

L'évolution est souvent vue comme un processus long, invisible à l'échelle d'une vie humaine. Après avoir expliqué pourquoi cette conception est fausse et montré quelles sont les méthodes utilisées pour observer l'évolution des populations, cet article présente deux exemples d'évolution rapide : celle des phalènes du bouleau et celles des pinsons des Galápagos.
L’évolution biologique a été introduite au xixe siècle comme un processus extrêmement lent, qui ne peut pas être observé directement. Cette idée a dominé la recherche académique au cours du xxe siècle, et reste commune chez les scientifiques, les enseignants, et le grand public.
Pourtant, les cinquante dernières années ont vu s’accumuler des avancées scientifiques qui montrent que les espèces peuvent évoluer significativement en quelques générations. Ces découvertes ouvrent une nouvelle fenêtre qui permet de comprendre les mécanismes de l’évolution avec plus de finesse, en les observant directement. Par ailleurs, les processus évolutifs doivent être pris en compte dans les rapports que les activités humaines entretiennent avec le vivant (gestion des maladies infectieuses, des agrosystèmes et écosystèmes…).
L’évolution darwinienne est née lente
D’un phénomène non observable…
L’évolution des espèces se traduit par des changements de leur phénotype (par exemple leur morphologie, leur physiologie, leur comportement…) dus à des changements génétiques. Ces changements peuvent mener à la disparition d’espèces ou à la formation de nouvelles espèces.
Ainsi, on sait aujourd’hui qu’une lignée de dinosaures théropodes a donné naissance aux oiseaux. Les 10 000 espèces d’oiseaux connues aujourd’hui, des manchots aux colibris, en passant par les poules et les moineaux, sont donc toutes issues d’un même ancêtre commun tandis que toutes les autres lignées de dinosaures se sont éteintes. Ces modifications de la biodiversité se sont déroulées sur plusieurs dizaines de millions d’années, à un rythme bien trop lent pour être perçu par un observateur humain.
L’idée que l’évolution est lente règne dans l’imaginaire populaire, et non sans raison : elle prend sa source dès la genèse de la biologie évolutive, avec Darwin et son Origine des espèces publiée en 1859. Le livre synthétise un large faisceau d’indices qui suggèrent que les espèces actuelles sont toutes apparentées et ont évolué à partir d’espèces anciennes qui se sont diversifiées et se sont modifiées par l’action de la sélection naturelle. Darwin présente ses arguments comme le ferait un historien : le support de ses hypothèses vient de traces laissées par des événements passés. Cette idée a dominé la recherche académique au cours du xxe siècle. À partir des années 1910, la théorie de l’évolution s’est progressivement renforcée de solides bases théoriques et mathématiques, tandis que l’étude du registre fossile et des organismes actuels continuait à la soutenir et à l’enrichir. Cependant, les processus de sélection naturelle et d’évolution restaient non observés, et supposés non observables.
Ce manque d’observations directes a certainement facilité les attaques de religieux fondamentalistes contre l’enseignement de l’évolution à l’école, victorieuses pour un temps aux États-Unis dans les années 1920-1930. Plus largement, elle a permis de perpétuer le mythe selon lequel l’évolution n’est qu’une hypothèse sans preuves, alors qu’il s’agit d’une des théories scientifiques les plus vérifiées.
… à une évidence mathématique
Formellement, l’évolution peut être définie comme le changement des fréquences de versions de gènes (allèles) dans une population. Cette dernière se définit comme un ensemble d’organismes de la même espèce se reproduisant entre eux et vivant dans un lieu donné, à un moment donné.
Dans le cadre de cette définition, l’évolution génétique entre chaque génération est une quasi-nécessité logique. En effet, même en l’absence de sélection, quand la propension des individus à survivre et à se reproduire est indépendante des différences phénotypiques et génétiques, la composition génétique de la population est déterminée par une sorte de tirage aléatoire avec remise : la dérive génétique. Dès lors qu’il existe des différences génétiques, le phénomène de dérive est suffisant pour produire des changements génétiques entre générations.
Cependant, l’évolution par dérive génétique est lente, avec une vitesse inversement proportionnelle à la taille de la population. De plus, elle n’a pas de direction préférentielle et ne favorise donc généralement pas l’adaptation des organismes à leur environnement. Les variations des fréquences alléliques dans une population peuvent également être due à des mutations, ou à la migration d’individus entre différentes populations.
La question de savoir si l’évolution se produit aujourd’hui est donc triviale : la réponse est « oui », par nécessité logique. Il est plus intéressant de se demander si l’évolution peut se produire rapidement, si la sélection naturelle est à l’œuvre, et si la vitesse de l’évolution change significativement la biologie et l’écologie des organismes.
Nous allons voir que la lenteur de l’évolution a été progressivement démentie, par petites touches, dans la seconde moitié du xxe siècle. L’évolution par sélection naturelle peut être suffisamment rapide pour être observée et pour changer les relations écologiques entre espèces. Dès lors, il devient clair qu’il est important de prendre en compte l’impact de l’évolution dans les activités humaines.
