Présentation de la démarche entreprise par Meselson et Stahl pour établir le mode de réplication de la molécule d'ADN.
Introduction
Cette expérience date de 1958. Elle permet de démontrer le caractère semi-conservatif de la multiplication de la molécule d’ADN chez les bactéries. Cette expérience a pu être réalisée grâce à plusieurs mises au point techniques :
- Meselson et Stahl mettent au point une technique d’obtention d'un gradient de densité par centrifugation.
En utilisant du chlorure de césium de densité moyenne 1,72, ils obtiennent, après 24 h de centrifugation à grande vitesse, un gradient de densité d’environ 1,70 à 1,75. Cette gamme englobe la densité de l’ADN (1,710). - Ils cultivent les bactéries dans un milieu dont les substances organiques utilisées comme source d’azote contiennent de l’azote lourd (15N). Au cours de la culture, toutes les molécules azotées et en particulier l’ADN contiennent une forte proportion d’azote 15N.
L’ADN « lourd » a une densité de 1,724 et peut être distingué de l’ADN « léger » (1,710). - Ils mettent au point une méthode qui permet de synchroniser pendant quelques générations la division des bactéries.
Le problème à résoudre
Depuis Watson et Crick (1953), on sait que l’ADN est une molécule formée de deux brins antiparallèles, formant une double hélice. Dès leur publication originale sur la structure de l’ADN, Watson et Crick ont proposé que cette double hélice puisse s’ouvrir, permettant ainsi la synthèse de nouveaux brins, complémentaires des brins originaux.
L’ADN peut ainsi servir de matrice à sa propre réplication, étape essentielle du cycle cellulaire. Cette duplication de l’ADN (et donc des chromatides) permet de passer de chromosomes à une chromatide à des chromosomes possédant deux chromatides identiques, portant la même information génétique. Lors de la mitose, ces deux chromatides sont réparties, chaque cellule-fille héritant d’une chromatide de chaque chromosome. On obtient ainsi deux cellules possédant la même information génétique que la cellule-mère.
Le problème qui se posait à Meselson et Stahl était alors de comprendre comment se réalisait cette réplication : selon quelles modalités passe-t-on d’une molécule d’ADN formée de deux brins à deux molécules d’ADN bicaténaires identiques ?
Les hypothèses
Pour expliquer la duplication d’un ADN bicaténaire, trois modèles ont été proposés. Ces modèles se basent tous sur l’utilisation de la molécule d’ADN « mère » comme matrice pour sa réplication, mais selon des modalités différentes :
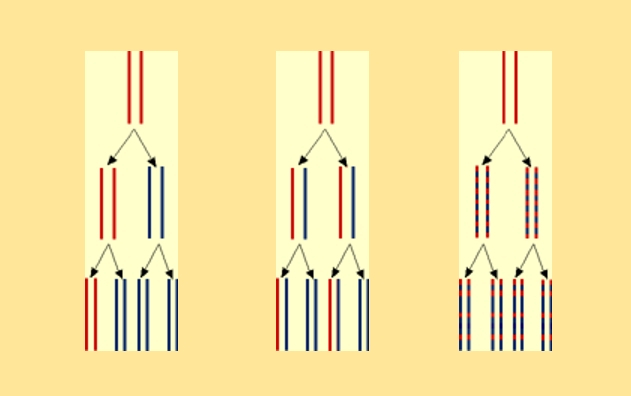
Ce schéma présente le devenir de l’ADN chez trois générations de cellules successives, selon les trois hypothèses de mode de réplication de l’ADN.
Hypothèse 1, à gauche : modèle conservatif
À partir d’une molécule d’ADN bicaténaire « mère », on forme une nouvelle molécule d’ADN bicaténaire. On garde donc ici une molécule « mère », non modifiée (elle est donc conservée), tout en « créant » une nouvelle molécule (« fille »).
Hypothèse 2, au centre : modèle semi-conservatif
On dissocie les deux brins de la molécule d’ADN bicaténaire « mère ».
Chaque brin sert donc de matrice à la synthèse d’un brin complémentaire, l’ensemble reformant une molécule d’ADN bicaténaire. Chaque nouvelle molécule « fille » ne conserve donc que la moitié de la molécule « mère ».
Hypothèse 3, à droite : modèle dispersif
On ne conserve aucun brin intact.
La copie se réalise par fragments dispersés dans l’ensemble de l’ADN, permettant de former les deux molécules d’ADN bicaténaires « filles ».
Légendes des couleurs
Rouge : Molécule d'ADN "mère" et son devenir. Bleu : ADN néo-formé.
L’expérience : résultats observés
Des bactéries cultivées depuis longtemps en présence de molécules azotées 15N sont repiquées sur un milieu contenant des molécules azotées 14N et permettant la synchronisation des divisions. Des fractions sont prélevées après différents temps correspondant à 1, 2, 3… divisions. L’ADN est extrait, placé dans la solution de chlorure de césium et centrifugé 24 h à 100 000 g. La position des ADN est repérée par une mesure de la densité optique.
