Présentation des voies optiques chez la grenouille et de la correspondance entre l'organisation de la rétine et celle du tectum (toit optique).
Protocole de l'expérience de Sperry (autogreffe d'œil en position inversée) et interprétation des résultats.
Introduction
Spécificité et plasticité sont deux propriétés essentielles des réseaux de neurones qui, bien qu’apparemment opposées, sont complètement liées, dans la mesure où la plasticité d’un réseau d’éléments connectés ne peut être révélée que par rapport à une organisation spécifique de ce même réseau. Une telle organisation apparaît la première au cours du développement du réseau. Sa plasticité peut s’exprimer ensuite sous l’action de facteurs provenant soit de l’environnement, soit induits par l’expérience.
Lors de ces trente-cinq dernières années, avec les progrès des techniques comportementales, électrophysiologiques, anatomiques expérimentales et immunocytochimiques (entre autres), des études fouillées portant sur différents modèles expérimentaux ont apporté un ensemble de données souvent complémentaires dans la connaissance de la spécificité et de la plasticité des réseaux de neurones.
Parmi ces modèles, celui du comportement alimentaire des Amphibiens (grenouilles Rana, crapauds Bufo, salamandres Salamandra) fut utilisé avec succès. Ce document permet d’illustrer comment des stimuli externes peuvent être représentés au niveau cortical, selon un principe rappelant la somesthésie somato-sensorielle. Les expériences présentées abordent quelques notions de spécificité nerveuse, mais aussi de plasticité neuronale. Le dossier est organisé en quatre courts documents complémentaires.
Les voies optiques chez la grenouille
Présentation générale
La vision permet à un organisme animal de s’informer sur son environnement, lui permettant ainsi de repérer ses proies, de s’éloigner de ses prédateurs, etc. Chez les Vertébrés, cette capacité est rendue possible par l’existence d’organes sensoriels particuliers : les yeux, et en particulier la rétine.
La rétine est une excroissance du cerveau, qui se forme pendant le développement embryonnaire à partir du diencéphale (une des cinq vésicules céphaliques). Elle est composée de divers types cellulaires, en particulier ceux capables de détecter certains stimulus lumineux : les cônes et les bâtonnets. Toutefois, l’œil en lui-même n’est pas capable d’interpréter les informations ainsi perçues. Cette interprétation est réalisée par l’encéphale de l’animal. Ainsi, les informations captées par les cellules de la rétine sont véhiculées, sous forme de messages nerveux, depuis l’œil jusqu’à ce cerveau, grâce au nerf optique. On parle pour cela de voies visuelles primaires. Selon les groupes de Vertébrés, la région du cerveau qui reçoit ces informations et les interprète en premier est variable. Ainsi, les nerfs optiques véhiculent chez les Mammifères une information visuelle qui, après relais au niveau du thalamus, aboutit au niveau du cortex occipital. Après les premières interprétations (forme du stimulus, couleur, mouvement, etc.), des mécanismes plus complexes (pouvant mettre en jeu d’autres zones de l’encéphale) aboutissent à la représentation consciente du stimulus par l’animal, et par voie de conséquence à une modification appropriée de son comportement.
Chez les Amphibiens, les informations véhiculées par les nerfs optiques aboutissent à une zone différente de l’encéphale : le toit optique, ou tectum. Celui-ci est issu de la différenciation de la paroi dorsale du mésencéphale (la troisième vésicule embryonnaire de l’encéphale). Pour plus d’information sur les vésicules neurales embryonnaires et leur devenir chez les Vertébrés, voir le document correspondant.
Deux grands types de voies
De manière plus précise, les fibres composant le nerf optique issu d’un œil se séparent en deux grands groupes, en fonction de leur destination : une partie de ces fibres se dirige vers le centre optique (tectum chez les Amphibiens) situé du même côté (droite ou gauche) que l’œil, alors que les autres se dirigent au contraire vers l’autre toit optique, situé donc du côté opposé. On parle respectivement de voie ipsilatérale et de voie controlatérale. Cette séparation entre les fibres controlatérales et ipsilatérales se réalise à l’endroit où les deux nerfs optiques se croisent. Cette zone de croisement des deux nerfs optiques est nommée le chiasma optique.