Comment observer l’évolution dans la nature ?
Observer est nécessaire, mais insuffisant
La première approche pour mesurer l’évolution consiste à suivre une population dans le temps, et à quantifier les changements dans un caractère phénotypique donné. Suivre une population sauvage pendant de nombreuses années et mesurer des caractères précisément peut poser des problèmes logistiques majeurs. Cependant, cette approche était évidemment disponible dès l’époque de Darwin et même avant, mais n’a pas vraiment été exploitée. Sa principale limite, identifiée dès le xixe siècle, est qu’un changement phénotypique observé pourrait être dû à l’effet direct de l’environnement sur le phénotype des organismes (plasticité phénotypique), plutôt qu’à la transmission de parents à enfants de caractères sélectionnés (évolution par sélection naturelle). Par exemple, un réchauffement du climat pourrait augmenter la taille moyenne d’une population de plantes par (i) plasticité, si le réchauffement augmente l’activité des enzymes qui contrôlent la croissance, par un effet thermodynamique direct ; ou (ii) évolution, si le réchauffement réduit la viabilité des plantes les plus petites, incapables d’absorber suffisamment d’eau avec leurs racines peu profondes, et que les allèles rendant les plantes plus grandes sont transmis préférentiellement à la génération suivante. Phénotypiquement, le changement pourrait être le même dans les deux cas, et une simple observation ne permettrait pas de dire si l’évolution contribue au changement (voir Figure 1).
Séquencer l’ADN
Heureusement, des avancées conceptuelles et techniques ont permis de dépasser ce problème au cours du xxe siècle. D’abord, la découverte de l’ADN comme support de la transmission des caractères , puis la possibilité de séquencer l’ADN d’individus de plus en plus rapidement (méthode de Sanger, puis diverses méthodes de séquençage haut débit) qui a permis de générer d’énormes quantités de données génétiques. Il est possible d’estimer les fréquences des allèles d’un gène à différents moments, et donc de mesurer l’évolution à l’échelle d’un ou de plusieurs gènes. Bien que très puissantes, parce que centrées sur le matériau ultime de l’évolution, ces méthodes dites de génétique des populations (voir Figure 1) présentent des limites quand il s’agit de comprendre l’évolution d’un organisme dans son ensemble. Notamment, il reste très difficile de lier les données moléculaires à des caractères phénotypiques, surtout pour les caractères continus (taille ou masse par exemple). Par ailleurs, l’ensemble des gènes considérés dans une étude n’explique souvent que quelques pourcents de la variation phénotypique existante. De plus, il est souvent impossible de séparer le rôle de la sélection naturelle d’autres processus (mutations, dérive génétique, migrations) avec des données moléculaires seules.
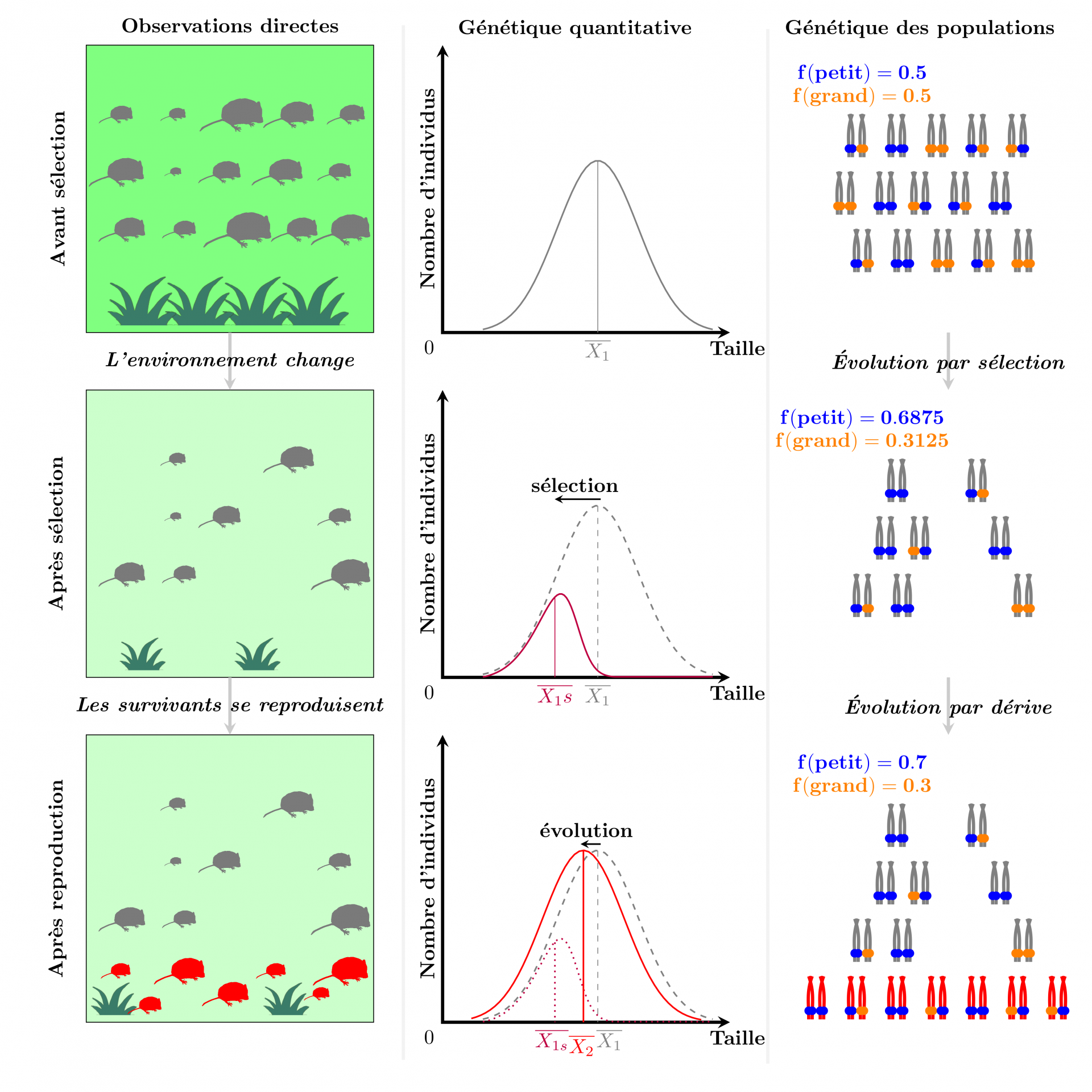
