

Savoir retrouver son nid ou s’orienter vers une source de nourriture est un processus adaptatif chez de nombreux animaux. En prenant l’exemple des insectes, sujets d’étude privilégiés en navigation, nous verrons différentes stratégies de navigation, leurs substrats neuronaux, et les implications écologiques et technologiques.
Les capacités de navigation exceptionnelles de certains vertébrés font les gros titres tant ces prouesses fascinent. Les oiseaux et tortues réalisent de remarquables migrations sur de très grandes distances. En comparaison, nos capacités de navigation semblent maigres aujourd’hui ; nous nous basons sur des signaux comme les panneaux de signalisation, mais peinons à nous retrouver lorsque notre GPS ne fonctionne plus en pleine forêt. Moins mis en lumière, les insectes ne sont pas en reste et ont démontré de remarquables capacités de navigation malgré leur cerveau, souvent plus petits qu’un grain de riz ! Les fourmis retrouvent avec succès leur nid après avoir parcouru un labyrinthe de cailloux et de brindilles, et le bogong, un petit papillon australien, parcourt plus de 2000 km dans sa vie. Rapporté à sa taille, c’est comme si un nouveau-né parcourait deux fois la distance entre la Suède et la Nouvelle-Zélande. Par rapport aux mammifères, les insectes présentent de nombreux avantages expérimentaux pratiques, autorisant souvent une compréhension plus fine des mécanismes mis en jeu dans la navigation spatiale. Dans cet article, nous présenterons un aperçu des différentes stratégies de navigation chez les insectes, leurs bases sensorielles et neuronales, ainsi que l’importance de ces études aujourd’hui, en termes de biologie de la conservation et de robotique.
Un aperçu des stratégies de navigation
Qu’est-ce que la navigation spatiale ?
La navigation peut être définie comme un mouvement dirigé vers un but et qui demande la réalisation de tâches cognitives complexes. Bien qu’il n’y ait pas de réel consensus sur la définition, la navigation se distingue des mécanismes élémentaires comme la cinèse (mouvement spontané en réponse à un stimulus) ou la taxie (mouvement dirigé vers un stimulus) par la distance qui sépare l’animal de son but, très grande en navigation, et de la présence de systèmes de planification complexes (1).
La navigation peut prendre des formes variées : orientation en ligne droite, migration et retour au nid (homing). Certaines espèces de scarabées bousiers cherchent s’éloigner en ligne droite de la bouse pour fuir la compétition (orientation en ligne droite). Les papillons monarques parcourent des milliers de kilomètres dans une direction précise vers un but lointain et inconnu (migration). Enfin, les abeilles, guêpes et fourmis, pour ne citer que ces insectes, explorent des environnements complexes à la recherche de nourriture, et doivent retrouver leur nid après un voyage d’exploration (homing). Notons qu’il existe d’autres contextes de navigation, qui ne seront pas abordés ici. Ainsi, l’exploration de l’environnement, qui repose sur le suivi de pistes de phéromones, de signaux intraspécifiques (danse d’orientation des abeilles) ou de mouvements stéréotypés (comportements de recherche) ne sera pas abordée dans cet article.
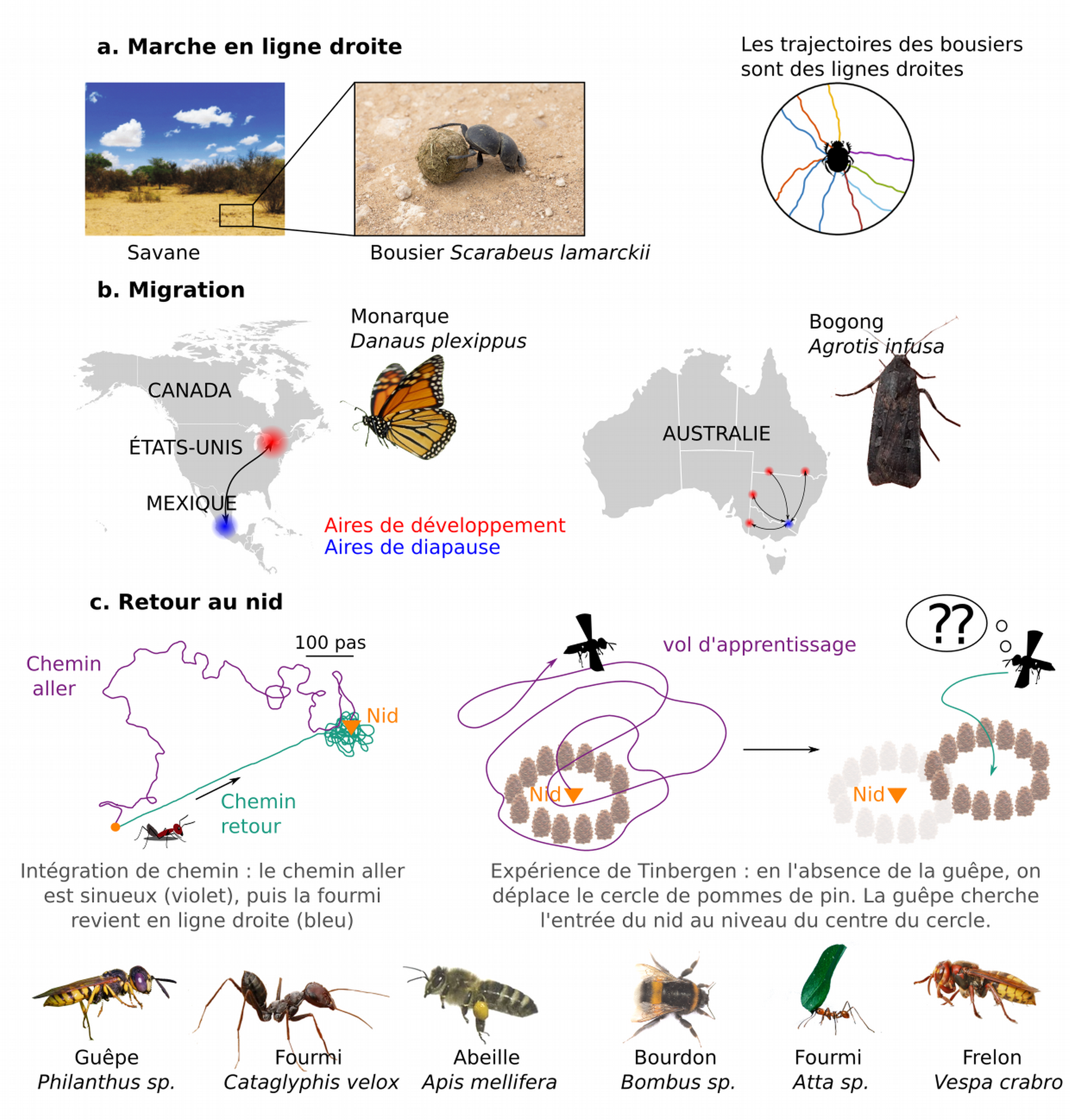
(a) Marche en ligne droite. Les bousiers roulent leur boule en ligne droite pour augmenter la distance avec la source le plus vite possible, et éviter ainsi tout conflit avec d’autres bousiers qui tenteraient de voler leur boule.