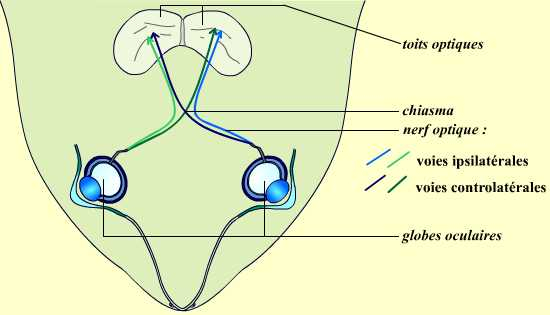
Les deux nerfs optiques, issus des deux yeux, se croisent au niveau du chiasma optique. À ce niveau, une partie des fibres nerveuses se projette vers le toit optique (centre primaire de la vision chez la grenouille) situé du côté opposé (voie controlatérale), alors qu’une autre partie des fibres se projette vers le toit optique situé du même côté (voie ipsilatérale).
Chez un Mammifère, on observe le même type de trajets, à la différence notable que le centre optique principal vers lequel se projettent les fibres n’est pas le toit optique mais le cortex visuel (région occipitale des hémisphères cérébraux).
Sur ce schéma, les proportions ne sont pas respectées.

Chez la grenouille, la voie controlatérale est très fortement majoritaire. Nous ne représenterons donc que cette voie dans l’ensemble du dossier, par souci de simplicité.
Une innervation de différentes parties du cerveau
Toutes les fibres du nerf optique ne se dirigent pas vers le tectum : certaines d’entre elles, chez la grenouille, se dirigent en fait vers le prétectum, une structure située en position juste antérieure du tectum, mais issue d’une autre vésicule embryonnaire, le diencéphale. C’est à ce niveau que certaines interprétations du stimulus visuel peuvent être réalisées, et en particulier que la distinction entre une proie et un prédateur est faite.
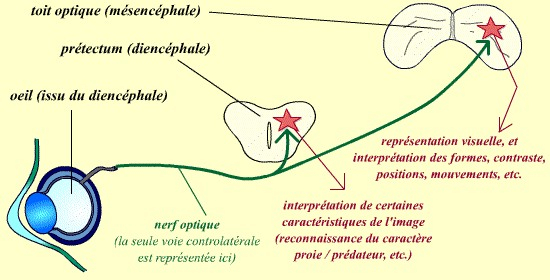
Alors que la majorité des fibres du nerf optique continue vers le toit optique, une partie des fibres se projette vers un deuxième centre optique (le prétectum).
Seule la voie controlatérale est représentée ici (elle correspond à 90 % des neurones visuels chez la grenouille, pour seulement 10 % à la voie ipsilatérale).
Il apparaît donc intuitivement que la survie d’une grenouille, intimement liée à sa bonne interprétation des stimuli visuels l’entourant, dépend d’une bonne perception de ces stimuli (par la rétine) et d’une bonne interprétation de ceux-ci (par le tectum et le prétectum). Cette interprétation ne peut être correcte que dans la mesure, a priori, où des connexions nerveuses spécifiques ont été réalisées entre les neurones récepteurs de la rétine et les neurones de ces structures céphaliques. Cette spécificité des connexions correspond à la rétinotopie.
Quelques informations complémentaires sur les voies visuelles primaires des Vertébrés sont disponibles dans le document correspondant. De même, des informations anatomiques sur ces voies et sur l’encéphale des Amphibiens sont disponibles sur le site Biologie et Multimédia.
La rétinotopie : objet réel, images optique et corticale
De la perception du stimulus à la réponse comportementale
L’œil est l’organe sensoriel qui permet à la grenouille (comme aux autres Vertébrés) de détecter des stimuli visuels. Les informations captées ainsi sont transmises à un centre nerveux du cerveau : le toit optique, ou tectum, chez les Amphibiens. C’est celui-ci qui assure l’essentiel du traitement de ces informations visuelles (avec l’aide du prétectum).
Du tectum partent ensuite des efférences motrices : ces voies nerveuses permettent les mouvements musculaires correspondant à la réponse comportementale que l’animal va apporter au stimulus perçu. Dans le cas de la grenouille (pour qui la question essentielle est de savoir si elle a affaire à une proie – à capturer – ou à un prédateur – à fuir), les muscles innervés par ces efférences motrices sont ceux permettant la locomotion, ainsi que les mouvements de la bouche.