Dans la première génération (individus et chromosomes gris), les grands individus survivent mal dans le nouvel environnement, pauvre en nourriture, ce qui entraîne un changement de la taille moyenne dans la population. Le changement persiste en partie après reproduction, dans la seconde génération (individus et chromosomes rouges) car les différences de taille sont en partie dues à des différences génétiques. L’observation directe ne permet pas de confirmer que le changement de taille est évolutif plutôt que plastique : la réduction de taille dans la seconde génération pourrait être directement due à des problèmes de croissance liés au manque de nourriture. Du point de vue de la génétique quantitative, le changement de taille permet de mesurer la sélection. Après avoir estimé l’héritabilité de ce changement de taille (en comparant la similarité des individus à leurs parents), cette donnée peut être utilisé pour prédire l’évolution. Du point de vue de la génétique des populations, les données moléculaires montrent qu’il existe deux allèles d’un même gène qui contrôle en partie la variation en taille. Puisque la sélection supprime les individus les plus grands, les allèles orange (donnant de grands individus) ont plus de chances de ne pas être transmis, et les allèles bleus (donnant de petits individus) deviennent plus communs après la sélection. Après reproduction, la fréquence des allèles bleus augmente un peu plus, sous l’effet aléatoire de la dérive génétique. Notez que la taille des individus ne dépend pas uniquement du gène étudié (sans quoi il y aurait au plus trois tailles différentes) mais également d’autres gènes et de facteurs environnementaux.
Méthodes statistiques
De nouvelles méthodes centrées sur les phénotypes et basées sur des avancées mathématiques et statistiques sont complémentaires des méthodes moléculaires, et ont été primordiales pour mesurer et comprendre l’évolution en temps réel. Ces méthodes, dites de génétique quantitative, mesurent la ressemblance entre individus apparentés pour distinguer les variations phénotypiques dues à des différences génétiques de celles dues à des différences environnementales. La méthode la plus simple utilise une régression linéaire entre parents et enfants (voir Figure 2). Cela permet de quantifier précisément et objectivement la force de la sélection naturelle agissant sur un caractère ainsi que l’héritabilité de ce caractère (voir Figure 2). La connaissance de ces deux paramètres permet de prédire l’évolution génétique attendue à la prochaine génération en réponse à la pression de sélection (réponse = force de la sélection × héritabilité).
D’autres méthodes de génétique quantitative permettent de mesurer le rythme de changement génétique en prenant en compte l’effet de l’ensemble du génome sur les traits considérés, avec peu ou pas d’effort de séquençage : il suffit de connaître les relations de parenté entre individus. Ces méthodes ont été développées avec grand succès pour l’amélioration variétale en agriculture à partir des années 70, avant d’être adoptées pour l’étude des organismes sauvages dans les années 2000.