(b) Migration. Monarques et bogongs sont deux espèces de papillons réalisant une migration aller-retour (en une ou plusieurs générations) entre l’aire de développement (rouge) et l’aire de diapause (bleu).
(c) Retour au nid. De nombreux insectes, en particulier des Hyménoptères, explorent l’environnement avant de revenir vers leur nid. Des observations et expériences historiques ont mis en évidence l’existence d’intégration de chemin (le chemin aller en violet est sinueux tandis que le chemin retour en bleu est en ligne droite) et de mémoire spatiale chez les insectes (expérience de Tinbergen (2)).
Remarque : il existe d’autres contextes de navigation : chez les drosophiles par exemple, l’intégration de chemin permet d’explorer l’environnement tout en conservant la possibilité de revenir à une source de nourriture.
Crédits : Savane et trajectoires des bousiers : Khaldy et coll., 2020, CC BY, Journal of Comparative Physiology A ; Bousier : Michael Mayer, CC BY, Flickr ; Monarque : dérivé de Dwight Sipler, CC BY, Flickr ; Bogong : Paul Clémençon, CC BY-NC ; Intégration de chemin : d'après Stone et coll., 2017, Current Biology ; Fourmi Atta : Scott Bauer, CC BY, Wikimedia ; Abeille Apis : domaine public ; Fourmi Cataglyphis : dérivé de José María Escolano, CC BY-NC-SA, Flickr ; Frelon : dérivé de Bernie Kohl, CC BY-SA, Wikimedia ; Philanthus : Frupus, CC BY-NC, Flickr ; Bourdon : CC0.
Stratégies de navigation chez les insectes
Les stratégies de navigation présentées ci-dessous ont été historiquement proposées pour expliquer la navigation chez les insectes. Elles ne sont pas mutuellement exclusives : une même espèce peut utiliser plusieurs de ces stratégies. Lorsque le nid est « proche », les insectes pourraient utiliser des stratégies de guidage (partie 1.2.1). Lorsque le nid est « loin », les insectes pourraient utiliser des stratégies d’intégration de chemin (1.2.2), de suivi de chemin ou de route (séquence de snapshots, 1.2.3) ou de carte cognitive (1.2.4). De plus, un insecte perdu pourrait revenir sur ses pas (backtracking, 1.2.3). Plusieurs projets de recherche ont permis de préciser petit à petit ces différentes stratégies.
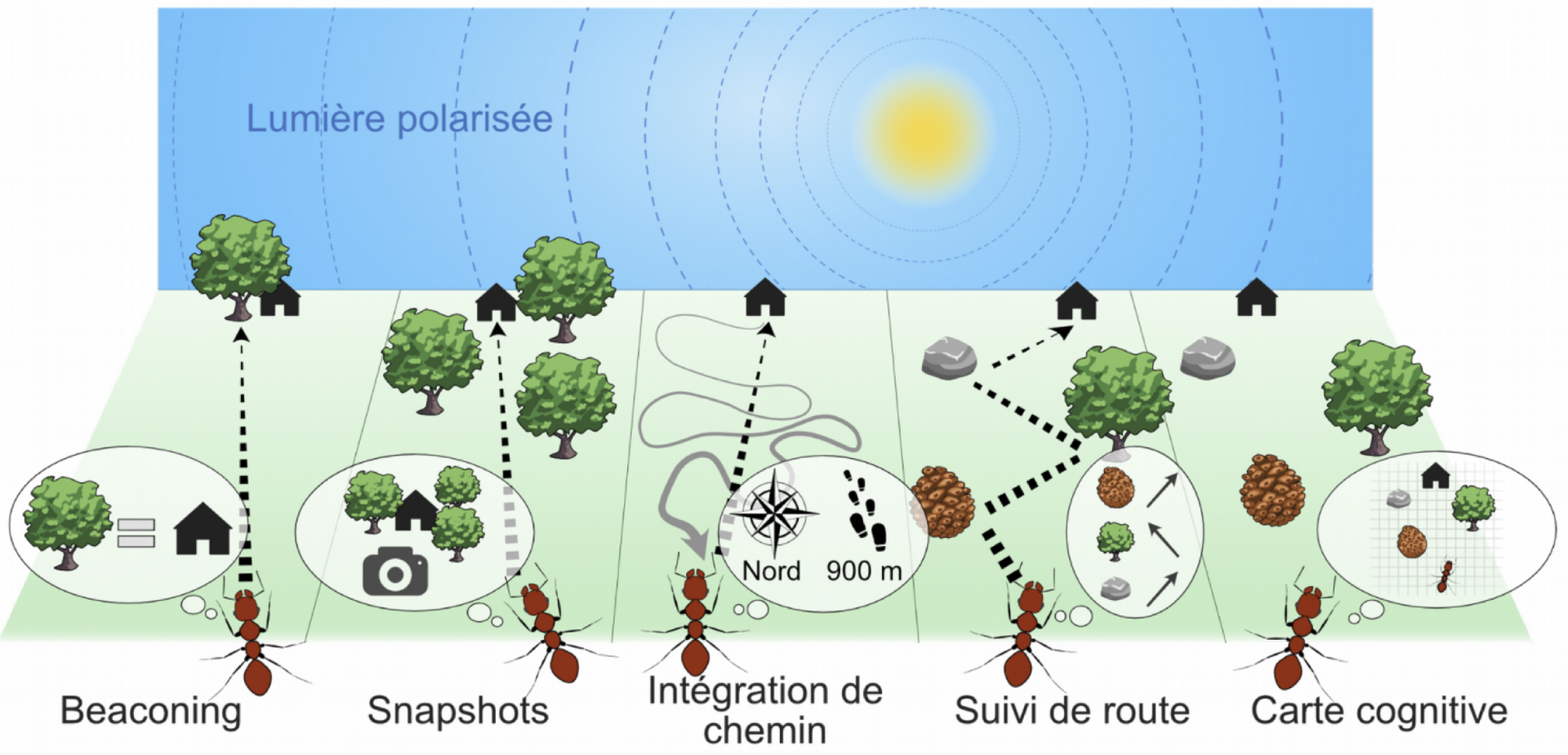
Le beaconing consiste à utiliser un élément du paysage (balise ou amer) pour s’orienter. Dans la stratégie snapshot, la scène visuelle actuelle est comparée avec une scène visuelle en mémoire. L’intégration de chemin repose sur l’enregistrement en continu de la distance et de la direction au nid. Dans la stratégie de suivi de route, une séquence d’images de référence est mémorisée. La carte cognitive consiste à avoir une représentation exocentrée de l’environnement. Comme précisé dans le texte, les stratégies sont illustrées ici de manière anthropomorphique pour faciliter la compréhension, et leur implémentation diffère chez les insectes. Figure inspirée de [10].