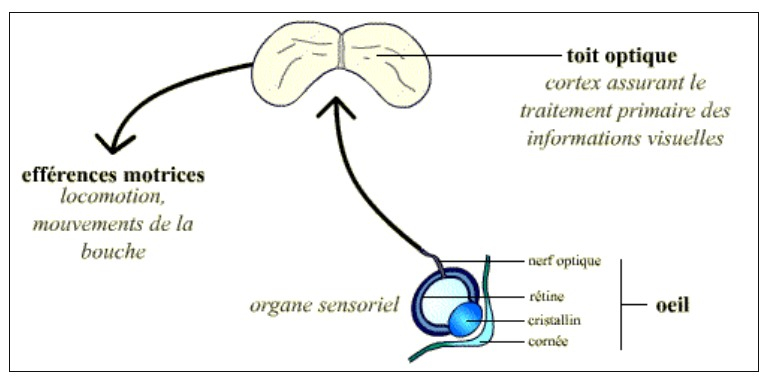
L’œil est un organe sensoriel; les informations qu’il capte (stimuli visuels) sont transmises à une zone précise du cerveau, qui les interprète. Chez la grenouille, il s’agit du toit optique.
Celui-ci émet des efférences motrices qui innervent divers muscles du corps (via le bulbe rachidien et la moelle épinière) permettant ainsi à l’animal d’adopter le comportement adapté (attaque d’une proie ou fuite devant un prédateur).
Ces efférences motrices se divisent en deux groupes, chacun correspondant à une posture : orientation et déplacement, d’une part, happement d’autre part.
Le premier groupe a pour origine principale la couche 6 du tectum antérieur, qui reçoit des terminaisons visuelles des régions temporale et centrale de la rétine. Les axones efférents constituent une voie croisée qui, après relais dans la réticulée bulbaire, se prolonge jusqu’aux différents étages de la moelle épinière, assurant la réalisation des mouvements nécessaires à l’orientation puis aux déplacements.
Le second groupe a sa source dans la couche 7 du tectum postérieur, qui reçoit les fibres rétinofuges nasales. Les fibres tectofuges se réunissent en un faisceau direct qui descend jusqu’à la moelle en réalisant sur son trajet de nombreuses synapses avec les noyaux moteurs des nerfs V et VII (ouverture de la mâchoire), et celui de l’équivalent du nerf XII humain (premier nerf spinal chez la grenouille ; dévagination de la langue), assurant ainsi le happement de la proie.
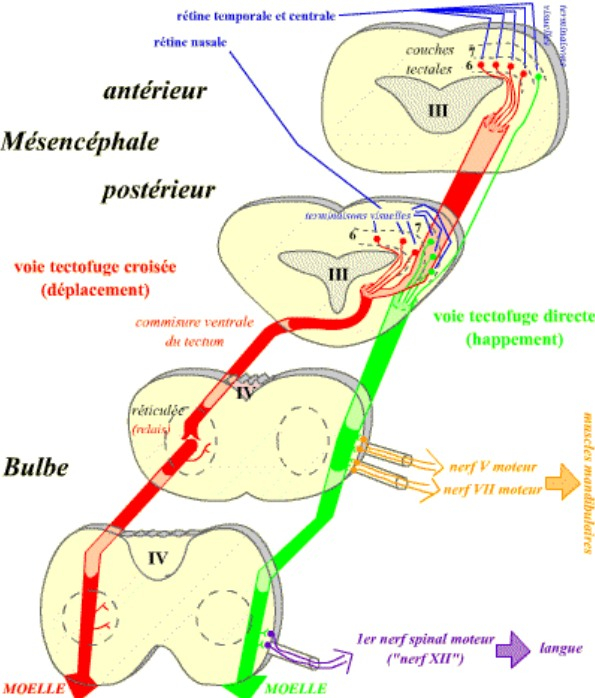
Ces voies sont représentées grâce à quatre sections schématiques au niveau du mésencéphale et du bulbe de la grenouille.
Les voies qui se poursuivent dans la moelle épinière permettent d'innerver les muscles des pattes arrières en particulier (et déclencher le déplacement par exemple).
Organe sensoriel et cortex peuvent être définis selon certains axes
Les deux yeux de la grenouille sont situés de part et d’autre de sa tête (donc en position latérale). On peut définir à leur niveau (et donc aussi au niveau de leur rétine) deux axes majeurs : l’axe nasal-temporal (N-T) et l’axe dorso-ventral (D-V).