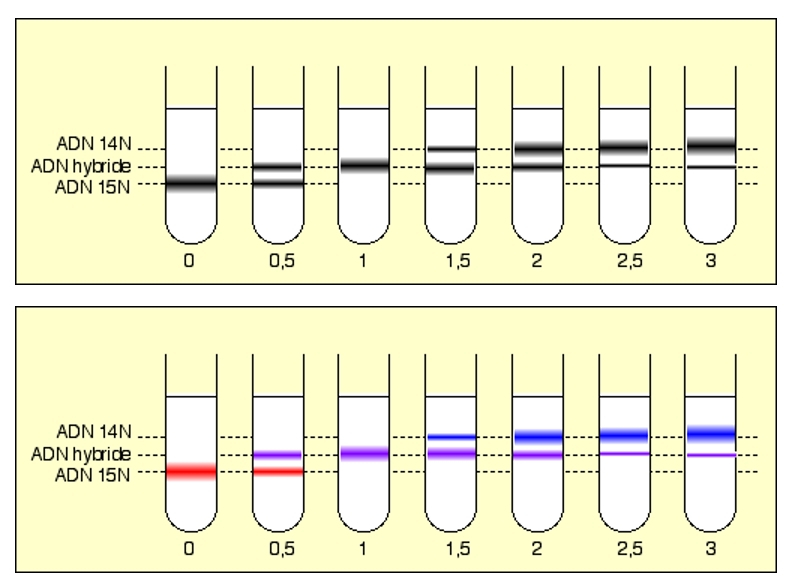
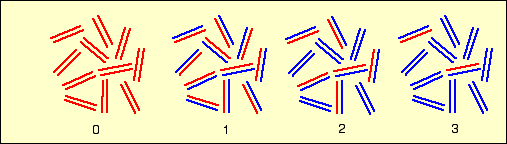
Ces schémas représentent la position des différentes bandes d'ADN observées au cours du temps (divisions successives), après centrifugation dans le gradient de chlorure de césium. Les chiffres donnent le nombre de divisions.
Le schéma du bas correspond à une interprétation colorée de celui du haut.
Après une génération, tout l’ADN est hybride (du point de vue de sa densité). Il n’y a plus d’ADN 15N.
Ensuite, l’ADN hybride disparaît progressivement au profit d’ADN « léger » (14N).
L’expérience : comparaison avec les modèles
L’expérience de Meselson et Stahl montre donc la présence d’un ADN hybride au bout d’une génération cellulaire. Or, qu’attend-on pour les trois modèles proposés ?
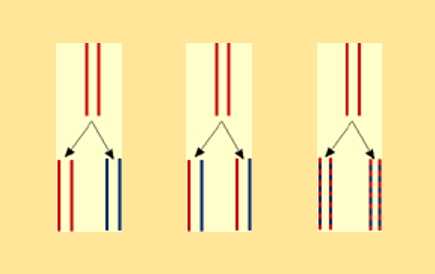
Ces schémas permettent de comparer le résultat obtenu après une génération, soit de l'ADN hybride, avec les résultats attendus selon les trois modèles de réplication de l'ADN envisagés.
A gauche, résultat attendu pour le modèle conservatif : ADN lourd (15N) et ADN léger (14N)
Au centre, résultat attendu pour le modèle semi-conservatif : ADN hybride (molécules formées d'un brin lourd et d'un brin léger)
A droite, résultat attendu pour le modèle dispersif: ADN hybride
On peut donc, dès cette première observation, rejeter le modèle conservatif.
Au bout de deux générations cellulaires, Meselson et Stahl observent la présence d’ADN hybride et d’ADN léger. Ceci permet de conclure quant aux deux modèles restants :
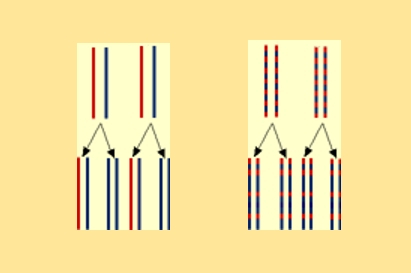
Ces schémas permettent de confronter les résultats obtenus après deux générations, à savoir de l'ADN hybride et de l'ADN léger, avec les résultats attendus selon les deux modèles encore possibles suite aux observations précédentes (après une génération).
A gauche, résultat attendu pour le modèle semi-conservatif : ADN hybride et ADN léger
A droite, résultat attendu pour le modèle dispersif : ADN hybride
En conclusion, seul le modèle semi-conservatif permet d’aboutir à une concordance entre résultats attendus et résultats observés.
Conclusion
L’expérience de Meselson et Stahl permet donc de mettre en évidence le fait que la réplication de l'ADN se réalise selon un mode semi-conservatif.
Au début de l'expérience tout l'ADN des bactéries est formé de deux brins d'ADN lourd (rouge).
A la première génération, après une réplication en milieu contenant 14N, tout l’ADN est « hybride » et constitué d’un ancien brin « lourd » (15N, ici en rouge) et d’un nouveau brin « léger » (14N, ici en bleu).
A la deuxième génération la moitié des fragments d'ADN est hybride (un ancien brin rouge et un nouveau brin bleu) et l'autre moitié de l'ADN est constitué de deux nouveaux brins légers (deux brins bleus).
Cette conclusion a été depuis confirmée par des études plus précises, pour aboutir au modèle actuel de fonctionnement de la réplication.
Quelques points importants de cette expérience sont à noter : tout d’abord le fait qu’il est nécessaire de séparer les ADN sur un gradient permettant de mettre en évidence leurs très faibles différences de densités ; une « simple » centrifugation ne suffit pas. L’utilisation d’un gradient de chlorure de césium est donc un point fondamental du protocole. De même, ces observations n’ont été possibles que parce que Meselson et Stahl avaient réussi à obtenir des populations de bactéries synchrones (pendant quelques générations).
De nombreux auteurs omettent ces points fondamentaux… Ce qui fait perdre tout sens à leurs conclusions…