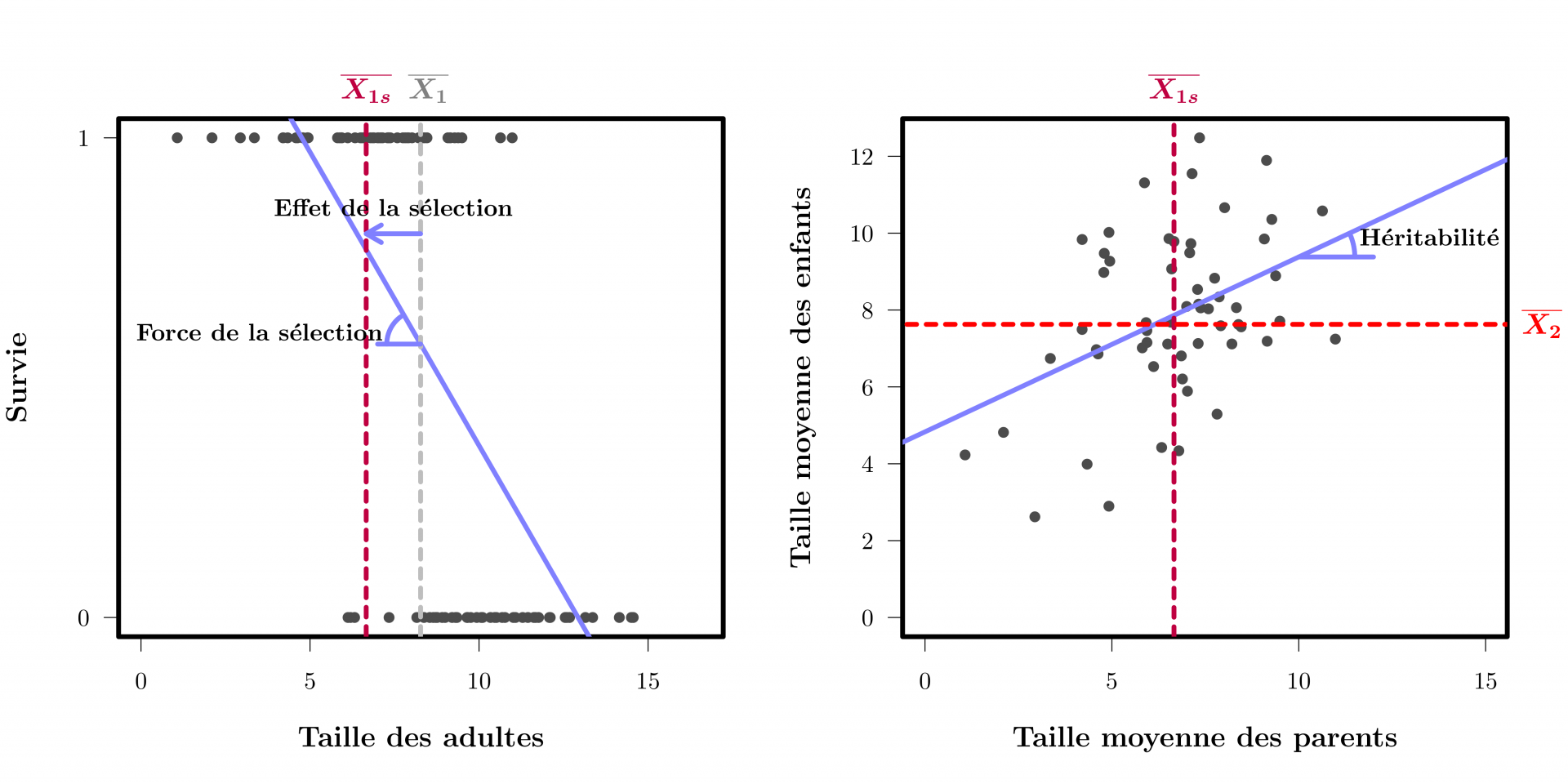
Ces deux graphiques développent l’approche de génétique quantitative présentée Figure 1. Le graphique à gauche montre comment une régression linéaire permet de mesurer la pression de sélection agissant sur la taille. Chaque point représente un individu, avec sa taille en abscisses et la survie jusqu’à l’âge de reproduction en ordonnées (0 pour mort avant de pouvoir se reproduire, 1 pour survie). La pente de la régression est une mesure objective de la force de la sélection. L’effet de la sélection, c’est à dire le changement de taille moyenne après sélection, peut-être obtenu en multipliant cette pente par la variance en taille et en divisant par la survie moyenne (ce qui est équivalent à la covariance entre la taille et la survie, divisé par la survie moyenne). Pour simplifier, on ne représente ici que la sélection due à la viabilité, et on ignore la sélection liée à la reproduction.
Sur le graphique de droite chaque point correspond à la taille moyenne des enfants d’un couple (en ordonnées) en fonction de la taille moyenne de leurs deux parents (en abscisses) montre comment la régression linéaire peut aussi être utilisée pour mesurer l’héritabilité, c’est-à-dire la proportion de variation d’un caractère qui est due aux gènes. La pente entre la taille moyenne des parents et la taille moyenne des enfants reflète la transmission de gènes ayant un effet sur la taille ; plus la pente est forte, plus les gènes expliquent une part importante de la variation en taille. En multipliant l’effet de la sélection par l’héritabilité, on peut prédire le changement génétique entre les deux générations.
