Cet article présente une technique de migration de l'ADN réalisable en classe, par électrophorèse sur gel d'agarose, et précise comment mettre en évidence des bandes de migration invisibles à l'oeil nu. Il indique la méthode permettant de déterminer la taille des fragments ainsi isolés les uns des autres. Enfin il propose une utilisation des banques de données de séquences d'ADN en ligne pour prévoir, en fonction des enzymes de restriction mises en jeu, le nombre de segments d'ADN que l'on devrait obtenir et le comparer avec les résultats de l'électrophorèse réalisée en classe.
Introduction
Les multiples applications scientifiques et technologiques impliquant l'étude de l'ADN font appel à différentes techniques. Parmi elles, l’électrophorèse sur gel d’agarose ou de polyacrylamide est une des plus communément utilisées car elle permet de séparer des molécules en fonction de leur taille, préalable indispensable dans de multiples applications, notamment pour identifier des fragments d’ADN découpés par des enzymes, pour identifier un gène ou pour établir des empreintes génétiques par Southern Blot.
L’électrophorèse d’ADN sur gel d’agarose ne présente guère de difficultés techniques et peut être utilisée aisément en travaux pratiques tant à l’université qu’au lycée pour familiariser les étudiants avec cette technique essentielle. En conjonction avec divers outils informatiques désormais communs (traitement de texte, tableur, logiciels d'acquisition et de traitement d’images), l’analyse des résultats peut conduire à des développements importants permettant de mieux comprendre l’intérêt tant scientifique que pratique des technologies de l’ADN. Différentes possibilités mettant à profit ces divers outils sont présentées ci-dessous.
Mise en pratique de l’électrophorèse d’ADN sur gel d’agarose
Gel d’agarose
- Mélanger tampon TBE et agarose à raison de 0,8 g d'agarose pour 100 mL de tampon (la proportion d’agarose dépend de la taille des molécules d’ADN à séparer).
- Faire fondre l'agarose au four à micro-ondes en surveillant pour éviter les projections ou au bain marie. Agiter de temps à autre pour homogénéiser le mélange.
- Laisser refroidir jusqu'à ce qu’il devienne possible de saisir le flacon à main nue (environ 60 °C).
- Placer les joints fournis avec la cuve pour fermer le support de gel et positionner le peigne à 1 mm du fond et à environ 1 cm de l’extrémité du support. Régler le niveau pour que le support de gel soit horizontal. Certains peignes sont réglables avec une vis : il y a alors une cale pour régler leur hauteur. D’autres sont préadaptés au support et ne nécessitent pas de cale.
- Couler lentement le gel sur 3 à 5 mm d'épaisseur en veillant à ce qu’il entoure bien les dents du peigne.
- Laisser refroidir, enlever le peigne et les joints. Le gel est prêt pour le dépôt des échantillons.
NB : On peut préparer du gel à l’avance et le conserver solide au réfrigérateur dans un flacon en verre (rempli aux 2/3 maximum). Au moment de l’emploi, refondre le gel au four à micro-ondes (ouvrir le bouchon et surveiller pour éviter les projections) ou au bain-marie bouillant (ouvrir le bouchon).

A gauche : la solution d'agarose (tampon TBE et agarose à raison de 0,8 g d'agarose pour 100 mL de tampon) dans son flacon de conservation.
A droite : la minicuve pour gel 8 x 6,5 cm, avec le peigne et les joints de coulage mis en place.
Préparation et dépôt des échantillons
Pour se familiariser avec cette technique, le plus simple est d’acheter dans le commerce des échantillons d’ADN, entier ou digéré par des enzymes de restriction. Les fournisseurs (voir en bas de page) proposent toute une gamme d’ADN de diverses sources digéré ou non par des enzymes.
Les échantillons d'ADN peuvent être obtenus en solution dans un tampon TE (Tris EDTA) ou lyophilisés. Dans ce cas, il sont remis en solution dans du tampon TE.
Les solutions d'ADN peuvent être conservées indéfiniment à -20 °C.
Les échantillons d’ADN sont le plus souvent distribués par les fournisseurs avec du colorant de charge qu’il suffit de mélanger aux échantillons avant le dépôt en suivant les proportions indiquées par le fabricant. Une pipette de précision est nécessaire pour la suite des opérations.