De même, le cortex visuel associé à cette rétine (le tectum) présente lui aussi deux axes principaux : l’axe rostro-caudal (R-C), et l’axe médio-latéral (M-L).
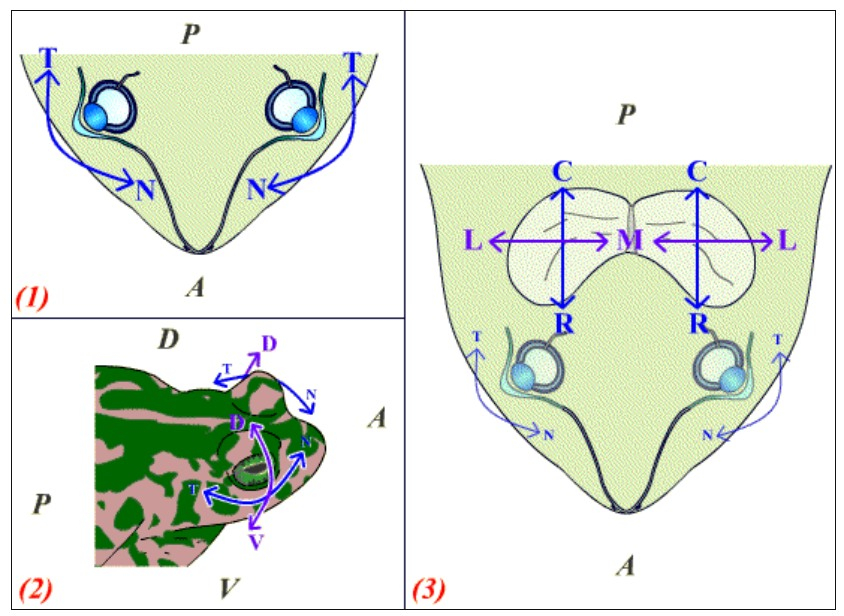
Plusieurs axes sont décelables au sein de ces deux organes.
Ils sont représentés ici sur une tête de grenouille en vue dorsale (1, 3) ou latérale (2). A : antérieur ; P : postérieur ; D : dorsal ; V : ventral.
(1) Vue dorsale montrant l’axe nasal (N) – temporal (T) des deux yeux de la grenouille.
(2) Vue latérale montrant les axes nasal (N) – temporal (T) et dorsal (D) – ventral (V) des yeux de grenouille.
(3) Vue dorsale montrant les axes rostro (R) – caudal (C) (similaire à l’axe antéro-postérieur) et médio (M) – latéral (L) du toit optique de la grenouille.
La rétinotopie : une correspondance entre ces axes
De manière intéressante, on peut observer qu’un neurone situé sur la rétine de la grenouille ne se projette pas n’importe où, au niveau du toit optique. Au contraire, on observe une véritable colinéarité entre les axes de la rétine et ceux du tectum. Ainsi, les neurones disposés selon l’axe dorso-ventral, sur la rétine de l’œil, se projettent sur l’axe médio-latéral du toit optique. Une même correspondance s’observe entre les deux autres axes.

Les neurones issus de la rétine de l’œil se projettent vers des régions spécifiques du tectum. Ainsi, les neurones rétiniens observables selon l’axe nasal-temporal (N-T) se projettent selon l’axe caudal-rostral du tectum (C-R) : les neurones nasaux se projettent vers la région caudale du tectum, tandis que les neurones temporaux se projettent vers la région rostrale du tectum.
On observe la même correspondance des neurones rétiniens de l’axe dorso-ventral (D-V) vers l’axe médio-latéral (M-L) du tectum.
N: nasal ; T : temporal ; D : dorsal ; V : ventral ; C : caudal ; R : rostral ; L : latéral ; M : médian.
Les trajets des neurones représentés ici sont simplifiés et schématisés pour une meilleure compréhension.
Cette rétinotopie (projection exacte des neurones rétiniens vers le tectum) s’observe aussi chez les autres Vertébrés, tels que les Mammifères, mais parfois de manière plus complexe.
Cette projection exacte des neurones de la rétine vers des régions bien définies du cortex qui interprète les caractéristiques du stimulus visuel est la base d’une « représentation » virtuelle de ce stimulus au niveau dudit cortex. Il s’agit là d’une spécificité entre la rétine et le tectum (ou le cortex occipital chez les Mammifères) fondamentale dans la bonne interprétation du stimulus visuel. On peut rapprocher cette spécificité de la représentation du corps que l’on observe au niveau des cortex sensoriels et moteurs des Mammifères (la somesthésie).