Ces deux techniques sont facilement généralisables pour mesure la sélection qui agit via la fécondité plutôt que la survie, pour mesurer la sélection agissant sur plusieurs traits simultanément, pour corriger pour les sources de similarités non-génétiques entre apparentés, ou encore pour estimer le rythme de changement génétique sur plusieurs générations...
L’évolution en direct
Papillons et pinsons montrent l’exemple

Une des premières observations de l’évolution en action concerne la phalène du bouleau, ce papillon de nuit dont il existe deux formes, une blanche, une noire (Figure 3). La forme noire, extrêmement rare avant la révolution industrielle devient progressivement abondante entre 1810 et 1895. À l’époque déjà, la cause première de ce changement ne faisait guère de doute : les rejets de suies par les centrales à charbon. En revanche, le mécanisme biologique à l’œuvre était ambigu (évolution ou induction chimique directe ?) jusqu’à ce que les expériences des années 1950 montrent que la sélection naturelle était à l’œuvre via la prédation préférentielle des papillons les moins bien camouflés : les noirs sur l’écorce naturellement blanche des bouleaux, les blancs sur les arbres et les murs noircis par la pollution au charbon. Un temps critiqué, ce résultat a été confirmé et renforcé en 2012 par de nouvelles expériences et analyses génétiques (Cook 2003, 2012, 2013, Saccheri 2008). Le mécanisme génétique à l’origine des deux couleurs a été identifié, et des collections anciennes ont permis de suivre les changements de fréquences alléliques rétrospectivement. Ces changements expliquaient effectivement les variations des couleurs observées et correspondaient à celles prédites par la pression de prédation observée (Figure 4). L’étude des phalènes a fourni une première observation complète et rigoureuse du phénomène d’évolution par sélection naturelle dans la nature, et a démontré que l’évolution pouvait être assez rapide pour être visible sur une vie humaine. Mais l’étude d’autres organismes sauvages a montré que l’évolution pouvait être observée sur une durée beaucoup plus courte.
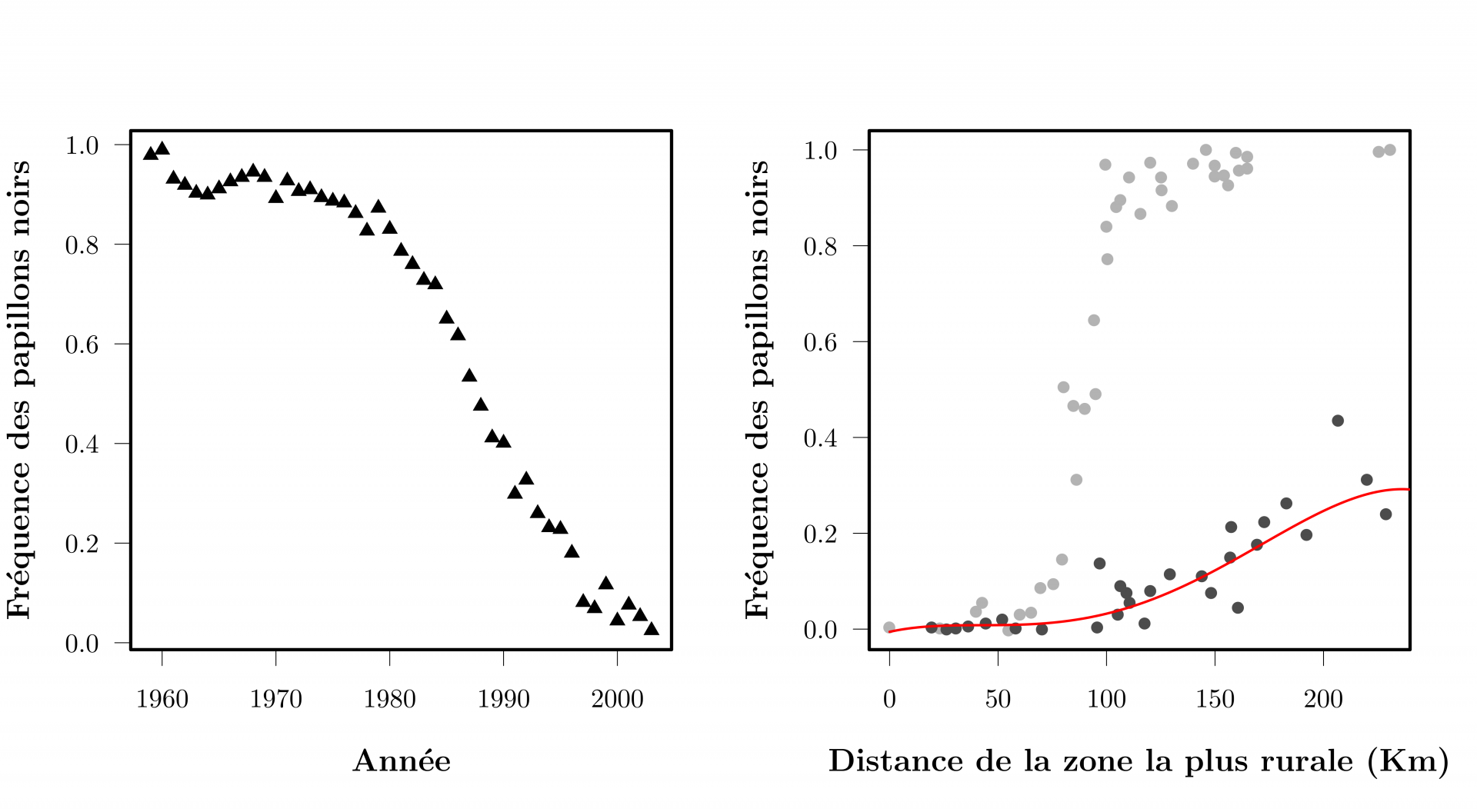
Le graphique à gauche montre la fréquence des phalènes du bouleau noires chaque année entre 1959 et 2003 dans le nord-ouest de l’Angleterre. Le graphique à droite montre la fréquence des phalènes du bouleau noires le long d’un transect partant d’Abersoch (0 km) une zone rurale du nord-ouest de l’Angleterre, et allant jusqu’à Leeds via Liverpool et Manchester, des zones urbaines et plus polluées. Les points gris clair correspondent à des mesures réalisées entre 1964 et 1975, les points gris sombres correspondent à des mesures réalisées en 2002. La chute importante de la fréquence des phalènes noires entre ces deux périodes s’explique par une diminution globale de la pollution (diminution qui a inversé la sélection naturelle, désormais contre les phalènes noires et en faveur des blanches). La ligne rouge représente les fréquences prédites pour 2002 en n’utilisant que les données de 1964 à 1975, la force de la sélection naturelle contre les phalènes noires et la dispersion moyenne des phalènes.
Adapté à partir des données présentées dans Cook (2003) The Rise and Fall of the Carbonaria Form of the Peppered Moth. The Quarterly Review of Biology 78(4). et dans Saccheri & al. (2008) Selection and gene flow on a diminishing cline of melanic peppered moths. Proceedings of the National Academy of Sciences of the USA 105(42).

Les points représentent la taille moyenne des becs (après transformation par analyse des composantes multiples) chaque année. Les flèches représentent la force et la direction de la sélection (plus positive en rouge, plus négative en bleu). Les changements de taille du bec sont fortement corrélés à la force de la sélection l’année précédente (coefficient de détermination 0.70).
À partir des données présentées dans Grant & Grant (2002) Unpredictable evolution in a 30-year study of Darwin’s finches. Science, 296, 707–711.
En 1973, Peter et Rosemary Grant démarrent l’étude d’animaux sauvages sans doute la plus détaillée de tous les temps. Sur certaines îles de l’archipel des Galápagos, les Grant et leurs collaborateurs ont marqué chaque pinson des Galápagos (Figure 5) et noté l’histoire de chaque individu en détail : date de naissance, de disparition, reproduction, partenaires, dimensions du bec, des ailes et des pattes… ces données phénotypiques ont été complémentées de données génétiques de qualité croissante. Toutes ces données, accumulées depuis 45 années ont permis de calculer la force de la sélection naturelle, la transmission génétique des caractères, et le rythme de l’évolution. Les Grant ont ainsi montré que les cycles El Niño de pluies intenses et de sécheresses provoquent certaines années une intense sélection naturelle sur la taille des becs des pinsons. Cette sélection se traduit par une évolution génétique des populations, apparente en seulement deux générations. Les fluctuations climatiques, les changements d’abondances des graines, et la compétition entre les différentes espèces de pinsons sont autant de pressions de sélection, toujours changeantes. La taille et de la forme des becs évoluent rapidement presque constamment, dans un sens, puis dans l’autre (Figure 6), de sorte que seul un suivi permanent des populations permet de réaliser l’amplitude et la rapidité du phénomène (Grant & Grant 2003; Weiner, 1994).

Plus impressionnant encore, cette étude a permis d’observer l’apparition d’une nouvelle espèce de pinsons en temps réel. Des chercheurs ont pu assister à la spéciation, depuis son initiation avec l’hybridation de deux individus appartenant à deux espèces différentes de pinsons, jusqu’à l’isolement reproducteur total, le tout se produisant en trois générations (Lamichhaney & al. 2017). La nouvelle espèce est différente morphologiquement des espèces qui lui ont donné naissance, et s’est rapidement étendue en occupant une niche écologique encore vacante sur l’île Daphne Major. Ce mode de spéciation, par hybridation, peut sembler quelque peu hétérodoxe, mais l’étude des génomes a montré qu’il était commun chez les plantes, et régulier chez les animaux, et pourrait être une source majeure de spéciation rapide (Mallet & al. 2016).
Les études des pinsons et des phalènes ont été parmi les premières à montrer l’exemple. Un exemple qui a été abondamment suivi, nous permettant aujourd’hui d’émettre quelques idées générales sur le rythme et les moteurs de l’évolution contemporaine.