Protocole
- Placer les échantillons d’ADN pendant 3 min à 60-65°C.
- Les transférer sur de la glace pendant 2-3 min.
Ces deux opérations sont destinées à empêcher la ligation de fragments possédant éventuellement des extrémités cohésives.
Pour le remplissage de puits de 10-12 mm de large bien adaptés à un usage pédagogique, une proportion de 5 µL de colorant de charge pour 10 à 15 µL d’ADN convient (de façon à obtenir de 1 à 4 µg d’ADN par puits). - Mélanger le colorant de charge et l’ADN sur un morceau de parafilm et prélever le mélange avec une micropipette réglée sur le volume approprié en changeant de cône à chaque prélèvement.
- Remplir les puits en faisant attention de ne pas déchirer le fond du gel avec la pointe de la pipette.
- Placer le support avec le gel chargé dans la cuve d'électrophorèse en positionnant les puits du côté de la cathode (pôle noir).
- Remplir la cuve de tampon TBE (réutilisable plusieurs fois) en versant délicatement et très lentement lorsque le gel commence à être recouvert pour éviter les fuites d’ADN vers le tampon.
- Fermer la cuve, brancher les fils et mettre sous tension.
- Laisser migrer jusqu’à ce que le colorant de charge arrive à proximité du bord du gel (environ 55 min à 100 V pour un gel de 80 mm dans une minicuve).
- Couper l’alimentation, débrancher les connections et récupérer le gel dans son support.
Attention au transport : le gel glisse très facilement de son support.

Noter la migration du colorant de charge de la cathode vers l'anode.
Révélation des bandes d’ADN
Attention, le gel doit être manipulé avec soin car il est très fragile.
- Recouvrir le gel avec le colorant.
- Laisser agir de 1 h à une nuit puis vider le colorant (qui peut être réutilisé). Les bandes d’ADN apparaissent en bleu.
Les gels sont observables à ce stade mais on peut vouloir décolorer le fond pour améliorer la qualité des photographies. - Rincer dans l’eau distillée pendant quelques heures en changeant l’eau de temps en temps.
- Photographier le gel avec un appareil numérique ou le scanner.
Les clichés ci-dessous, réalisés avec un scanner, présentent les résultats obtenus pour deux tailles de gels d'agarose.
Noter que le contraste des images a été amélioré par traitement numérique des images afin d'éviter la perte d'informations résultant de la prise de vue. En effet, les bandes les moins marquées mais néanmoins visibles à l'oeil nu ne sont parfois pas détectées par l'appareil de prise de vue.

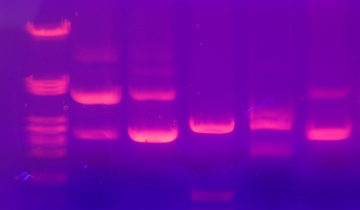
A gauche : minigel 8 x 6,5 cm coloré par le bleu de Nile.
A droite : gel 15 x 10 cm coloré par le bleu de Nile.