Guidage : beaconing et snapshot
Lorsque le nid est proche, les insectes pourraient utiliser des stratégies de guidage. La navigation basée sur un point de repère (on utilise plus couramment le terme anglais « beaconing », (5)) consiste simplement à identifier un repère visuel saillant comme un arbre (appelé landmark) et à l’associer avec le nid. Ce repère visuel est parfois nommé « amer » en référence au vocabulaire en navigation maritime. Rentrer au nid consiste alors à diminuer la distance entre la position actuelle et l’amer. Aujourd’hui, de nombreuses découvertes suggèrent que les insectes n’utilisent pas de repères spécifiques pour la navigation, mais plutôt de larges panoramas visuels (voir partie 2), c’est la stratégie snapshot. Dans cette stratégie, l’entrée du nid n’est pas associée à un unique repère visuel, mais à plusieurs repères et à leur agencement spatial relatif (le nid se situe au centre d’un cercle de pommes de pins par exemple). Pour cela, un insecte pourrait extraire et comparer les différents points de repère dans la scène visuelle actuelle avec une image panoramique de référence (instantané ou snapshot) en mémoire. Il s’alignerait alors lui-même avec celle-ci. Il n’est pas rare de voir des abeilles ou guêpes nées récemment voler en cercles autour du nid avant de partir. Ces vols d’apprentissage leur permettraient d’associer l’entrée de leur nid avec un instantané qui synthétise l’arrangement spatial relatif de plusieurs repères visuels. Dans une expérience historique, Tinbergen a démarqué l’entrée du nid d’une guêpe Philantus par un cercle de pommes de pin. Une fois la guêpe partie, le chercheur a déplacé ce cercle, et à son retour, la guêpe cherchait son nid au niveau du cercle de pommes de pin (Figure 1c) (2). On observe également des marches d’apprentissage chez de nombreuses espèces de fourmis. Bien entendu, cette stratégie suppose que l’insecte puisse voir ce repère visuel continuellement, ce qui n’est valable que lorsqu’il est à proximité de sa cible.
Intégration de chemin
Lorsque le nid est plus loin et non « visible », les abeilles peuvent utiliser une stratégie appelée intégration de chemin (aussi connue sous le nom de navigation à l’estime). Elle consiste à estimer en temps continu la distance vers le nid (grâce à un odomètre, un système permettant d’évaluer la distance parcourue), ainsi que la direction vers le nid (grâce à un compas). Les mécanismes de compas reposent sur des indices externes (voir partie 2) et sont au cœur des stratégies d’orientation en ligne droite des bousiers, ou de la migration des papillons monarques.
En intégrant continuellement la direction et la distance vers le nid, un insecte serait capable de former inconsciemment un vecteur-direction avec pour origine son point de départ, et pointant vers sa propre position. Une fourmi ou une abeille, par exemple, pourrait donc suivre ce vecteur en sens inverse pour rentrer au nid en ligne droite, même dans un environnement complètement nouveau. Cette stratégie permet notamment de revenir au nid (ou au dernier point connu) après avoir exploré un environnement non familier. Néanmoins, elle repose sur des estimations de distances et d’angles, et accumule les erreurs sur de longues distances.
Suivi de chemin, de routes et retour arrière
Avant de parler du suivi de routes, mentionnons brièvement la stratégie de suivi de chemin (trail-following). Cette stratégie concerne certaines fourmis, qui utilisent un chemin d’odeur pour revenir au nid, ou certains Hyménoptères volants qui suivent des routes goudronnées pour naviguer.
La stratégie de suivi de routes (route-following) est, comme celle de l’intégration de chemin, basée sur un référentiel égocentré, c’est-à-dire ayant pour référence l’animal lui-même. La position des éléments du paysage est définie par rapport à l’insecte – « l’arbre se situe à ma gauche » – et non par rapport aux autres éléments – « l’arbre est à droite de la maison ». Cette stratégie consiste à suivre un protocole séquentiel de déplacements à réaliser, et rappelle par sa nature les chasses aux trésors de notre enfance (« à l’arbre, je tourne à gauche et je marche jusqu’à la maison… »). De nombreuses abeilles et fourmis se retournent fréquemment pour réaliser une série d’instantanés. Ces insectes auraient en mémoire une série d’instantanés, chacun correspondant à un point temporel (6). À la différence de la stratégie snapshot, la stratégie de suivi de routes repose non pas sur un, mais sur plusieurs instantanés, et fonctionne lorsque le nid est « loin ». En réalité, de nombreuses expériences suggèrent un mécanisme un peu différent de celui de la carte au trésor décrit précédemment. Les instantanés en mémoire ne donneraient d’informations ni sur l’endroit où l’insecte se trouve, ni sur l’endroit où il doit aller ensuite. Ils donneraient plutôt une information sur la familiarité (est-ce que je suis déjà venu ici ?) : lorsqu’une fourmi retourne à son nid, une vue familière lui procurerait un sentiment positif l’incitant à continuer sa route, alors qu’une vue non familière l’inciterait au contraire à tourner jusqu’à trouver une vue familière. De plus, contrairement aux primates, les insectes navigueraient sans extraire les objets spécifiques d’une scène visuelle (un arbre, un rocher… voir partie 2.3).
Ces routes ne sont pas figées. Elles sont au contraire très flexibles et remodelées en fonction des changements de l’environnement. Les routes associées à une expérience désagréable, comme une chute ou un long détour, sont alors évitées au profit d’autres routes. Cela souligne les capacités d’adaptation de ces insectes (7).
De plus, lorsqu’un insecte se perd, il peut réaliser un retour arrière (backtracking), c’est-à-dire revenir sur ses pas pour augmenter les chances de revenir sur un territoire connu (8).
Une carte cognitive ?
