

Présentation des modalités de germination de quatre types de graines : pois, blé, maïs et soja. Description des plantules.
Introduction
Les jeunes plantules constituent un matériel expérimental facile à obtenir en un temps court (quelques jours) de manière très reproductible. Pour bien analyser les réponses expérimentales de ces organismes, il est important de bien connaître leur morphologie et les caractéristiques de leur morphogenèse naturelle. Quatre espèces bien caractéristiques sont étudiées ici, deux dicotylédones (le pois et le soja) et deux monocotylédones (le blé et le maïs).
 |
 |
 |
 |
| Pois | Blé | Maïs | Soja |
Pour chacune de ces plantules, on essaiera de soulever quelques problèmes intéressant l’expérimentateur.
Voir aussi sur ce sujet le dossier Plantules modèles
La germination du pois
Trois étapes de la germination d’une graine de petit pois. La germination au sens strict débute par une imbibition et se termine par la sortie de la radicule. La suite (germination au sens large) peut être considérée comme de la croissance. On observe successivement la croissance de la jeune racine, puis celle de la jeune tige. Celle-ci s’élève au-dessus des cotylédons de la graine : il s’agit d’une tige épicotylée. Le reste de la graine, et en particulier les cotylédons, ne se soulèvent pas. Ils restent à la surface ou sous la surface de la terre : il s’agit d’une germination hypogée.
 |
 |
 |
L’ouverture de la graine au début de la germination montre clairement que les cotylédons sont en relation directe avec l’axe embryonnaire (fig. 3). Ils font partie intégrante de l’embryon. Les réserves sont situées, dans ce cas, dans les cotylédons donc dans l’embryon lui-même. Il n’y a pas d’albumen chez la graine mature : la graine est dite exalbuminée.
 |
L’observation du système racinaire d’une plantule plus âgée montre que la radicule est devenue une racine principale qui grandit verticalement vers le bas (gravitropisme positif).
 |
|
Le long de cette racine se développent des racines secondaires (ou de deuxième ordre) qui grandissent horizontalement ou selon un certain angle avec la verticale. On verra que ce n’est pas toujours le cas (monocotylédones). Ces racines apparaissent à une distance assez précise et constante de l’extrémité puis grandissent au fur et à mesure que la racine principale grandit. |
La germination du blé
Le grain de blé est un caryopse c’est-à-dire un akène (fruit sec indéhiscent contenant une graine) particulier caractéristique des graminées. Son enveloppe protectrice est constituée de deux parties, le péricarpe du fruit et le tégument de la graine. Ces deux parties sont plus ou moins soudées. Cependant, la graine constituant la majeure partie du caryopse, il est possible d’expérimenter sur cet organe comme si c’était une graine.
 |
 |
 |
| On observe la sortie d'une première racine et d'un organe aérien (en vert) le coléoptile. Très rapidement le coléoptile grandit verticalement et de nouvelles racines apparaissent. Elles sont ici en nombre de trois. | ||
Des agrandissements de détail permettent de mieux comprendre la formation de ces racines.
 |
 |
| La première racine était protégée par un étui, le coléorhize, qu'elle a rapidement percé. Les deux autres racines ne sont pas des racines secondaires apparues sur la racine principale mais des racines adventives initiées dans le corps de l'axe embryonnaire. |
 |
 |
Le coléoptile est un étui qui protège l’apex caulinaire et les jeunes feuilles. À partir d’un certain stade, sa croissance résulte essentiellement d’une croissance par élongation des cellules (auxèse). C’est cette propriété qui en fait un matériel de choix pour l’expérimentation sur la croissance cellulaire (de Darwin à Went et Thimann). Lorsque sa croissance est terminée, les jeunes feuilles le perce au niveau d’une ouverture préétablie.