La rétinotopie permet l’existence de plusieurs « images » du stimulus visuel
Le stimulus visuel perçu par la grenouille est un objet « réel ». Il peut être perçu par la capacité qu’il a de réfléchir la lumière (plus rarement de l’émettre directement), selon certaines longueurs d’onde. Ce sont ces rayons lumineux, de longueurs d’ondes précises, qui sont détectés par les neurones récepteurs de la rétine de l’œil de grenouille.
Pour atteindre la rétine, ces rayons lumineux passent à travers un système optique complexe, composé de la cornée, du cristallin, et des humeurs de l’œil (qui remplissent le globe oculaire). Ce passage à travers ce système optique permet la formation d’une image optique sur la rétine, parfaitement nette quand le stimulus est situé à une distance permettant à la grenouille d’accommoder. C’est en fait cette image optique qui est détectée par la rétine. Il s’agit donc d’une représentation de l’objet réel qu’est le stimulus placé devant la grenouille. On peut remarquer qu’en cas « d’anomalie » du système optique, l’image optique est modifiée, et ainsi la perception du stimulus est incorrecte. C’est ce qui se passe, par exemple, dans le cas d’une myopie, d’une presbytie, etc. chez l’homme.
Cette image optique, détectée par la rétine, est transmise sous forme d’impulsions électriques (les trains de potentiels d’action) jusqu’au tectum. La rétinotopie, en permettant une correspondance exacte entre les axes de la rétine et ceux du tectum, permettent donc la formation d’une image tectale, « virtuelle », à son niveau : le stimulus détecté est représenté au niveau du tectum par l’excitation de neurones précis ; ces neurones excités reproduisent la forme (et les autres caractéristiques) du stimulus.
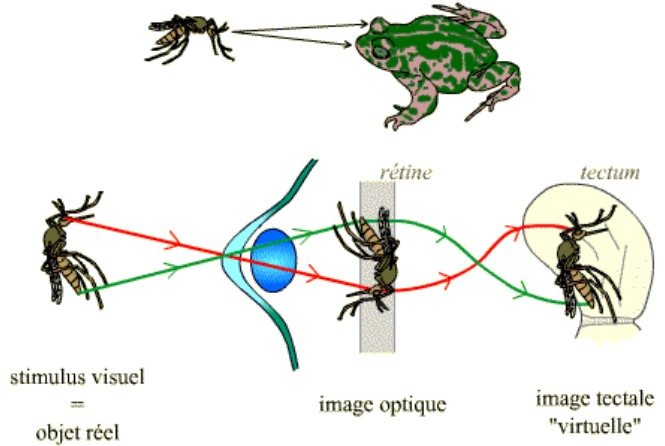
Le système optique constitué par l’œil, ainsi que la rétinotopie entre la rétine et le tectum permettent d’obtenir une image optique, au niveau de la rétine, du stimulus (objet réel), puis une représentation de celle-ci sous forme d’une image tectale au niveau du toit optique.
Les traits verts et rouges schématisent le trajet des rayons lumineux (du stimulus à la rétine) puis les informations visuelles (de la rétine au tectum).
On peut remarquer que l’image optique est inversée par rapport à l’objet réel. Ceci est dû au fait que le système optique de l’œil se comporte, globalement, comme une lentille convergente. Le cheminement des neurones de la rétine vers le tectum permet en fait d’aboutir à une image tectale qui est elle dans le « même sens » que l’objet réel.
Expérimentalement, il est possible chez la grenouille de modifier l’orientation des axes de la rétine. Ces expériences sont riches d’enseignements, aussi bien sur la spécificité déjà présentée ici des connexions rétino-tectales, mais aussi sur la plasticité du système nerveux…
La spécificité des connexions : expérience de Sperry
L’expérience de Sperry
Cette expérience consiste, globalement, en le retournement d’un œil de grenouille. Pour cela, on procède en deux temps : dans un premier temps, un œil est retiré d’une grenouille. Ceci permet d’avoir une orbite vide, qui sera ensuite utilisée pour y greffer, dans un deuxième temps, l’autre œil de la grenouille. Il s’agit donc d’une auto-greffe, un œil de la grenouille étant greffé du côté opposé.