Enfin, terminons par la dernière stratégie, dont l’existence chez les insectes est controversée. La navigation à la carte cognitive (ou vraie navigation) correspond à une stratégie exocentrée qui consiste à représenter mentalement les localisations de tous les indices sensoriels ainsi que les distances relatives entre elles. Un animal déplacé n’importe où dans une carte devrait être capable de retrouver son but. Si des preuves expérimentales existent chez les mammifères, la présence de cartes cognitives est encore controversée chez les insectes (9).
Navigation et indices sensoriels
Le succès des stratégies et processus de navigation décrits dans la partie précédente repose sur l’importance des points de repères (beaconing, suivi de route), des instantanés (snapshots, suivi de route), des compas (intégration de chemin, migration, marche en ligne droite) ou encore des odomètres (intégration de chemin). Nous allons détailler ce qui se cache derrière ces termes, et nous verrons que de nombreux indices sensoriels (11) permettent de les estimer (Figure 3).
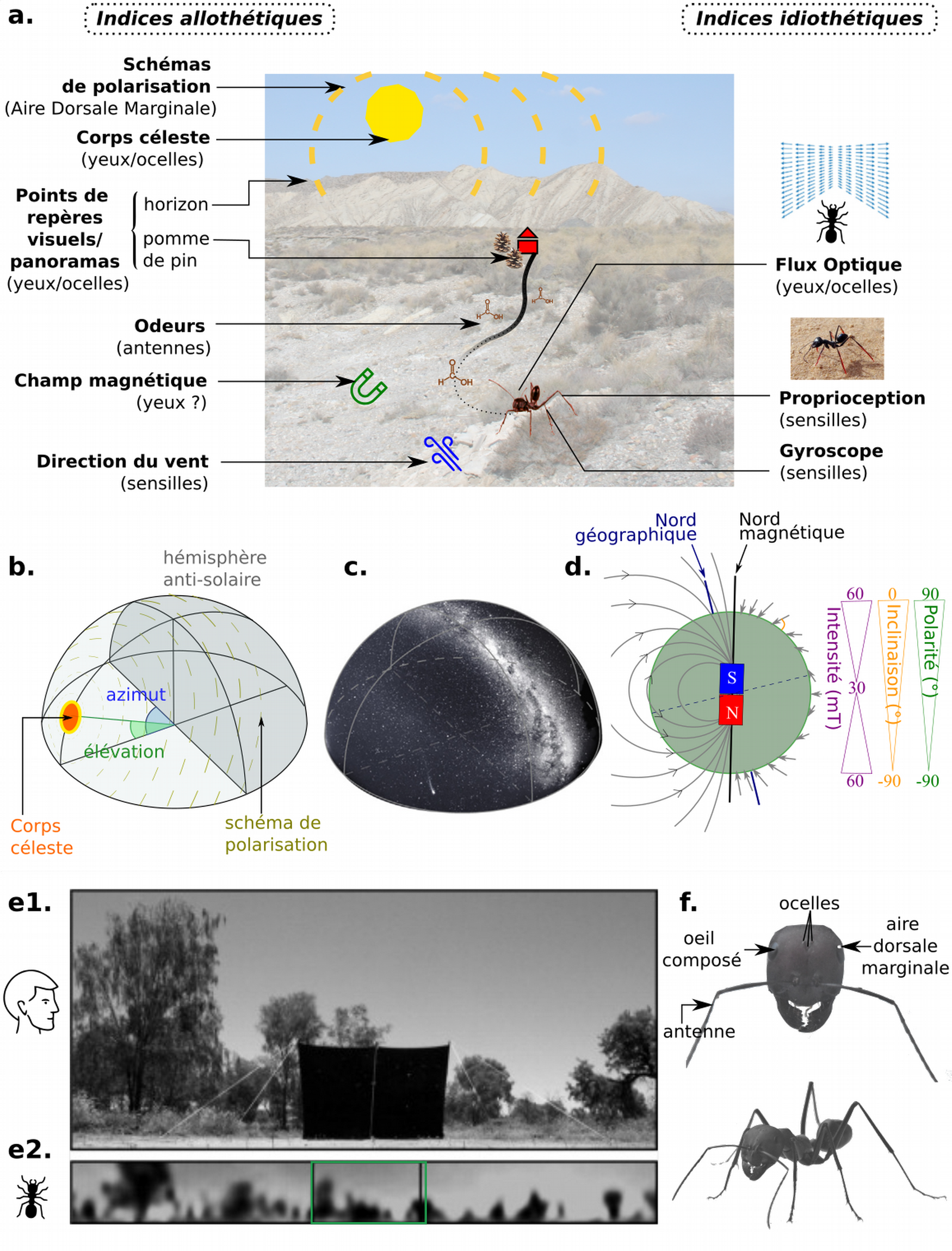
(a) Indices externes (allothétiques) et internes (idiothétiques) pouvant être utilisés pour la navigation par les insectes. (b) Indices donnés par un corps céleste : azimut, élévation, gradient d’intensité et lumière polarisée. (c) Voie Lactée. (d) Champ magnétique terrestre. (e) Perspectives visuelles différentes chez l’espèce humaine (e1) et la fourmi (e2), d’après (12). Le cadre vert sur la figure e2 (fourmi) délimite le champ visuel de l’être humain. (f) Quelques organes sensoriels d’une fourmi Cataglyphis, mentionnés dans (a).
Crédits : Fourmi avec échasses : [13] ; Désert : Auteur inconnu, CC0 Domaine public, PxHere ; Voûte céleste : redessinée d’après [11] ; Perspectives visuelles : [12] ; Fourmi Cataglyphis : Paul Clémençon, CC BY-NC.
Odomètres
L’estimation de la distance parcourue repose sur des indices idiothétiques, ou internes. Pour retrouver leur nid, les fourmis du désert ne peuvent pas suivre une ligne de phéromones comme les fourmis des milieux plus humides en raison de l’évaporation des odeurs. Il a été montré que ces fourmis pouvaient compter leur pas grâce à un pédomètre. En modifiant expérimentalement la longueur des foulées (soit en rallongeant les pattes des fourmis avec des soies de cochons – comme des échasses – soit en coupant une partie de la patte), des chercheurs ont montré que les fourmis n’arrivaient plus à estimer correctement la distance au nid (13).
Les abeilles perçoivent le mouvement grâce au flux optique, la vitesse de défilement du champ visuel pendant un déplacement (14). Enfin, la perception des accélérations grâce à des sensilles couplées à des mécanorécepteurs permet également d’évaluer le mouvement : lorsqu’un insecte se déplace, les forces mécaniques sont différentes de celles qui s’appliquent sur un insecte au repos.
