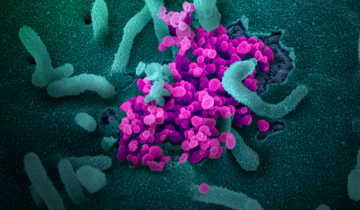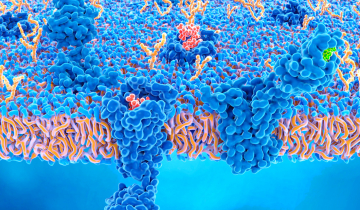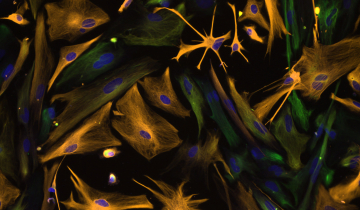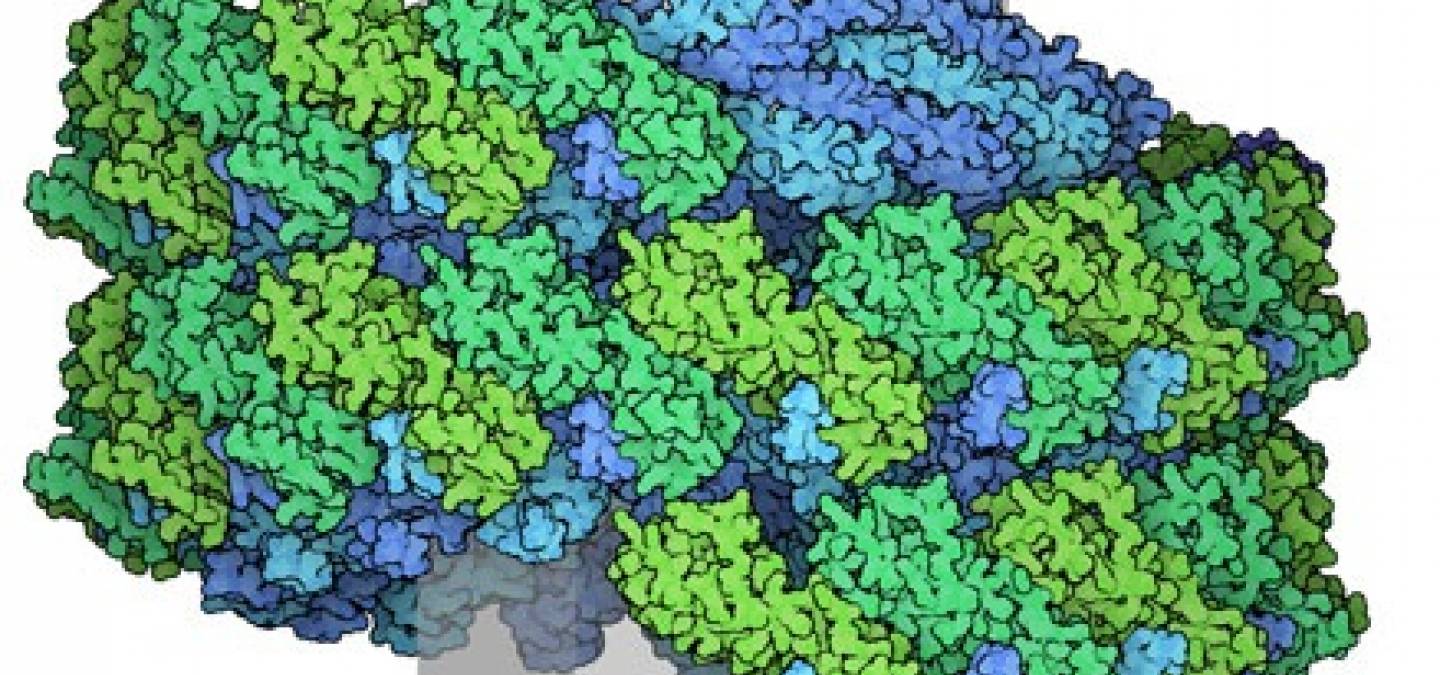
Le complexe ESCRT-III forme des assemblages hélicoïdaux qui remodèlent les membranes cellulaires.
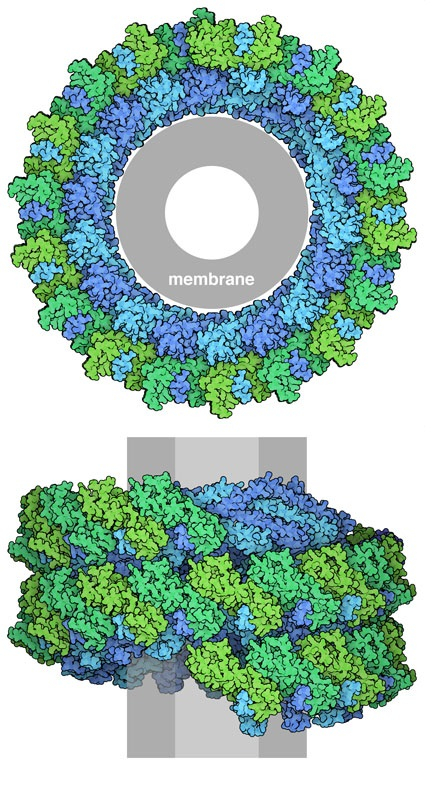
La membrane est représentée en gris.
Les membranes sont comme des murs qui structurent nos cellules, formant une barrière protectrice autour de chacune d’entre elles et délimitant des compartiments à l’intérieur de celles-ci, comme le noyau. Cependant, les membranes ne sont pas fixes comme les murs de nos maisons. Les cellules modifient constamment leurs membranes en fonction de leurs besoins, en façonnant les différents compartiments et en contrôlant le trafic vésiculaire entre ces derniers. Les protéines ESCRT contribuent à ce processus, en remodelant les membranes et en pinçant des parties qui forment des vésicules. Ce rôle se reflète dans leur nom de « complexes de tri endosomal requis pour le transport » (Endosomal Sorting Complexes Required for Transport en anglais). Le complexe ESCRT-III joue un rôle central en fournissant la force permettant de remodeler les membranes cellulaires.
Protéines étranglantes
Nos cellules construisent plusieurs types de protéines ESCRT-III avec différentes fonctions. Celles représentées sur la figure 1 (entrée 6tz4 de la banque de données sur les protéines (PDB)) s’assemblent en hélices ou en spirales qui forment un collier autour d’une section tubulaire d’une membrane 1. Ce grand assemblage associe deux protéines : CHMP1B et IST1. Ensemble, elles forment au centre un rétrécissement qui étrangle la membrane. Ce processus est utilisé, par exemple, lors du bourgeonnement de petites vésicules à partir d’un endosome plus grand 2.
À l’envers
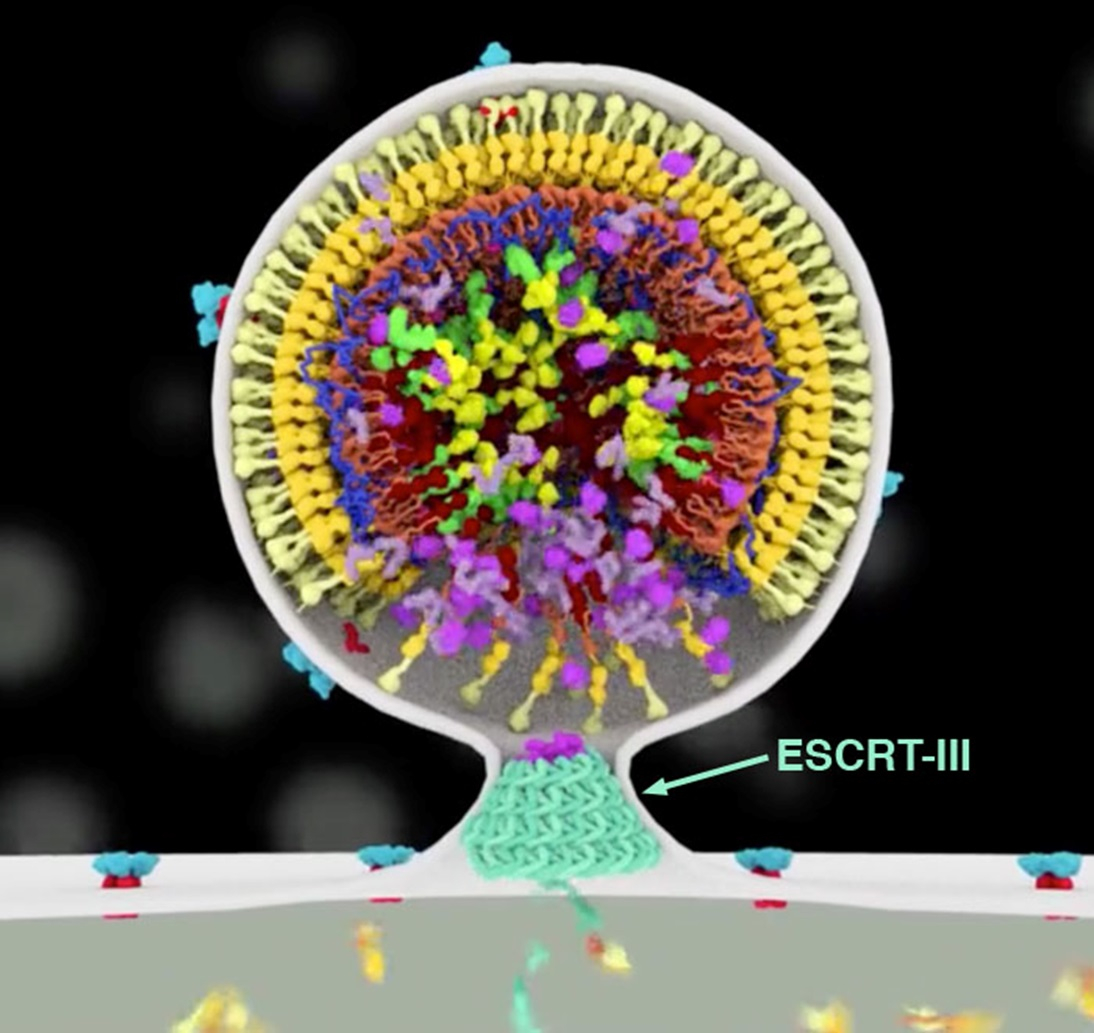
Étonnamment, de nombreux types d’ESCRT-III fonctionnent de manière opposée, en étant situé non pas autour des membranes, mais à l’intérieur. Ces hélices protéiques tirent alors sur les membranes, qui les recouvrent à la manière d’un manchon. De cette manière, elles resserrent les membranes de l’intérieur, comme illustré sur la figure 2. Ce mécanisme joue plusieurs rôles essentiels dans nos cellules. Par exemple, lorsqu’une cellule se divise, le complexe ESCRT-III aide à rétrécir le sillon de division, permettant aux deux cellules filles de se séparer. De même, ESCRT-III scelle les trous dans les membranes et aide à réparer les lésions subcellulaires. Ce complexe joue également un rôle important dans l’organisation des formes membranaires spécifiques de certaines cellules, comme les longues extensions des cils et les structures ramifiées des cellules nerveuses.
Aides détournées
Le VIH-1 détourne le système ESCRT et l’oblige à participer aux dernières étapes du cycle viral. Lorsque le virus bourgeonne à la surface de la cellule infectée, ESCRT-III réduit la portion de membrane qui relie le virus à la cellule, ce qui permet au virus de se détacher et de s’échapper. Comme le montre la figure 2, issue d’une animation de Janet Iwasa, le complexe ESCRT-III formerait une spirale qui rétrécirait progressivement la membrane. Pour voir l’ensemble du processus, il est possible de consulter l’animation complète du cycle de vie du VIH-1.
Façonner la photosynthèse
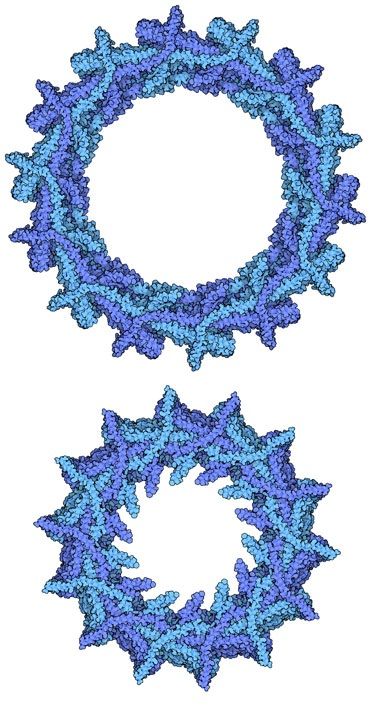
Des protéines similaires à ESCRT-III sont utilisées dans de nombreux types cellulaires et façonnent leurs membranes. Le complexe VIPP1, représenté sur la figure 3, contribue à organiser les complexes photosynthétiques des thylakoïdes chez une cyanobactérie 1. Des protéines homologues jouent probablement le même rôle dans les algues vertes et les plantes 2. Deux structures du complexe sont représentées sur la figure 3 (entrées PDB 7o3w et 7o3z), montrant comment VIPP1 forme des assemblages hélicoïdaux de différents diamètres en se resserrant autour d’un tubule membranaire 3.
Les conformations du complexe ESCRT-III
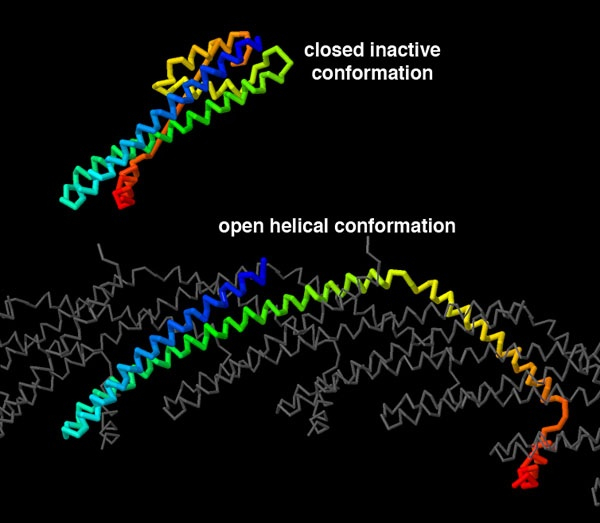
Les protéines du complexe ESCRT-III sont très dynamiques, adoptant différentes conformations lorsqu’elles se lient aux membranes en formant des hélices de plus en plus petites. Par exemple, dans le cytoplasme, la protéine CHMP3 du complexe ESCRT-III adopte une forme fermée, inactive, comme représentée sur la figure 4 1 (entrée PDB 3frt). Avec l’aide d’autres protéines ESCRT, CHMP3 s’ouvre et se lie à CHMP2A pour former l’assemblage hélicoïdal qui resserre les membranes 2, comme représentée sur la figure 4 également (entrée PDB 7zcg). Pour comparer ces deux structures, vous pouvez utiliser par exemple le logiciel de visualisation de molécules disponible sur le site PDB-101.
Ce texte correspond à la traduction par Cédric Bordi de l’article Molecule of the Month : ESCRT III 1 écrit par David S. Goodsell et paru en août 2024 sur le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB).
Crédits

Cet article est publié en partenariat avec le site PDB-101, le portail éducatif de la base de données sur les protéines (PDB).