La germination du maïs
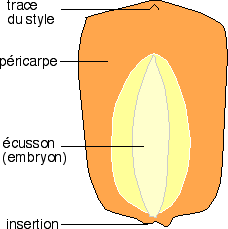
Le grain de maïs (graminées) est un caryopse comme le grain de blé.
 |
 |
 |
|
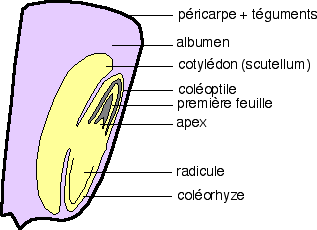
Une coupe longitudinale sagittale montre l’organisation générale d’un caryopse. Une coloration par le lugol montre que les réserves amylacées sont localisées dans un tissu spécial indépendant de l’embryon, l’albumen. La graine de ce caryopse est dite albuminée. |
La germination du grain de maïs suit les mêmes règles. Comme il est plus gros, il se prête mieux à certaines expérimentations. Sa morphogenèse présente quelques particularités.
 |
 |
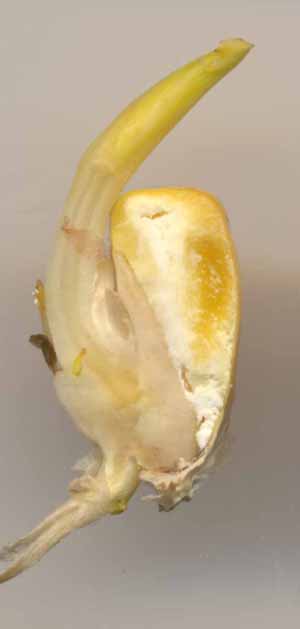
La germination à la lumière ressemble à celle du grain de blé. la germination à l’obscurité fait apparaître une particularité à savoir l’existence d’un organe tubulaire entre le corps de l’embryon et l’insertion du coléoptile : le mésocotyle.
 |
 |
| A gauche, on observe très nettement le coléorhize percé par la radicule. A droite, le mésocotyle, qui soulève le coléoptile (et donc l'apex et les jeunes feuilles contenus dans le coléoptile) est bien délimité. On constate que de nouvelles racines adventives apparaissent de manière assez irrégulières à partir de l'axe embryonnaire mais non à partir de la racine comme dans le cas du pois. |
Il n’y a donc pas un système racinaire pivotant constitué d’une racine principale et de racines de deuxième ordre mais un système racinaire fasciculé formé de racines adventives de même ordre.
Des sections longitudinales sagittales à divers stades de la germination permettent de bien montrer les relations entre les différentes parties.
 |
 |
La germination du soja
Le soja vert, utilisé dans l’alimentation asiatique sous forme de graines ou de germinations étiolées, représente un excellent matériel expérimental pour l’étude de la croissance cellulaire. En effet, comme les coléoptiles, à partir d’un certain stade la croissance de son hypocotyle est principalement due à de l’auxèse. Comme on le voit sur la série d’images représentant des plantules d’âges différents, les cotylédons sont soulevés au-dessus du sol (germination épigée) grâce à la croissance d’un organe intermédiaire entre la racine et l’apex, l’hypocotyle.
Les deux images suivantes montrent deux états des plantules de soja dans une barquette de germination. Douze heures séparent les deux photographies.
 |
 |
| Début de l'observation (t = 0) | Fin de l'observation (t = 12 h) |
Une séquence vidéo montre la cinétique de cette expérience.
Il est relativement aisé avec ce matériel d’établir une courbe de croissance de manière qualitative et quantitative. Toutes les germinations épigées de type « haricot » se réalisent selon le même processus.
 |
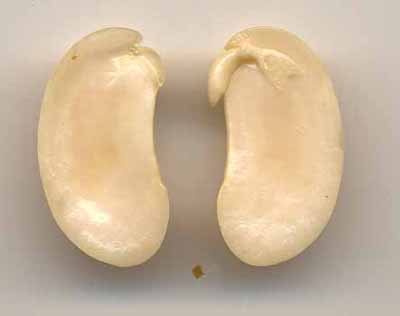
Comme chez le pois, les réserves sont contenues dans les cotylédons qui appartiennent à l’embryon. Ces graines sont donc exalbuminées. Chez le soja, une grande partie des réserves est de nature protéique. Chez le haricot, les réserves sont en majorité amylacées. En ouvrant une graine de haricot entre les deux cotylédons puis en la colorant par le lugol, on met bien en évidence son anatomie et la localisation de ses réserves.
