

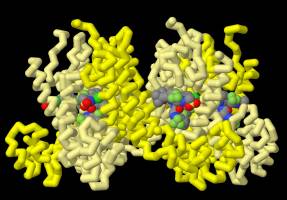
Le génome du VIH étant formé d’ARN à polarité positive, il peut être directement traduit en protéines par les ribosomes de la cellule infectée. Les ARN peuvent également provenir de la transcription du génome viral, après son intégration dans le génome de l’hôte. Trois cadres de lecture peuvent être suivis par les ribosomes, ce qui mène à la synthèse des différentes protéines virales. Le 5’LTR est suivi des gènes gag (codant les protéines de structure : matrice p17, capside p24, nucléocapside p7/p6), pol (codant les protéines virales enzymatiques : protéase p10, transcriptase inverse domaine p51/domaine RNase p15, intégrase p31) et env (codant les protéines de l’enveloppe gp120 et gp41), ce dernier étant entouré des gènes accessoires et de régulation. Les gènes de régulation tat et rev sont chacun constitués de deux exons répartis sur deux cadres de lectures différents. Pour tat, après épissage, les exons restent sur deux cadres de lecture différents. Lors de la traduction, les ribosomes changent de cadre de lecture à la jonction entre l’exon 1 et l’exon 2 (frameshifting). Pour rev, le processus d’épissage permet de relier les exons sur un même cadre de lecture.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}