Au cours de cette greffe, on retourne l’œil de la grenouille. Ceci peut être réalisé soit selon l’axe dorso-ventral, soit selon l’axe nasal-temporal. Les deux types d’expériences donnent des résultats sensiblement comparables.
Dans la suite de ce dossier, nous détaillerons l’exemple d’un retournement dorso-ventral de l’œil.
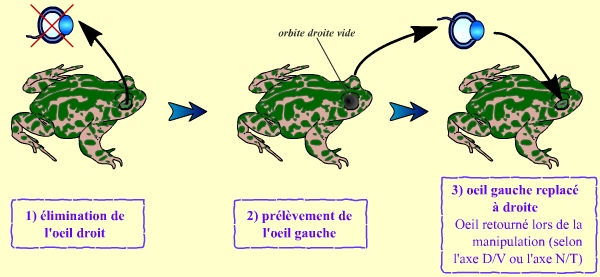
Cette expérience consiste en une autogreffe d’un œil de grenouille du côté opposé à son emplacement initial.
La manipulation permet de retourner l’œil greffé, soit selon l’axe dorso-ventral (D-V), soit selon l’axe nasal-temporal (N-T).
Résultat de l’expérience : plasticité et spécificité
Rétablissement des connexions : plasticité
Lorsque l’expérience est terminée, les deux extrémités des nerfs optiques (le bout de l’œil gauche transplanté à droite et le bout du nerf droit resté en place) sont aboutés fermement. On observe alors que les axones issus de la rétine de l’œil greffé régénèrent (au moins une partie significative d’entre eux), et retrouvent le chemin du tectum.
Ce processus traduit une plasticité régénérative notable. On peut penser qu’il est rendu possible par l’émission de signaux dits « de position » par les cellules gliales (entre autres) de l’environnement neuronique.
Rétablissement des bonnes connexions : spécificité
Dans le tectum, les mêmes axones reconnaissent leur territoire de projection d’origine (par exemple, les axones issus de la rétine ventrale rejoignent le tectum médial, etc.) : la projection rétinotopique est conservée.
Ainsi, les connexions rétinotopiques manifestent une réelle spécificité. Là encore, des signaux moléculaires émis par les différentes parties du tectum sont probablement en jeu.
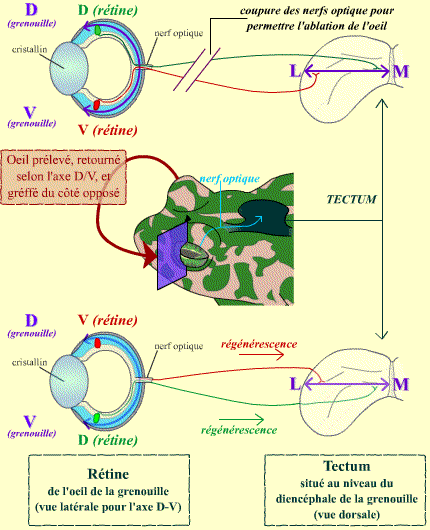
Après réalisation de l’ablation d’un œil de grenouille, suivie de sa greffe, on observe que les connexions nerveuses entre cet œil greffé et le toit optique (tectum) se rétablissent.
Ce rétablissement se fait de manière spécifique, en fonction des polarités originelles de la rétine de l’œil greffée (par exemple, les axones issus de la rétine dorsale se projetant au niveau du tectum médial), mais pas de la position réelle de l’œil greffé. Ceci conduit donc en fait à une inversion des connexions, l’œil étant retourné au cours de la manipulation.
Le schéma présente les connexions « normales », en haut, et les connexions inversées obtenues après la greffe, en bas.
Conséquences neuronales de la greffe
L’inversion de la polarité rétinienne entraîne une inversion de l’image tectale, précisément parce que ces connexions rétino-tectales restent spécifiques. La perception qu’a ainsi la grenouille de son environnement est modifiée : ceci bouleverse le comportement de l’animal dans la compréhension de cet environnement.