 |
 |
L’étiolement
La plupart des expériences concernant la croissance sont réalisées sur des plantules étiolées, c’est-à-dire dans des conditions peu naturelles. Pourquoi ? La lumière inhibe sensiblement la croissance. À l’obscurité, la croissance par auxèse des coléoptiles, épicotyles ou hypocotyles est plus forte et les réponses hormonales (à l’auxine en particulier) plus nettes. On peut ainsi se poser la question de l’environnement lumineux sur la morphogenèse d’une plantule.
Observons deux exemples, le pois (dicotylédones) et le maïs (monocotylédones).
 |
| Germinations de pois cultivées à la lumière (à gauche) et à l'obscurité (à droite) |
 |
|
| Germinations de maïs cultivées à la lumière (à gauche) et à l'obscurité (à droite) | |
Le pois cultivé à l’obscurité présente les symptômes suivants, comme la plupart des dicotylédones :
- croissance en longueur des entre-nœuds beaucoup plus forte ;
- croissance des feuilles plus faible ;
- absence de chlorophylle (la tige est blanche et les feuilles jaunes).
Le maïs réagit de manière légèrement différente, comme beaucoup de monocotylédones.
- croissance en longueur du coléoptile plus forte ;
- croissance des feuilles peu modifiée ;
- absence de chlorophylle (le coléoptile est blanc et les feuilles jaunes).
Les modifications touchent donc :
- la croissance en longueur des tiges ;
- la différenciation de l’appareil photosynthétique (développement des feuilles, différenciation des chloroplastes, synthèse de chlorophylle).
Quelle peut être la signification biologique de la croissance exacerbée observée à l’obscurité ? Réalisons une expérience très simple. Dans un pot recouvert de papier aluminium, plaçons des grains de maïs à différentes profondeurs. Lorsque les jeunes feuilles sortent à la surface, déterrons les germinations.
 |
 |
On constate que quelle que soit la profondeur du semis, les parties aériennes sont au même niveau. Ceci est dû à la croissance du mésocotyle, très rapide à l’obscurité (en profondeur) et qui se ralentit lorsque les parties aériennes arrivent en surface.
Cette expérience montre que le facteur externe lumière (présence ou absence) permet à la plante de régler sa morphogenèse en s’adaptant aux conditions locales et momentanées.
Conclusion
Germinations et jeunes plantules constituent un matériel de choix pour l’expérimentation sur la croissance. Chaque type de germination possède ses règles propres de morphogenèse dues à son génotype. Cependant le phénotype de la plantule, c’est-à-dire son aspect et sa physiologie (et aussi sa constitution biochimique) dépend des facteurs de l’environnement qui ont pu agir sur son développement.
Le facteur le plus aisé à mettre en évidence est la présence ou l’absence de lumière (étiolement). La lumière agit dans ce cas par son intensité, et plus spécifiquement par sa longueur d’onde comme signal morphogène.
Dans certains cas, la lumière peut également agir par sa direction. Elle produit alors des réactions de croissance différentielle qui modifient la position de certains organes. La réaction de la plante à cet éclairement directionnel est appelé le phototropisme.
D’une manière plus générale, les tropismes sont des réactions de courbure dues à une croissance différentielle provoquée par un facteur dissymétrique de l’environnement. On peut étudier assez aisément les réactions provoquées par la lumière (le phototropisme), par la gravité (le gravitropisme) et par un contact (le thigmotropisme). Ces réactions, mises en évidence expérimentalement lorsque l’on ne fait varier qu’un seul facteur, s’intègrent dans les réactions naturelles à l’ensemble des facteurs internes et externes qui règlent la morphogenèse.
Voir également le second article sur les plantules modèles, ainsi que les chapitres correspondants de l’article sur la Localisation de la croissance.