L’importance des changements environnementaux
Les études actuelles suggèrent que l’évolution est généralement lente dans les populations dont l’environnement ne change pas (Kingsolver & al. 2012). En effet, dans un environnement stable, il s’établit un équilibre entre les différentes pressions de sélection. Par exemple, les mésanges bleues adultes sont sélectionnées pour produire de plus gros poussins (qui survivent mieux), mais aussi pour limiter leur effort dans l’élevage des poussins et ainsi conserver une masse corporelle importante pour survivre jusqu’à la prochaine saison de reproduction. Les masses des poussins et celles des adultes sont sujettes à un conflit, et la sélection naturelle a poussé la population vers un point d’équilibre entre ces deux pressions de sélection (Thomson & al. 2017), équilibre qui ne sera déplacé que par de nouvelles mutations génétiques (procédé très lent), ou par un changement environnemental (procédé potentiellement immédiat).
Dans les populations sauvages éloignées des activités humaines, les exemples de changements environnementaux qui ont provoqué des changements évolutifs rapides sont souvent liés aux variations du climat (d’origine naturelle ou anthropique). De nombreuses espèces changent phénotypiquement en réponse aux variations climatiques, mais seulement une poignée d’études a pu démontrer le rôle des mécanismes évolutifs dans ces changements (par exemple Bonnet & al. 2017), peut-être parce que la plasticité phénotypique est plus importante dans ces conditions, mais certainement en partie aussi parce que les données nécessaires pour tester l’origine évolutive des changements n’existent pas dans la majorité des populations (Merilä & Hendry 2014).
Si les espèces éloignées des pressions humaines semblent souvent évoluer lentement, c’est tout l’inverse pour celles dont les activités humaines modifient l’environnement (Lefèvre & al. 2016 Chapitre 21), comme nous allons le voir ci-dessous.
Implications socio-économiques
L’étude de l’évolution contemporaine n’est pas seulement motivée par la curiosité scientifique. Le fait que l’évolution se manifeste à l’échelle de quelques années signifie qu’elle peut interagir avec les activités humaines.
Bactéries pathogènes, moustiques et plantes adventices, sont les cibles de nos attaques, avec presque toujours le même résultat : quand un nouvel antibiotique/insecticide/herbicide est développé, son efficacité est presque parfaite et l’abondance des organismes indésirables diminue drastiquement. Cependant, après quelques années, ces organismes reviennent en force et sont maintenant résistants à nos attaques. La coupable est l’évolution : des allèles de gènes qui étaient présents en très faible fréquence dans la population (ou plus rarement qui sont apparus aléatoirement après l’exposition) et qui permettent la résistance aux antibiotiques/insecticides/herbicides ont été sélectionnés positivement. Les individus porteurs ayant une meilleure probabilité de survie, ils se reproduisent rapidement et comblent le vide écologique laissé par la mort de la majorité des individus. Après quelques générations, les seuls individus présents seront tous porteurs des allèles de résistances. Il est toutefois possible de ralentir l’apparition de résistances en prenant en compte les processus évolutifs dans la création de substances antibiotiques, insecticides ou autres. Par exemple, en combinant plusieurs substances létales agissant sur des voies métaboliques distinctes, l’évolution d’une résistance est beaucoup moins probable, parce qu’elle nécessite l’apparition simultanée de mutations de résistance contre toutes les substances utilisées (Brown & Stankova 2017, Lefèvre & al. 2016 Chapitre 22).
Les activités de chasse et de pêche affectent les stocks non seulement démographiquement, mais aussi évolutivement, ce qui menace la pérennité de ces activités. Par exemple, la taille de nombreuses espèces de poisson diminue en réponse à une pression de pêche ciblant les grands individus, entraînant du même coup des changements génétiques dans le cycle de vie de ses espèces, diminuant leur succès reproducteur (Eikeset & al. 2013). De la même façon, la chasse sportive aux trophées constitue une pression sélective : la chasse visant les plus gros trophées a réduit la taille des cornes chez les mouflons ou celle des défenses chez éléphants en l’espace de quelques décennies. La réduction de la taille des cornes menace l’économie de la chasse et les financements alloués à la conservation des populations par les sociétés de chasse. Par ailleurs, les cornes sont des caractères sexuels secondaires impliqués dans le rut, et leur régression perturbe la reproduction de ces espèces diminuant ainsi l‘abondance des espèces au-delà de la perte des quelques individus chassés (Huparinen & Festa-Bianchet, 2017).
De nombreuses essences d’arbres sont également affectées par le réchauffement climatique, qui modifie leur zone de viabilité. Il est crucial de connaître le rythme auxquels les arbres sont capables d’évoluer génétiquement en réponse au climat pour pouvoir planter des espèces qui seront adaptées aux conditions climatiques jusqu’à leur maturité sexuelle plusieurs décennies plus tard (Franks & al. 2014).