Conséquences comportementales de l’expérience de Sperry
Comportement alimentaire des Amphibiens
Le comportement alimentaire des Amphibiens implique la vision comme fonction sensorielle (afférente) et les membres ainsi que la région buccale comme fonctions motrices (efférentes). Dans la mesure où les anoures et les urodèles sont des carnassiers, leur comportement alimentaire est celui d’une capture de proie. Les éthologistes qui l’ont étudié en ont conclu qu’il s’agissait d’un comportement stéréotypé (rigide), divisible en plusieurs étapes. Chaque étape est caractérisée par une attitude corporelle (posture) qui induit l’étape suivante.
-
Étape 1 : détection du stimulus
Le champ visuel des Amphibiens, en particulier celui de la grenouille, est particulièrement vaste (jusqu’à 320°). Tout stimulus mobile y sera détecté, mais sa reconnaissance reste encore rudimentaire, puisque la grenouille n’a une vision nette que des objets situés à moins de 1 mètre de son museau ! Néanmoins, deux paramètres du stimulus, en plus de la mobilité, sont bien analysés par l’animal : le contraste (par exemple un stimulus noir sur un ciel bleu) et la taille.
-
Étape 2 : orientation
Une fois le stimulus détecté, la grenouille tourne d’abord la tête dans sa direction de façon à faire converger ses deux axes optiques vers le stimulus. Elle place ensuite son corps entier dans l’axe de sa tête, orienté donc vers le stimulus.
-
Étape 3 : le déplacement
L’animal reste rigidement dirigé face au stimulus, qu’il suit dans ses déplacements (on parle parfois de « poursuite »)
-
Étape 4 : l’interprétation du stimulus
C’est une étape dont le mécanisme est encore inconnu précisément. Pourtant, il s’agit là d’une phase cruciale du comportement, qui conditionne son déroulement complet. En effet, selon l’interprétation que fait l’animal de la taille réelle du stimulus, celui-ci est interprété soit comme une proie, soit comme un prédateur (il en va donc de la survie de l’animal !). En plus de la taille réelle, la position du stimulus par rapport à son axe de déplacement joue aussi un rôle non négligeable : si l’objet est placé perpendiculairement à cet axe, il sera interprété comme un prédateur. En fonction de l’interprétation, deux postures peuvent être adoptées :
- L’évitement : il s’agit d’une fuite devant le stimulus, orientée à 90° par rapport au sens de déplacement de celui-ci, considéré alors comme prédateur. On sait que l’évitement est enclenché à partir d’un ensemble de neurones situés dans le prétectum.
- L’approche : dans le cas d’une interprétation « proie », la grenouille continue à avancer en direction du stimulus.
-
Étape 5 : le happement
Suite à l’approche vers la proie : lorsque la proie est environ à 30 cm de son museau, la grenouille saute en direction de sa proie, ouvre sa bouche et dévagine sa langue. Cette posture implique le fonctionnement coordonné d’un ensemble de plusieurs voies motrices.
-
Étape 6 : l’essuyage
C’est un curieux réflexe, inexpliqué, qui consiste en un passage d’une patte antérieure sur le museau après ingestion de la proie. Ce réflexe est surtout présent chez le crapaud.
Expérience de Sperry, conséquences neuronales
Un œil de grenouille adulte est prélevé, puis greffé dans l’orbite laissée vide par l’élimination de l’œil situé du côté opposé. Au cours de la manipulation, l’œil greffé est retourné selon l’axe dorso-ventral, par exemple.
On observe alors une remarquable plasticité du système nerveux, les axones régénérant et les connexions nerveuses se rétablissant, mais aussi une importante spécificité : en effet, les axones issus de la rétine de l’œil greffé se projettent vers les mêmes territoires du tectum qu’avant la greffe. Ainsi, les axones issus de la rétine dorsale se projettent toujours vers le tectum médial, et les axones issus de la rétine ventrale vers le tectum latéral.
Or, l’œil ayant été inversé au cours de la greffe, ce rétablissement des connexions conduit en fait à inverser les connexions normales : Les axones de la rétine dorsale, qui se projettent vers le tectum médial, sont en effet situés (du fait du retournement expérimental) en position ventrale… Il en est de même pour les axones issus de la rétine ventrale.