Compas/Boussoles
En plus des indices idiothétiques, un compas externe (indice allothétique) est nécessaire pour permettre à l’insecte de calibrer sa direction. Cette absence de fiabilité des indices idiothétiques seuls peut se mettre en évidence par une expérience très simple. Bandez-vous les yeux et tentez de marcher en ligne droite sur une grande distance : vous finirez par marcher en cercles à cause de l’accumulation d’erreurs (15) !
De nombreux indices allothétiques peuvent servir de compas, mais la distance à cet indice est importante. Un indice idéal est situé infiniment loin, car plus un indice est loin, moins sa position relative change avec l’effet de parallaxe (la translation d’un objet par rapport à son arrière-plan, en réponse à la translation de l’observateur). Le Soleil (pour les insectes diurnes) est un indice idéal. Des expériences historiques consistant à « renverser le soleil » avec un miroir (en cachant sa vraie position avec un écran) ont montré l’importance de cet astre chez les bousiers ou encore les fourmis (16). Lorsque la position du Soleil change artificiellement de 180°, ces insectes tournent de 180° et partent dans la direction opposée. Pour les insectes nocturnes, la Lune et même la Voie Lactée (17) peuvent servir de compas. Des chercheurs ont remarqué que des bousiers placés dans un planétarium présentant un faux ciel rencontraient des difficultés à naviguer correctement.
Cependant, pour utiliser ces indices célestes, les insectes doivent résoudre deux défis. Tout d’abord, la position du Soleil, de la Lune et de la Voie Lactée change tout au long de la journée/nuit, et même au cours des saisons. Ce n’est pas un problème pour des insectes se déplaçant sur de courtes distances, mais pour les migrateurs comme le monarque, cela pourrait entraîner une déviation importante. Des expériences ont permis de montrer que les insectes peuvent apprendre la course du Soleil (18), sans que l’on en connaisse le mécanisme précis. Ils pourraient compenser ce mouvement grâce à un mécanisme circadien situé dans leurs antennes qui permettrait de mesurer le temps (19), ou peut-être utiliser le centre de rotation de la Voie Lactée (fixe) comme cela a été montré chez certains oiseaux migrateurs (20).
Le second problème posé par l’utilisation d’un repère céleste est que celui-ci peut être caché par les nuages. Certains insectes peuvent cependant déduire des indices sur la position de ces repères, par exemple le gradient chromatique ou d’intensité lumineuse (l’hémisphère solaire contient plus de vert et est légèrement plus clair que l’hémisphère anti-solaire), ou encore la lumière polarisée. La lumière non polarisée des corps célestes devient polarisée lorsqu’elle passe à travers l’atmosphère, en raison d’un processus appelé diffraction de Rayleigh. La lumière du Soleil est polarisée de manière maximale à environ 90° du Soleil, et l’angle de polarisation forme des cercles concentriques autour du Soleil. Il est alors théoriquement possible d’inférer la position du Soleil en milieu de journée grâce à ces indices. Beaucoup d’insectes possèdent des récepteurs spécifiques pour détecter cette lumière polarisée. D’autres, comme le monarque ou le bogong, utilisent des indices non célestes, comme le champ magnétique terrestre (21). Celui-ci présente l’avantage d’être relativement stable et d’être toujours disponible n’importe où sur la surface du globe (à l’exception des pôles), quelles que soient les conditions météorologiques. Chaque point du globe est caractérisé par une intensité du champ magnétique, mais aussi par un angle d’inclinaison et une polarité des lignes de champ.
À une échelle plus restreinte, des gradients d’odeurs ou encore des indices mécanosensoriels (22) comme le vent ou la pression ambiante peuvent servir de compas.
Points de repères (landmarks) et instantanés (snapshots)
Sur la figure 2, l’arbre est le point de repère utilisé par la fourmi pour retrouver son nid. En fait, les points de repères peuvent être variés : tout objet suffisamment constant et saillant peut en constituer un. Certains insectes combinent points de repères et compas (21), en s’alignant avec un point de repère géographique comme une rivière ou une montagne (stratégie probablement cognitivement plus simple et rapide), tout en actualisant la direction du compas de manière occasionnelle (stratégie probablement plus complexe cognitivement, mais plus fiable). Aujourd’hui, des études suggèrent que les insectes utilisent en fait une image panoramique de tous les indices disponibles (instantané). Un insecte se dirigerait vers un point de repère seulement en raison de son caractère saillant dans l’instantané (12).
En effet, on a longtemps pensé que la navigation des insectes se basait, comme chez les humains, sur l’extraction d’objet(s) pour réaliser ces images panoramiques. En fait, de nombreux travaux suggèrent que les insectes utilisent les scènes visuelles comme un tout sans avoir besoin d’extraire individuellement les repères visuels. Comme illustré sur la figure 3, les fourmis par exemple perçoivent le monde d’une manière bien différente de la nôtre. Ce qui apparaît comme un désavantage pourrait être en réalité un atout : la faible résolution spatiale de leurs yeux (la capacité à distinguer deux objets proches) permettrait par exemple d’encoder le panorama visuel complexe en une image simple et sans détails superflus (12,23).
La navigation, une intégration multimodale
En fonction de l’environnement et du mode de vie de l’insecte, les indices sensoriels varient, même au sein d’un groupe d’individus phylogénétiquement proches : les bousiers diurnes utilisent plutôt les corps célestes, alors que les bousiers nocturnes utilisent plutôt la lumière polarisée la nuit (24). Le cerveau de ces espèces proches a globalement la même structure et ce sont plutôt leurs connexions nerveuses fines qui expliquent ces différences. De plus, on sait qu’il n’y a souvent pas une, mais plusieurs modalités sensorielles impliquées dans la navigation (11). Cependant, leur coordination est mal comprise.
Navigation et cerveau
Chez les mammifères, la navigation repose sur le système mnésique, notamment l’hippocampe, une zone connue du grand public en raison de son importance dans les pathologies neurodégénératives. La navigation des insectes, beaucoup moins médiatisée, offre pourtant de nombreux outils de compréhension notamment par sa relative simplicité.
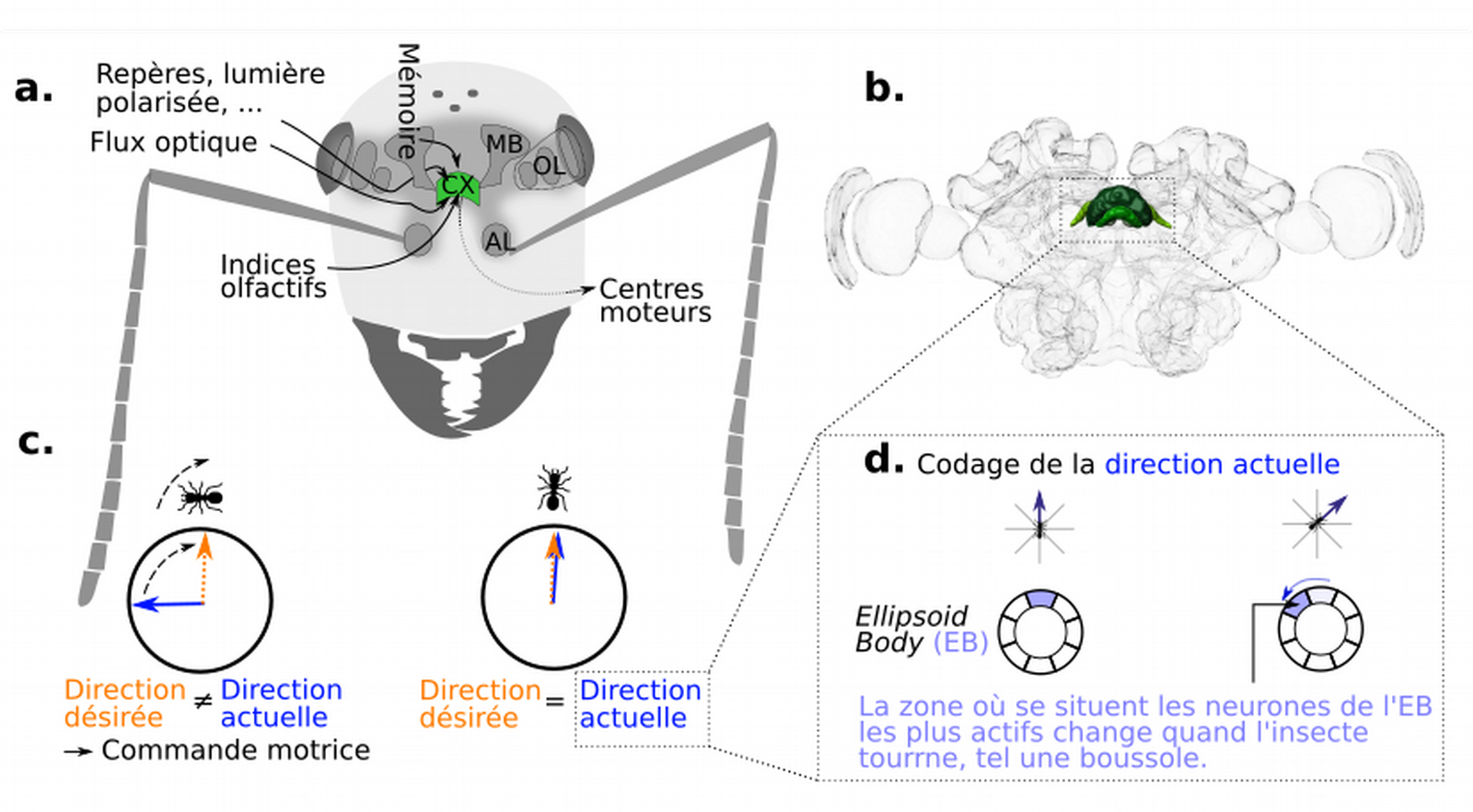
(a) Les indices sensoriels sont prétraités par des régions périphériques : lobes optiques (notés OL pour Optic Lobes) et lobes antennaires (AL, Antennal Lobes). Ces informations sont ensuite traitées dans d’autres régions, dont les corps pédonculés (MB, Mushroom Bodies) et le complexe central (CX, Central compleX), en vert (b). Les insectes compareraient le paysage observé avec le paysage en mémoire pour corriger leur trajectoire (c). Dans le complexe central, il existe un circuit neuronal qui représente la position de l’insecte par rapport à son environnement (d), c'est-à-dire la direction actuelle.
Crédits : Cerveau de fourmi : InsectBrainDatabase, CC BY-NC
Perception et prétraitement des informations sensorielles
Comme le cerveau des mammifères, celui des insectes comporte plusieurs régions, chacune associée à une ou plusieurs fonctions spécifiques (Figure 4a). Les indices sensoriels sont prétraités par des systèmes primaires, puis intégrés dans des centres supérieurs, avant de produire un unique comportement. Chez de nombreuses espèces, les systèmes sensoriels sont particulièrement sélectifs aux stimuli les plus pertinents de l’environnement pour l’animal (hypothèse du matched-filtering de Wehner) (25). Par exemple, les caractéristiques anatomiques de l’œil des fourmis permettraient d’encoder le panorama visuel complexe en une image simple mais informative.
Les systèmes primaires correspondent aux lobes antennaires, optiques et ocellaires, qui synthétisent et prétraitent l’information provenant des antennes pour l’olfaction et des yeux et ocelles pour la vision. Dans les lobes optiques, l’information visuelle est notamment synthétisée par des détecteurs élémentaires de mouvement, qui détectent la corrélation temporelle entre deux signaux et donnent une estimation du mouvement.
Les corps pédonculés (MB pour Mushroom Bodies) sont impliqués dans les tâches nécessitant une mémoire visuelle ou olfactive à long terme, comme le suivi de routes. La stratégie snapshot repose sur la mémoire d’instantanés familiers, et sur l’alignement avec ceux-ci. Les instantanés familiers seraient encodés et associés à une valence positive par les corps pédonculés (26).
Enfin, le complexe central (CX pour Central compleX) est une zone d’intégration majeure pour la navigation. Il combinerait les multiples informations sensorielles pour encoder la propre direction de l’insecte (dans quelle direction l’insecte se dirige, voir partie 3.2). Dans le complexe central, la propre direction de l’individu serait alors comparée avec la direction désirée, qui peut correspondre au vecteur-direction qui dérive du compas et d’un odomètre (stratégie d’intégration de chemin), ou encore à un instantané familier (stratégie de suivi de route, image-matching, Figure 4c). Si les directions actuelles et désirées sont différentes, le complexe central initie alors la locomotion de l’insecte en envoyant des influx nerveux aux centres moteurs. La trajectoire de l’insecte est ainsi corrigée.
Le circuit de cap suit la direction de l’individu
Mais comment la direction de l’individu est-elle encodée ? Dans le complexe central, des populations de neurones complémentaires forment des circuits complexes capables d’opérer des calculs vectoriels essentiels à la représentation spatiale. Il existe notamment une zone appelée corps ellipsoïde qui ressemble à un anneau et qui enregistre tout changement de direction dans un plan en deux (et peut-être même en trois) dimensions.
Au sein de cette zone, une population de neurones innerve des sections particulières appelées paquets (bundles), de taille régulière et qui se succèdent le long de l’anneau. L’activité de chacun de ces paquets de neurones est corrélée à une direction de l’insecte avec une résolution de 45°.
Lorsque l’insecte tourne à 45°, c’est un nouvel ensemble de neurones, adjacent au précédent, qui devient actif. Cela rappelle l’aiguille d’une boussole qui tourne de 45° lorsque l’on tourne sur nous-mêmes de 45°. Ensemble, ces huit paquets de neurones représentent l’environnement de l’insecte à 360°. L’encodage de la direction dépend à la fois d’indices internes (le système fonctionne dans le noir complet) et externes qui sont intégrés pour former un signal unique. Lorsque l’on immobilise une drosophile sur un support, et que l’on tourne artificiellement une scène visuelle, la zone du corps ellipsoïde activée tourne aussi, démontrant ainsi que le signal dépend d’indices externes en priorité (Figure 4d) (27). La robustesse de ce système repose sur un schéma de connectivité très précis des neurones, qui est conservé chez un grand nombre d’espèces d’insectes (11), suggérant son importance.
La représentation de la direction désirée et son intégration avec le circuit de cap sont moins bien comprises, mais on peut raisonnablement espérer des avancées étant donné certains avantages expérimentaux inhérents aux insectes, comme la capacité d’identifier et d’enregistrer des neurones spécifiques ou la faible variance structurale entre les individus (28).
Navigation, pollution lumineuse et robotique
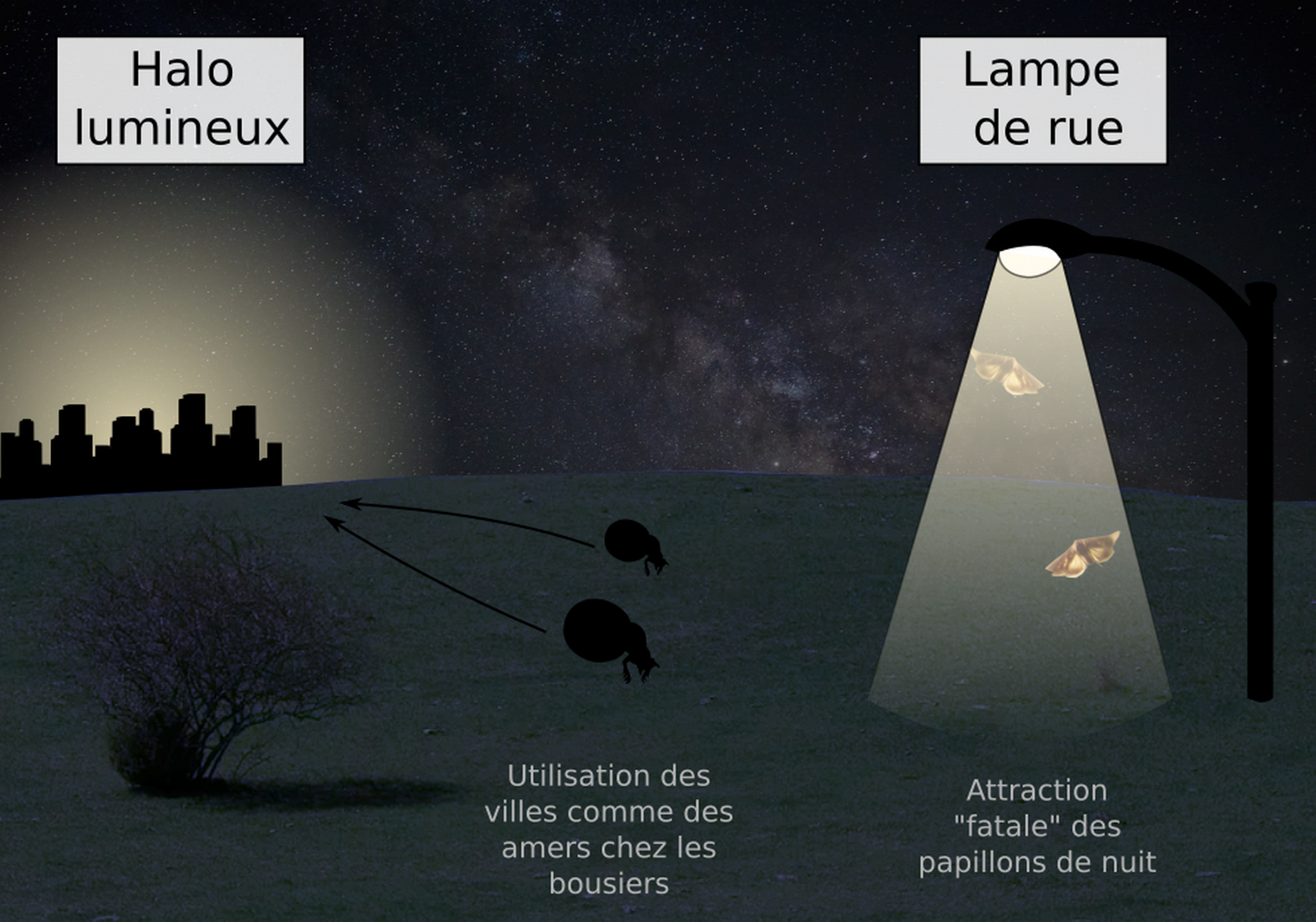
La pollution lumineuse a des conséquences importantes sur la navigation spatiale des insectes. Par exemple, il est connu que les papillons de nuit sont attirés par la lumière. Pour voler en ligne droite, un papillon doit voler à un angle constant relativement à une source de lumière céleste. Cependant, s’il identifie une lampe de rue comme un corps céleste, le papillon finit par faire des spirales autour de cette lampe et par mourir d’épuisement.
Par ailleurs, dans les aires urbaines, la Voie Lactée est masquée par la pollution lumineuse. Au lieu de se disperser, les bousiers marchent en ligne droite vers la lumière (beaconing), ce qui augmente la compétition entre ces insectes (30). La pollution lumineuse pourrait donc grandement affecter l’écologie de nombreux migrateurs, ainsi que les chaînes trophiques qui en dépendent. Par exemple, la migration des bogongs en Australie représente une source de lipides importante pour de nombreux animaux des Alpes australiennes, et le déclin des populations de bogongs pourrait entraîner des conséquences en chaîne.
