Des molécules thérapeutiques qui ciblent les protéines et entraînent leur dégradation.
Dans nos cellules, les protéines sont continuellement synthétisées et assurent une multitude de fonctions permettant la vie. Lorsque les protéines sont endommagées, mal repliées ou devenues inutiles, elles sont dégradées et les acides aminés qui les composaient sont recyclés. La dégradation des protéines est principalement assurée par le système ubiquitine-protéasome, dans lequel les protéines sont marquées par une chaîne de molécules d’ubiquitine, fixées de manière covalente, ce qui conduit à la dégradation de la protéine ciblée. Le protéasome reconnaît cette chaîne d’ubiquitine et dégrade la protéine étiquetée en courts fragments peptidiques.
L’ubiquitinylation des protéines est un processus en plusieurs étapes soigneusement contrôlées. Tout d’abord, les enzymes E1 activent l’ubiquitine et la transfèrent aux enzymes de conjugaison de l’ubiquitine E2. Les ubiquitine-ligases E3 rapprochent ensuite l’enzyme E2 porteuse d’ubiquitine et la protéine cible, permettant à cette dernière d’être ubiquitinylée et ainsi marquée pour sa destruction par le protéasome. La découverte de cette voie, récompensée par le prix Nobel de chimie en 2004, a donné naissance à une nouvelle classe de molécules à usages thérapeutiques visant à détruire sélectivement des protéines à l’origine de maladies.
Le thalidomide : un pionnier des dégradeurs par colle moléculaire
L’une des plus importantes percées dans ce domaine a commencé par une tragédie. À la fin des années 1950 et au début des années 1960, le thalidomide était un médicament largement prescrit pour traiter les nausées matinales chez les femmes enceintes. Ce n’est qu’après que des milliers d’enfants dans le monde soient nés avec des malformations congénitales, en particulier du développement des membres, que ce dernier a été reconnu comme un puissant tératogène humain. Malgré sa triste notoriété, le thalidomide a encore été prescrit dans quelques cas ciblés en tant que sédatif, ce qui a permis aux chercheurs de découvrir que cette molécule et ses dérivés étaient remarquablement efficaces pour traiter certaines maladies, comme la lèpre et le myélome multiple. Ce n’est cependant qu’en 2010 que le mécanisme d’action du thalidomide a été compris.
Ce dernier agit en se liant à une protéine appelée cereblon, qui fait partie du complexe ubiquitine-ligase E3. Dans les cellules saines, la protéine cereblon (CRBN) cible des protéines portant, en position C-terminale, des modifications cycliques imides qui résultent de dommages ou de modifications enzymatiques. Cependant, lorsqu’elle est liée au thalidomide, la protéine cereblon présente un fonctionnement altéré qui entraîne l’ubiquitinylation d’un sous-ensemble complètement différent de protéines, comprenant les facteurs de transcription à doigts de zinc Ikaros (IKZF1) et Aiolos (IKZF3) 1. La destruction de ces nouvelles cibles est responsable à la fois des effets tératogènes et thérapeutiques du thalidomide.
Le thalidomide et ses analogues, incluant le lénalidomide et le pomalidomide, sont les premiers membres d’une classe de médicaments désormais connue sous le nom de dégradeurs par colle moléculaire. De manière générale, ces médicaments agissent en se liant fortement au domaine de liaison au substrat d’une ligase E3, ce qui bloque à la fois la liaison des substrats normaux et présente une interface de liaison modifiée capable de reconnaître de nouveaux substrats 1. Sur la figure 1, la protéine cereblon (CRBN) est représentée liée à l’analogue du thalidomide, le pomalidomide, et au facteur de transcription Ikaros 2 (entrée de la base de données des protéines (PDB) 6h0f et 4a0k).
Le fait de découvrir qu’une petite molécule pouvait « reprogrammer » le système cellulaire de dégradation des protéines a ouvert de nouveaux horizons pour la découverte de médicaments. Aujourd’hui, plus de vingt dégradeurs par colle moléculaire sont en phase d’essais cliniques, plusieurs ayant atteint la phase III pour le traitement de cancers, notamment le myélome multiple et le lymphome à cellules B.
PROTAC : une approche modulaire de la destruction des protéines
Bien que les dégradeurs par colle moléculaire puissent être des médicaments efficaces, la conception rationnelle de molécules ciblant spécifiquement une protéine d’intérêt s’est avérée difficile. Proposés pour la première fois en 2001, les PROTAC (abréviation en anglais de PROteolysis TArgeting Chimeras, ou chimères ciblant la protéolyse en français) utilisent une approche beaucoup plus favorable à la conception de nouveaux médicaments. Les PROTAC sont créés comme des molécules à deux têtes, avec une extrémité se liant spécifiquement à une protéine cible d’intérêt, et l’autre extrémité se liant à une ubiquitine-ligase E3. Un court connecteur relie les deux. En attachant une protéine cible à une ubiquitine-ligase E3, les PROTAC peuvent diriger son ubiquitinylation et, par la suite, sa dégradation.