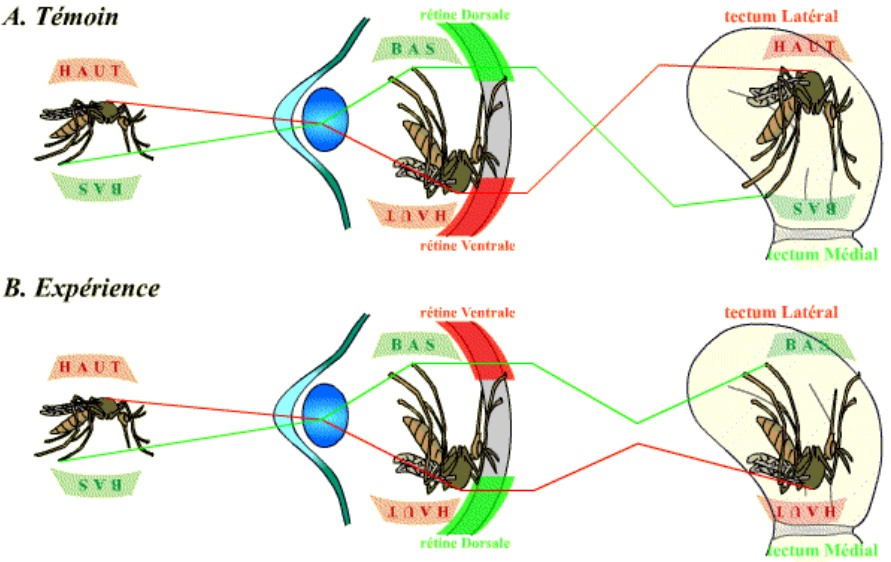
A. Témoin. Chez l’animal non manipulé, un stimulus (à gauche) se projette en une image optique au niveau de la rétine (au milieu), qui est traduite au niveau du toit optique sous la forme d’une image tectale (à droite). Les connexions entre les régions dorsales et ventrales de la rétine et les régions, respectivement, médianes et latérales du tectum conduisent à la représentation du « haut » du stimulus au niveau du tectum latéral, et du « bas » du stimulus au niveau du tectum médial.
B. Expérience. On a réalisé ici une autogreffe d’un œil de grenouille, en inversant la polarité dorso-ventrale de celui-ci. Les connexions entre la rétine et le toit optique reconstituées correspondent à la polarité originelle de la rétine : ainsi, la partie ventrale de la rétine (positionnée en dorsal du fait de l’expérience) ré-établit des connexions nerveuses avec le tectum latéral. Cette inversion conduit alors à une représentation tectale inversée : le « haut » du stimulus est désormais représenté au niveau du tectum médial, et le « bas » du stimulus est représenté au niveau du tectum latéral.
On s’aperçoit ainsi que ce retournement de l’œil conduit à l’obtention d’une image tectale inversée par rapport à la normale. Cette inversion bouleverse le comportement de la grenouille.
Des modifications comportementales spectaculaires
On observe le comportement d’une grenouille témoin et celui d’une grenouille chez qui on a greffé un œil en inversant la polarité dorso-ventrale, face au stimulus constitué par une proie se déplaçant du bas vers le haut.
Chez l’animal témoin, l’image rétinienne est inversée et l’image tectale se déplace latéro-médialement. Ainsi lorsque l’objet est au sommet de sa trajectoire, il est « vu » sur la partie la plus médiane du tectum. La grenouille s’étant déjà suffisamment rapprochée de cet objet (ici une proie), elle va pouvoir le happer en sautant vers le haut.
Chez l’animal greffé, la même image rétinienne que celle de l’animal témoin est projetée sur le tectum en sens inverse que chez ce dernier. Ainsi, l’animal voit et interprète la trajectoire de l’objet en sens inverse de la réalité. Il en résulte un comportement « a contrario », c’est-à-dire que la grenouille voit sa proie non plus au zénith, mais en bas du champ visuel inférieur. Après l’approche (non affectée par la transplantation), la grenouille va sauter vers ce qu’elle croit être sa proie (en fait un objet virtuel qui n’existe que dans son tectum) en direction du bas, et sa bouche ne va happer que du vide !
Conclusion
La vision de la grenouille, comme de tout vertébré, est permise par des connexions spécifiques entre ses organes visuels (les rétines) et le cortex traitant ces informations (ici le tectum). Les expériences de greffe de Sperry montrent ainsi une formidable spécificité de ces connexions, en même temps qu’une plasticité importante. Cette plasticité est corrélable à la nature même du système nerveux, ensemble de réseaux de neurones extrêmement complexe, qui se met en place de manière progressive au cours du développement, embryonnaire, et continue à évoluer pendant toute la vie de l’individu.