Pistes du minigel :
| 1 | 2 | 3 | 4 |
| ADN génomique de lentille | Marqueurs de taille | ADN de phage lambda digéré par EcoRI et HindIII | ADN de phage lambda digéré par HindIII |
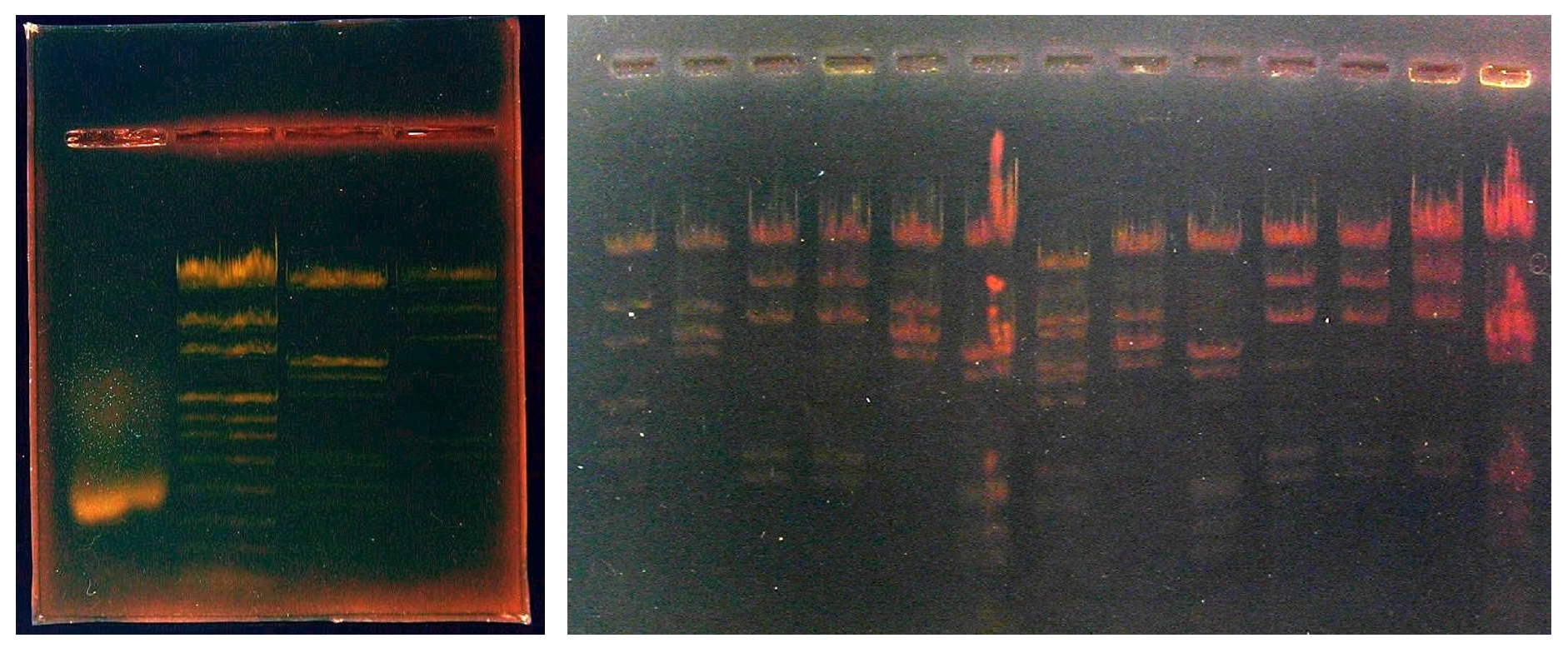
Pistes du gel 15 x 10 : (gel destiné à évaluer la qualité de l'ADN conservé au laboratoire depuis parfois plusieurs années avec décongélations - recongélations successives)
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
| pUC Mix (P) | EcoRI (P) | HindIII (P) | HindIII (S) | EcoRI (P) | EcoRI + HindIII (P) | BstII (S) | EcoRI (S) | EcoRI + HindIII (P) | HindIII (P) | HindIII (P) | HindIII (P) | EcoRI + HindIII (P) |
Puits 1 : marqueurs de taille : pUC mix (Promega)
Puits 2 à 13 : ADN de phage lambda digéré par diverses enzymes de restriction. Origine de l'ADN : Promega : P ; Sigma : S
Traitement des images et détection de bandes invisibles à l'oeil nu
Traitement simple
On peut améliorer la visibilité des bandes, notamment des moins marquées par le colorant, par des traitements numériques des images réalisés avec un logiciel de traitement d'images (PhotoEditor, Paint Shop Pro, Photoshop, etc.). On peut jouer sur la luminosité, sur le contraste, sur l'intensité relative des couleurs. On peut également obtenir une image en négatif souvent plus lisible comme ci-dessous.
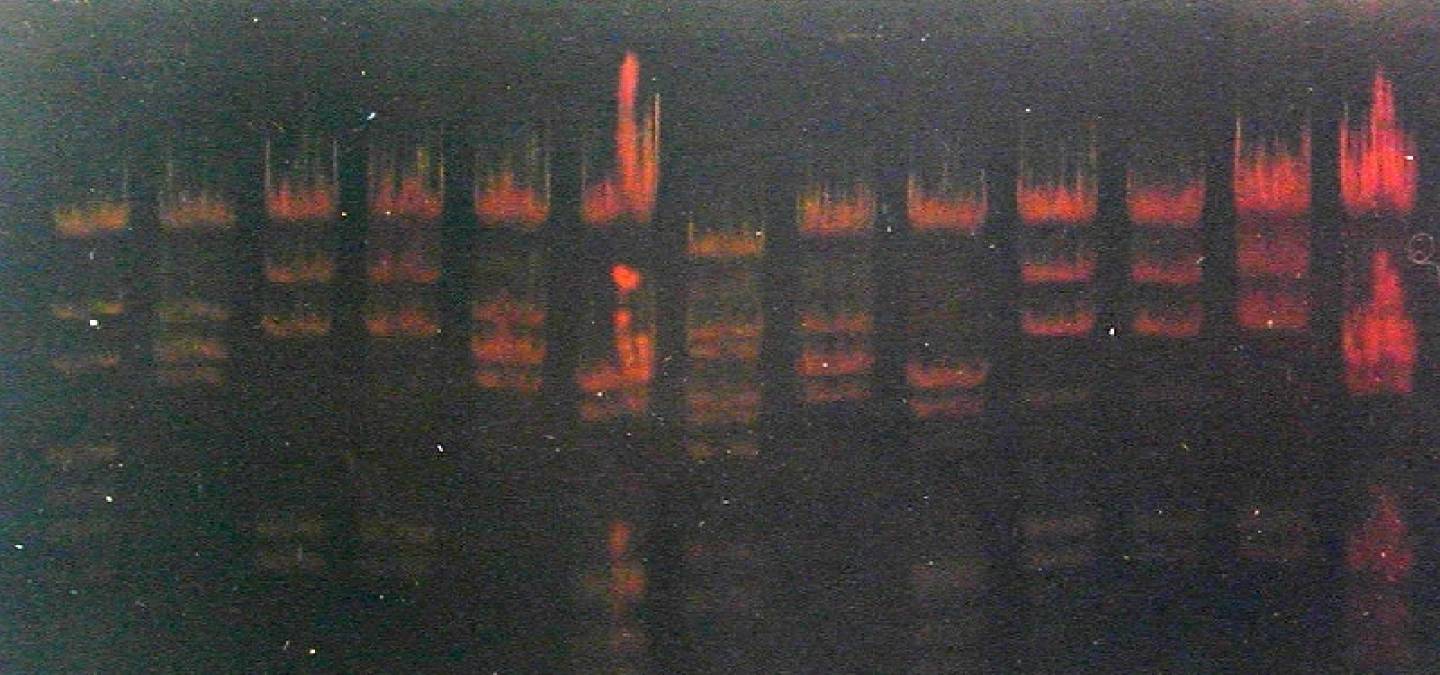
A gauche : minigel 8 x 6,5 cm
A doite : midigel 15 x 10 cm.
Détection de bandes invisibles par traitement numérique
Un logiciel de traitement d'images numériques, Digispec, donne la possibilité de mesurer l'intensité lumineuse de chaque pixel de l'image et celle de ses trois composantes colorées (télécharger le logiciel en projet Digispec de F. Borie). La représentation graphique de chacun de ces tracés à l'aide d'un logiciel tableur comme Excel permet alors de détecter des bandes invisibles à l'oeil nu en repérant les variations d'intensité sur les courbes. La figure ci-dessous présente les tracés obtenus avec la bande 2 du minigel ci-dessus (marqueurs de longueur, pUCMix).
Un autre logiciel, Mesurim, développé par J.F. Madre et librement téléchargeable sur le site de l'IFE (Institut Français de l'Education) permet également ce type de traitement. On se référera aux rubriques d'aide disponibles sur ce site pour son utilisation.
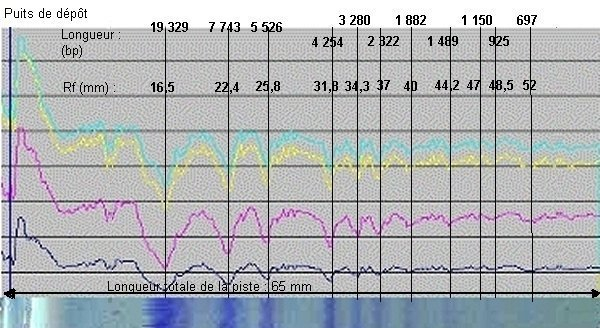
En bas de l'image, la piste 1 : marqueurs de taille de l'ADN [pUC Mix, Promega]
Sur cette piste 1, notez que 9 bandes seulement sont visibles à l'oeil nu, mais que les variations simultanées des quatre tracés densitométriques permettent d'identifier deux bandes supplémentaires (925 et 697 paires de bases respectivement).