Les insectes intéressent les ingénieurs par leur capacité à combiner expérience passée et entrées sensorielles pour coordonner un mouvement dans un environnement dynamique et complexe (31). Des robots inspirés des insectes pourraient naviguer en milieu complexe pour localiser des victimes de catastrophe naturelle par exemple. La compréhension du réseau de neurones sous-jacent à l’intégration de chemin pourrait permettre aux robots de retourner à leur position initiale après des trajets d’exploration sans demander de calculs trop complexes. Cela limiterait ainsi leur consommation énergétique, un des points limitants des robots autonomes (28).
Conclusion
La diversité des stratégies de navigation chez les insectes soulève de nombreuses questions quant à leurs mécanismes. Elles pourraient trouver des réponses grâce aux progrès des techniques d’imagerie et aux avantages expérimentaux inhérents aux insectes. Comprendre les mécanismes de la navigation pourrait permettre d’anticiper la réponse aux changements globaux, et de développer de nouveaux algorithmes de navigation autonome.
Références
1. R. Grob, B. el Jundi, P. N. Fleischmann, Towards a common terminology for arthropod spatial orientation. Ethol. Ecol. Evol. 33, 338–358 (2021).
2. N. Tinbergen, On the Orientation of the Digger Wasp Philanthus triangulum. Fabr Zs Uber Vergl Physiol, 305–334 (1932).
3. L. Khaldy, C. Tocco, M. Byrne, E. Baird, M. Dacke, Straight-line orientation in the woodland-living beetle Sisyphus fasciculatus. J. Comp. Physiol. A 206, 327–335 (2020).
4. A. Honkanen, A. Adden, J. da Silva Freitas, S. Heinze, The insect central complex and the neural basis of navigational strategies. J. Exp. Biol. 222, jeb188854 (2019).
5. S. Healy (ed.). Spatial representation in animals (Oxford University Press, 1998).
6. B. A. Cartwright, T. S. Collett, How honey bees use landmarks to guide their return to a food source. Nature 295, 560–564 (1982).
7. A. Wystrach, C. Buehlmann, S. Schwarz, K. Cheng, P. Graham, Rapid Aversive and Memory Trace Learning during Route Navigation in Desert Ants. Curr. Biol. 30, 1927-1933.e2 (2020).
8. A. Wystrach, S. Schwarz, A. Baniel, K. Cheng, Backtracking behaviour in lost ants : an additional strategy in their navigational toolkit. Proc. Biol. Sci. 280, 20131677 (2013).
9. M. Collett, L. Chittka, T. S. Collett, Spatial Memory in Insect Navigation. Curr. Biol. 23, R789–R800 (2013).
10. C. Doussot, Leaving and returning home. Visually-guided homing in Bombus terrestris (2020) https:/doi.org/10.4119/unibi/2944818 (September 4, 2022).
11. S. Heinze, Unraveling the neural basis of insect navigation. Curr. Opin. Insect Sci. 24, 58–67 (2017).
12. A. Wystrach, P. Graham, What can we learn from studies of insect navigation ? Anim. Behav. 84, 13–20 (2012).
13. M. Wittlinger, R. Wehner, H. Wolf, The Ant Odometer : Stepping on Stilts and Stumps. Science 312, 1965–1967 (2006).
14. M. Lindauer, H. Heran, Windkompensation und Seitenwindkorrektur der Bienen beim Flug über Wasser. Z. Ffir Vgl. Physiol., 39–55 (1963).
15. J. L. Souman, I. Frissen, M. N. Sreenivasa, M. O. Ernst, Walking Straight into Circles. Curr. Biol. 19, 1538–1542 (2009).
16. F. Santchi, Obervations et remarques critiques sur le mecanisme de l’orientation chez les fourmis. Rev Suisse Zool 19, 303–338 (1911).
17. M. Dacke, E. Baird, M. Byrne, C. H. Scholtz, E. J. Warrant, Dung Beetles Use the Milky Way for Orientation. Curr. Biol. 23, 298–300 (2013).
18. M. Lindauer, Angeborene und erlernte Komponenten in der Sonnenorientierung der Bienen Bemerkungen und Versuche zu einer Mitteilung von Kalmus. Z. Für Vgl. Physiol. 42, 43–62 (1959).
19. C. Merlin, R. J. Gegear, S. M. Reppert, Antennal circadian clocks coordinate sun compass orientation in migratory monarch butterflies. Science 325, 1700–1704 (2009).
20. S. T. Emlen, Migratory Orientation in the Indigo Bunting, Passerina cyanea. Part II : Mechanism of Celestial Orientation. The Auk 84, 463–489 (1967).
21. D. Dreyer, et al., The Earth’s Magnetic Field and Visual Landmarks Steer Migratory Flight Behavior in the Nocturnal Australian Bogong Moth. Curr. Biol. 28, 2160-2166.e5 (2018).
22. M. Dacke, et al., Multimodal cue integration in the dung beetle compass. Proc. Natl. Acad. Sci. 116, 14248–14253 (2019).
23. F. Le Moël, “Navigation visuelle chez la fourmi : approche modélisatrice,” Université de Toulouse, Université Toulouse III – Paul Sabatier. (2021) (September 4, 2022).
24. B. el Jundi, et al., Neural coding underlying the cue preference for celestial orientation. Proc. Natl. Acad. Sci. 112, 11395–11400 (2015).
25. R. Wehner, ‘Matched filters’ – neural models of the external world. J. Comp. Physiol. A 161, 511–531 (1987).
26. C. Buehlmann, et al., Mushroom Bodies Are Required for Learned Visual Navigation, but Not for Innate Visual Behavior, in Ants. Curr. Biol. CB 30, 3438-3443.e2 (2020).
27. J. D. Seelig, V. Jayaraman, Neural dynamics for landmark orientation and angular path integration. Nature 521, 186–191 (2015).
28. T. Dalgaty, E. Vianello, B. De Salvo, J. Casas, Insect-inspired neuromorphic computing. Curr. Opin. Insect Sci. 30, 59–66 (2018).
29. A. Owens, et al., “Light Pollution Is a Driver of Insect Declines” (Social Science Research Network, 2019) https:/doi.org/10.2139/ssrn.3378835 (April 4, 2022).
30. J. J. Foster, et al., Light pollution forces a change in dung beetle orientation behavior. Curr. Biol. 31, 3935-3942.e3 (2021).
31. J. Dupeyroux, J. R. Serres, S. Viollet, AntBot : A six-legged walking robot able to home like desert ants in outdoor environments. Sci. Robot. 4, eaau0307 (2019).




{kind=link}
{kind=link}
.jpeg){kind=link}
{kind=link}