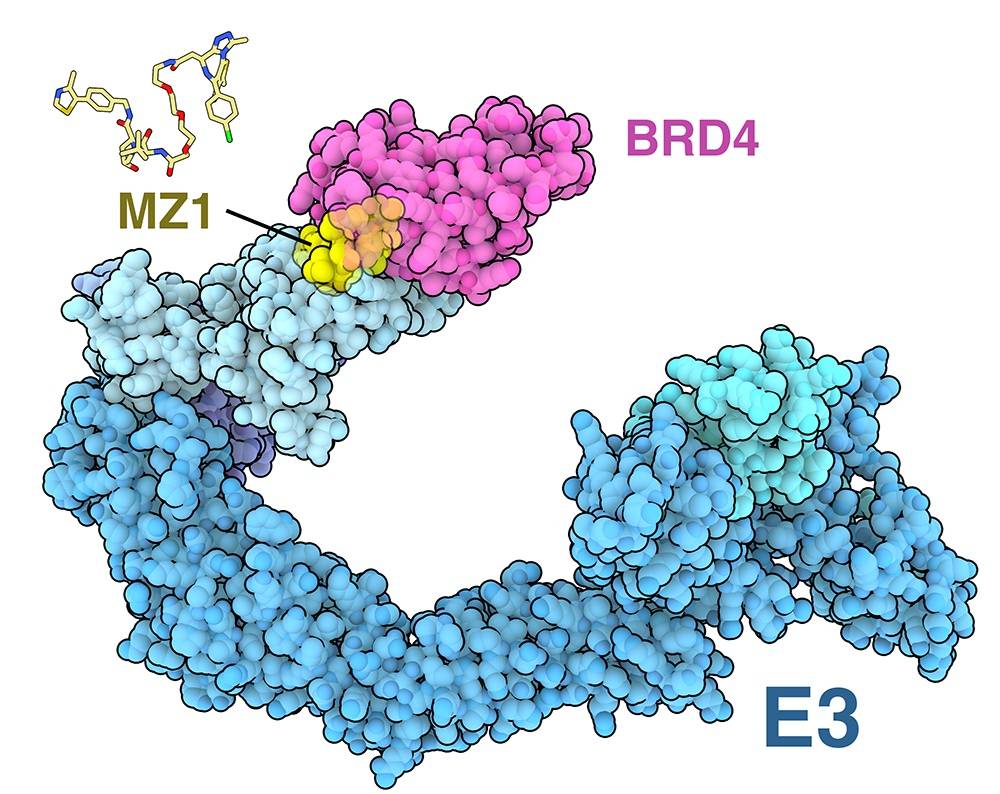
Les premières versions des PROTAC étaient basées sur des protéines, en utilisant de courtes séquences peptidiques pour recruter à la fois les ubiquitine-ligases E3 et la protéine cible. Actuellement, les PROTAC sont largement conçues à partir de petits ligands reliés par un connecteur chimique. Par exemple, le PROTAC MZ1 (représenté en jaune sur la figure 2) cible à la fois BRD4 (en rose), un régulateur transcriptionnel qui joue un rôle dans certains cancers et maladies d’origine immunitaire, et la protéine von Hippel-Lindau du complexe ubiquitine-ligase E3 1 (en bleu ; entrées PDB 5t35 et 5n4w) 2.
La conception « prête à l’emploi » rend les PROTAC fortement adaptables à une large gamme de cibles thérapeutiques potentielles. Il existe actuellement plus de 50 PROTAC en essais cliniques dans le monde, incluant plusieurs essais de phase III ciblant les cancers du sein et de la prostate.
Les dégradeurs par colle moléculaire peuvent altérer la conformation des ligases E3
La protéine cereblon peut adopter différentes conformations : une première conformation ouverte (entrée PDB 8cvp) et une autre fermée, nécessaire à la liaison au substrat et à la dégradation. Des recherches récentes ont montré que les dégradeurs par colle moléculaire stabilisent la conformation fermée (entrée PDB 8d7u), et ce de manière d’autant plus efficace que les dégradeurs sont puissants (comme le mezigdomide, un dérivé du thalidomide) 1. La conséquence est une ubiquitinylation et une dégradation accrues de la protéine cible Ikaros (entrée PDB 8d7z), ainsi qu’un effet thérapeutique plus important. Vous pouvez examiner de plus près les effets des dégradeurs par colle moléculaire en vous rendant sur la page de l’article d’origine (en anglais), et en cliquant sur l’onglet JSmol de la section Exploring the structure.
Pour aller plus loin
- Vous pouvez en apprendre davantage sur l’ubiquitine et la machinerie d’ubiquitinylation ainsi que sur le protéasome dans d’anciens articles de la molécule du mois, ainsi que dans un article sur le site Planet-Vie : La voie de dégradation ubiquitine dépendante.
- Vous pouvez découvrir le fonctionnement de l’auxine, une molécule qui est naturellement un dégradeur par colle moléculaire qui joue un rôle important dans le développement des plantes.
- Le facteur induit par l’hypoxie α (HIF-α) est un exemple de facteur de transcription pouvant être rapidement dégradé via la voie ubiquitine-protéasome.
Ce texte correspond à la traduction par Cédric Bordi de l’article Molecule of the Month : PROTACs and molecular glue degraders 1 écrit par Janet Iwasa en mai 2026 sur le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB).
Crédits

Cet article est publié en partenariat avec le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB).