Il est difficile de savoir quels gènes sont responsables du phénotype trisomique 21. Pour essayer d'y voir plus clair, des chercheurs ont produit des modèles murins possédant en triple exemplaire certains gènes suspectés d'être à l'origine de ce phénotype.
La trisomie 21 ou syndrome de Down, anciennement dénommé mongolisme, affecte chez les humains une naissance sur 700. Elle se manifeste de façon plus ou moins sévère mais comporte toujours les caractéristiques suivantes : anomalies du développement crâno-facial et retard mental. Des études menées récemment chez la souris apportent des informations nouvelles sur son déterminisme.
La trisomie 21 chez l’Homme
La trisomie 21 consiste en l’adjonction de tout ou partie d’un troisième chromosome 21, au lieu des deux existant normalement (Lire également La trisomie 21 : origine et quelques chiffres ). Cette triplication totale ou partielle est délétère parce qu’elle dérègle l’action de certains gènes, soit dans le sens de l’excès, soit dans celui du manque.
Grâce au séquençage du génome humain et aux énormes efforts déployés pour en décompter les gènes, le chromosome 21 humain est mieux connu : l’estimation actuelle est de 231 gènes sur son bras long (appelé q21), couvrant 33,5 millions de paires de bases (33,5 Mb).
La trisomie 21 totale
C’est de loin la plus fréquente avec 95 % des cas. Elle provient le plus souvent d’une mauvaise disjonction des chromosomes homologues 21 à deux chromatides lors de la première division de méiose paternelle ou maternelle. Au terme de la méiose 1, les deux chromosomes homologues se retrouvent donc ensemble dans l’une des cellules filles (l’autre en étant privée). Ce surnombre est conservé au cours de la deuxième division de méiose qui fait passer les chromosomes de deux chromatides à une chromatide. On peut aussi avoir une mauvaise répartition des chromosomes fils lors de la deuxième division de méiose.
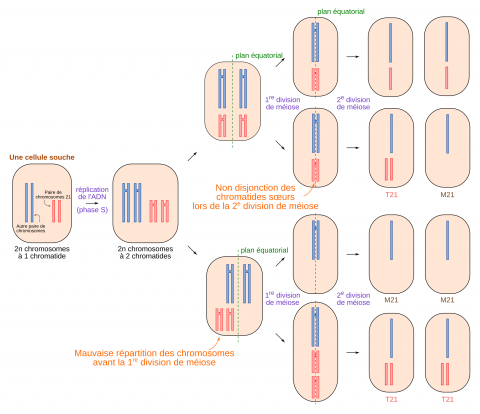
Des gamètes contenant deux chromosomes 21 peuvent être formés à cause d’anomalies lors de la première ou lors de la seconde division de méiose. Si un tel gamète féconde un gamète possédant un chromosome 21, l’embryon résultant possédera trois chromosomes 21. De telles méioses produisent également des gamètes sans chromosome 21. Cependant des embryons issus de ces gamètes ne sont pas viables.
Si un gamète à deux chromosomes 21 s’unit lors de la fécondation à un gamète pour lequel la méiose s’est déroulée sans accident (c’est-à-dire possédant un seul chromosome 21), cela donne une cellule-œuf à trois chromosomes 21 : c’est la trisomie totale. La question se pose alors de savoir lesquels parmi les 231 gènes présents en triple exemplaire sont à l’origine du syndrome observé.
La trisomie 21 partielle et l’hypothèse DSCR
La trisomie 21 partielle est bien plus rare que la trisomie totale : elle représente moins de 5 % des cas. Les trisomiques concernés n’ont que deux chromosomes 21 mais un examen attentif de leur caryotype révèle la présence d’un segment surnuméraire de chromosome 21 sur l’un des deux chromosomes 21, ou sur un autre chromosome. Le mécanisme à l’origine de la présence de ce morceau de chromosome surnuméraire est une translocation. Ce segment est donc représenté en trois exemplaires dans le caryotype des personnes atteintes, d’où le terme de trisomie 21 partielle.
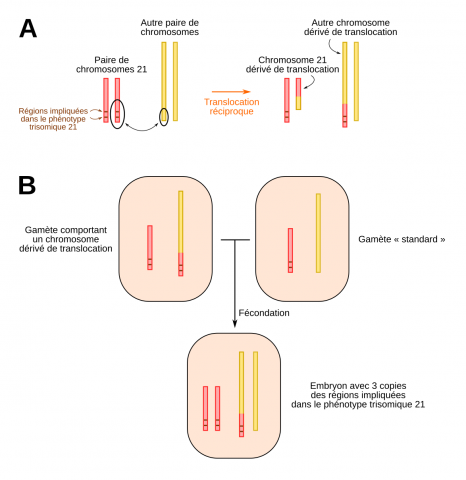
(A) Une translocation réciproque correspond à un échange de matériel entre deux chromosomes. (B) Si un des gamètes contient un chromosome dérivé de translocation, l’embryon possédera donc trois morceaux de chromosomes 21 et présentera donc le phénotype trisomique 21.
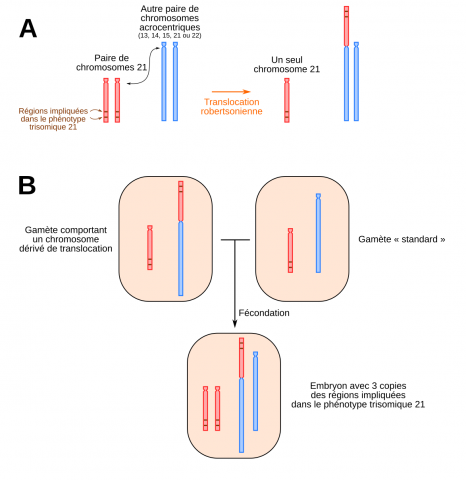
(A) Une translocation robertsonienne correspond à la fusion de deux chromosomes acrocentriques. (B) Si un des gamètes contient un chromosome dérivé de translocation, l’embryon possédera donc trois morceaux de chromosomes 21 et présentera donc le phénotype trisomique 21.
En 1989, la description par Rhamani de patients au phénotype de syndrome de Down, mais porteurs d’uniquement deux chromosomes 21, dont l’un possédait une petite région dupliquée, a permis de suspecter l’implication forte et quasi exclusive de cette région dans le syndrome de Down. Cette région est dénommée région critique du syndrome de Down (DSCR) et ne comporte que 33 gènes.
Si on arrivait à montrer – hypothèse attrayante – que c’est bien la triplication de la région DSCR qui est à l’origine des troubles trisomiques, le résultat serait particulièrement intéressant parce qu’il permettrait de resserrer la recherche sur 33 gènes au lieu de 231. Ce serait donc parmi eux qu’il faudrait rechercher la cible de médicaments susceptibles de soulager les troubles qui s’aggravent au cours de la vie du trisomique 21 : sensibilité aux infections respiratoires, prédisposition aux leucémies, Alzheimer précoce, etc. Ceci à la condition que ce petit nombre de gènes soupçonnés d’être majoritairement impliqués dans le syndrome de Down exerce son action en solitaire et non de conserve avec les gènes résidents des chromosomes 21.
Pour explorer cette hypothèse, il était nécessaire de disposer d’un modèle animal chez lequel on puisse produire aussi bien une trisomie totale qu’une trisomie partielle avec triplication de la seule DSCR. La souris est ce modèle animal pour les raisons suivantes :
- son génome séquencé a permis d’identifier des gènes très homologues des gènes humains (gènes dits orthologues) et particulièrement de ceux du chromosome 21 (150 gènes orthologues chez ce dernier) ;
- elle possède la même série d’os de la face et du crâne que l’homme;
- ses capacités d’apprentissage peuvent être testées de multiples façons, par exemple dans des labyrinthes.
La trisomie 21 chez la souris
Cependant la tâche fut loin d’être aisée. En effet, chez la souris les orthologues du chromosome 21 humain sont répartis sur trois chromosomes : le 10, le 16 et le 17. Et si le 16 porte la majorité de ces derniers, il héberge aussi des gènes qui, s’ils viennent à être tripliqués, conduisent à la mort de l’animal. Il faut donc tailler ces chromosomes murins sur mesure : un tour de force à chaque fois (voir Figure 3).
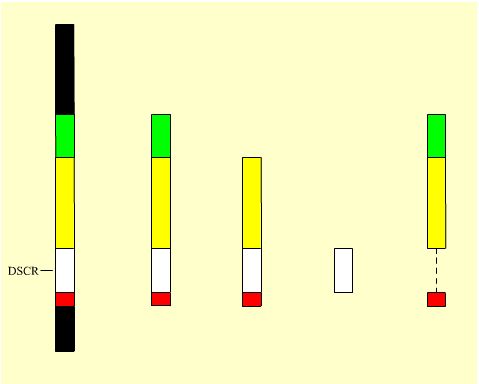
| Le K21 humain | Le K21 murin équivalent du K21 humain (modèle long) | Le K21 murin équivalent du K21 humain (modèle court) | Le K21 murin avec sa seule DSCR | Le K21 murin sans sa DSCR mais avec le reste de l'équivalent du K21 humain (modèle long) |
 |
||||
|
Mb : 33,5 Gènes : 231 |
16,4 104 |
10,3 81 |
5,3 33 |
11,1 70 |
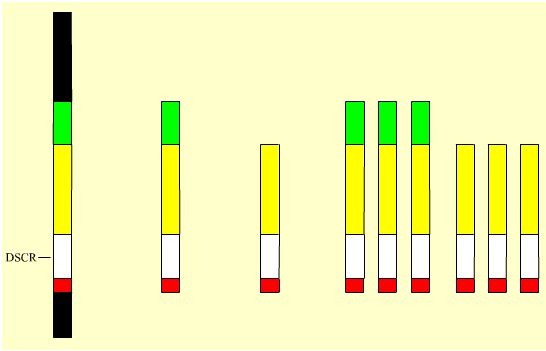
Deux modèles viables ont été obtenus (voir Figure 4 ci-dessous). Ils contiennent chacun la DSCR mais diffèrent par leur degré de trisomie (104 orthologues du chromosome 21 humain pour le modèle le plus long, 81 pour le modèle le plus court).
| Le K21 humain | Le K21 murin équivalent du K21 humain (modèle long) | Le K21 murin équivalent du K21 humain (modèle court) | Le caryotype équivalent-humain de la souris (modèle long) | Le caryotype équivalent-humain de la souris (modèle court) |
 |
||||
|
Mb : 33,5 Gènes : 231 |
16,4 104 |
10,3 81 |
16,4 16,4 16,4 104 104 104 |
10,3 10,3 10,3 81 81 81 |
De phénotypes semblables [2, 3], ces souris présentent maints aspects de la trisomie 21 humaine :
-
modifications affectant les os du crâne, de la face et de la mâchoire (modifications connues de longue date). Il faut une analyse morphométrique tridimensionnelle, avec traitement mathématique des coordonnées entre points osseux repères, pour établir cette correspondance. En effet, si l’on reconnaît d’emblée un trisomique humain, il n’en va pas de même pour la souris.
-
diminution de la densité des granules contenus dans les neurones du cervelet (modification de découverte récente [4]). Ce caractère, d’abord mis en évidence chez la souris trisomique (particulièrement chez le modèle long), a ensuite été retrouvé chez l’Homme trisomique dès lors que ce phénotype y a été recherché.
Cette correspondance étroite – et même à caractère prédictif – fait de ces souris de bons modèles du syndrome de Down. Elle témoigne aussi de l’excellente homologie entre gènes gouvernant ces caractères chez la souris et chez l’être humain.
C’est à l’équipe de R.H. Reeves, de l’université John Hopkins, Baltimore (États-Unis) que l’on doit d’avoir testé l’hypothèse d’une implication majeure de la DSCR dans la trisomie 21 au moyen d’une expérience élégante mettant en jeu deux modèles de souris complémentaires [5].
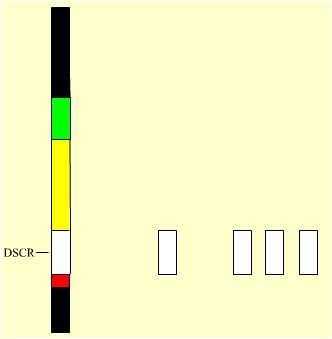
Première étape : la triplication de la seule DSCR.
Un chromosome murin a été construit avec les orthologues de la seule DSCR. Puis une souris a été produite, portant ce chromosome à l’état tripliqué (voir Figure 5 ci-dessous). Ses caractéristiques crânio-faciales ont alors été étudiées (et seulement elles à ce jour).
| Le K21 humain | Le K21 murin avec sa seule DSCR | Le caryotype équivalent-humain de la souris tri-DSCR |
 |
||
|
Mb : 33,5 Gènes : 231 |
5,3 33 |
5,3 5,3 5,3 33 33 33 |
Déception ! Toutes mesures faites, le phénotype crânio-facial trouvé n’était pas celui du syndrome de Down. La mâchoire, par exemple, est plus longue que la normale, alors qu’elle est plus courte chez la souris trisomique. La triplication de la seule DSCR ne conduit donc pas au phénotype crânio-facial typique du syndrome de Down.
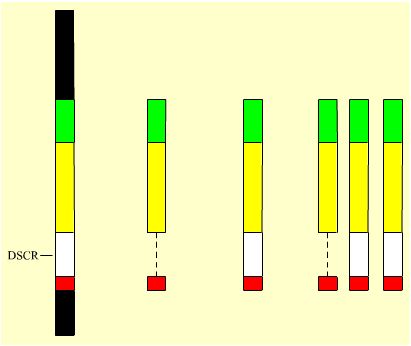
Deuxième étape : simple duplication de la DSCR, mais triplication des autres orthologues du chromosome humain. Un autre chromosome murin a été construit, excluant la DSCR, mais conservant les autres orthologues du chromosome 21 humain (modèle long). Puis il a été associé, grâce à des croisements adéquats, à deux orthologues complets du chromosome 21 humain (modèle long). La DSCR était donc à l’état disomique, et le reste de l’équivalent du chromosome 21 humain à l’état trisomique (voir Figure 6 ci-dessous).
| Le K21 humain | Le K21 murin sans sa DSCR mais avec le reste de l'équivalent du K21 humain (modèle long) | Le K21 murin équivalent du K21 humain (modèle long) | Le caryotype de la souris disomique pour la DSCR et trisomique pour le reste de l'équivalent K21 humain (modèle long) |
 |
|||
|
Mb : 33,5 Gènes : 231 |
11,1 70 |
16,4 104 |
11,1 16,4 16,4 70 104 104 |
Résultat de cette deuxième expérience : le phénotype crânio-facial, dont l’existence d’une mâchoire courte, est celui caractéristique du syndrome de Down… La déception était donc confirmée : ce n’est pas la triplication de la DSCR qui, à elle seule, conduit à l’apparition du syndrome de Down chez la souris.
Conclusion
Les modèles murins ont donc fait voler en éclats l’hypothèse attrayante de la DSCR. Et ce qui est vrai chez la souris risque fort de l’être aussi chez l’Homme, si l’on se réfère à la forte conservation des gènes observés entre ces deux espèces. L’hypothèse à explorer devient à présent la suivante : l’apparition du syndrome de Down chez l’Homme impliquerait de nombreux gènes des chromosomes 21, peut-être en interaction avec le reste du génome. Les combinaisons sont donc innombrables.
L’espoir de resserrer l’enquête sur un nombre réduit de gènes s’envole, et avec lui celui d’ouvrir dans un avenir proche des pistes thérapeutiques. Mais R.H. Reevees s’adresse alors aux chercheurs, en ces termes : « Inspirer largement, accepter la complexité de ce syndrome et repartir de l’avant ! »
Et ainsi va la recherche, et le progrès humain.
Bibliographie
- Rodolphe Dard, Gilles Furelaud, La trisomie 21 : origines et quelques chiffres , Planet-Vie, mercredi 21 novembre 2018
- Richtsmeier J.T., Baxter L.L. et Reeves R.H. " Parallels of craniofacial maldevelopment in Down syndrome and Ts65Dn mice " Dev. Dyn. (2000) 217(2):137-145.
- Richtsmeier J.T., Zumwalt A., Carlson E.J., Epstein C.J. et Reeves R.H. " Craniofacial phenotypes in segmentally trisomic mouse models for Down syndrome " Am. J. Med. Genet. (2002) 107(4):317-324.
- Reeves R.H. " A complicated genetic insult " The Lancet supplement, décembre 2001.
- Olson L.E., Richtsmeier J.T., Leszl J. et Reeves R.H. " A chromosome 21 critical region does not cause specific DS phenotypes " Science (2004) 306:687-690.