permettront de comprendre où sont passés tous les géants. À moins que de nouvelles stratégies apparaissent pour contourner les contraintes liées à la taille, ou que des taxons soient capables de faire avec les éventuels effets délétères d’une faible vitesse, nous pourrions ne jamais revoir de géants.
Article Source: Where Have All the Giants Gone? How Animals Deal with the Problem of Size
Dick TJM, Clemente CJ (2017) Where Have All the Giants Gone? How Animals Deal with the Problem of Size. PLOS Biology 15(1): e2000473. doi: 10.1371/journal.pbio.2000473