L’être humain modifie très rapidement son propre environnement. Ces modifications entraînent en retour des pressions de sélection conduisant à une adaptation de notre espèce (Beauchamp, 2016). Les conséquences de cette évolution sont encore sujettes à un difficile débat, perturbé par les craintes de politiques eugénistes, mais il semble possible que notre dépendance médicale et technologique augmente (Lynch 2016).
En conclusion, il faut prendre garde à ne pas considérer les pressions de sélection comme fixes dans le temps. Ces pressions de sélection changent en effet sous l’effet de variations du milieu, que celles-ci soient d’origine naturelle ou anthropique. Des variations rapides dans les pressions de sélection peuvent mener en un temps court (quelques générations) à des changements phénotypiques importants, pouvant entraîner des spéciations.
Références
Références générales
- Weiner, 1994. The Beak of the Finch. (Prix Pulitzer 1995)
- Darwin, 1859. On the Origin of Species. (L’Origine des Espèces)
- Le livre est disponible, y compris en français, gratuitement à http://darwin-online.org.uk/contents.html#origin
- Lefèvre T., Raymond M., Thomas F. (2016) Biologie évolutive. 2nde édition.
En particulier Chapitres 21 (Évolution induite par les activités anthropiques) et 22 (Applications de la biologie évolutive).
Phalène du bouleau
- Cook, L. M., & Saccheri, I. J. (2013). The peppered moth and industrial melanism: evolution of a natural selection case study.. Heredity, 110(3), 207-212.
- Cook, L. M., B. S. Grant, I. J. Saccheri and J. Mallet. (2012). Selective bird predation on the peppered moth: the last experiment of Michael Majerus. Biology Letters online,:doi: 10.1098/rsbl.2011.1136.
- Cook (2003) The Rise and Fall of the Carbonaria Form of the Peppered Moth. The Quarterly Review of Biology 78(4)
- Saccheri & al. (2008) Selection and gene flow on a diminishing cline of melanic peppered moths. Proceedings of the National Academy of Sciences of the USA 105(42).
- Blog de Jerry Coyne : https://whyevolutionistrue.wordpress.com/2012/02/10/the-peppered-moth-story-is-solid
Évolution des pinsons des Galápagos, et spéciation par hybridation
- Grant, P.R. & Grant, B.R. (2002) Unpredictable evolution in a 30-year study of Darwin’s finches. Science, 296, 707–711.
- Lamichhaney & al. (2017) Rapid hybrid speciation in Darwin’s finches. Science, eaao4593.
- Mallet, J., Besansky, N. & Hahn, M.W. (2016) How reticulated are species? BioEssays, 38, 140–149.
Exemples d’évolution contemporaine, et difficultés à la détecter, dans des populations relativement non-perturbées
- Bonnet, T. & al. (2017) Bigger Is Fitter? Quantitative Genetic Decomposition of Selection Reveals an Adaptive Evolutionary Decline of Body Mass in a Wild Rodent Population. PLOS Biology, 15, e1002592.
Version popularisée en français: http://www.sciencejournalforkids.org/uploads/5/4/2/8/54289603/voles-article-fr.pdf - Kingsolver, J.G., Diamond, S.E., Siepielski, A.M. & Carlson, S.M. (2012) Synthetic analyses of phenotypic selection in natural populations: lessons, limitations and future directions. Evolutionary Ecology, 26, 1101–1118.
- Merilä, J. & Hendry, A.P. (2014) Climate change, adaptation, and phenotypic plasticity: The problem and the evidence. Evolutionary Applications, 7, 1–14.
- Thomson, C.E., Hadfield, J.D., Bayer, F., Cassinello, M., Crouch, N., Heap, E., Mittell, E. & Hadfield, J.D. (2017) Selection on parental performance opposes selection for larger body size in a wild population of blue tits. Evolution, 71, 716–732.
Résistances et stratégies évolutives de contrôle
- Brown J.S. & Stankova K. (2017) Game theory as a conceptual framework for managing insect pests. Current Opinion in Insect Science, 21, 26-32.
Exemples d’évolution contemporaine en réponse aux collectes humaines
- Franks, S.J., Weber, J.J. & Aitken, S.N. (2014) Evolutionary and plastic responses to climate change in terrestrial plant populations. Evolutionary Applications, 7, 123–139.
- Huparinen & Festa-Bianchet. Harvest-induced evolution: insights from aquatic and terrestrial systems 2017 Philosophical Transactions of The Royal Society B Biological Sciences 372 (1712)
- Eikeset & al. 2013. Economic repercussions of fisheries-induced evolution. Proceedings of the National Academy of Science USA. 110 (30).
Evolution humaine contemporaine
- Beauchamp, J.P. (2016) Genetic evidence for natural selection in humans in the contemporary United States. Proceedings of the National Academy of Sciences, 113, 7774–7779.
- Lynch, M. (2016) Mutation and Human Exceptionalism: Our Future Genetic Load. Genetics, 202, 869–875.