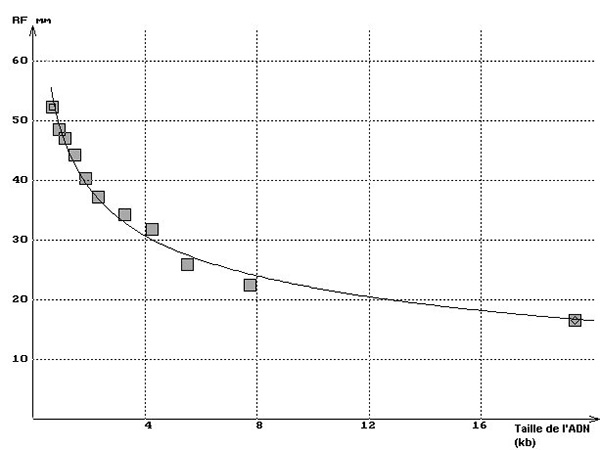
Les bandes étant identifiées, leur distance de migration est mesurée. Pour cela, l'image obtenue est affichée dans un logiciel de traitement d'image (PhotoEditor, Paint Shop Pro, Photoshop, etc.) et la distance de chaque bande au puits est mesurée directement avec la souris, le logiciel affichant le nombre de pixels correspondant. Les valeurs obtenues ont été reportées sur le graphique ci-dessus (Rf en mm). Connaissant la taille en paires de bases des fragments d'ADN donnée par le fournisseur, il est alors possible de construire la courbe exprimant la distance de migration en fonction de la taille des fragments d'ADN.
Construction de la courbe Distance de migration = f (taille de l'ADN)
La construction de la courbe nécessite, d'une part, de disposer des informations données par le fournisseur sur la taille des différents fragments de restriction présents dans le mélange et, d'autre part, des mesures réalisées précédemment sur l'image pour déterminer la distance de chaque bande à l'origine, c'est à dire au puits de dépôt.
N'importe quel tableur peut être utilisé pour entrer les données (taille des fragments en abscisses, distances de migration en ordonnées) et construire le graphique correspondant (Rf = a(1/log longueur) + b) qui constitue la courbe étalon permettant ensuite de déterminer la taille de fragments inconnus à partir de leur distance de migration. En outre, la construction de la courbe permet de vérifier la bonne adéquation des résultats obtenus avec la dimension des fragments de restriction indiquée par le fournisseur.
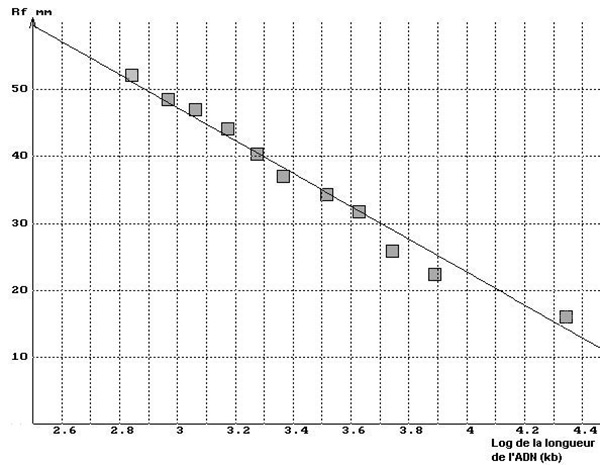
Sur le graphique ci-dessous à gauche on a superposé ainsi les résultats obtenus avec la piste 2 du minigel ci-dessus et la courbe théorique. Le tracé peut être aussi linéarisé en utilisant en abscisses le logarithme de la taille des fragments d'ADN comme le montre le graphique ci-dessous à droite ou en utilisant une échelle semi-logarithmique.
Détermination de la longueur de fragments d'ADN de taille inconnue
Les tracés ci-dessus peuvent être utilisés pour déterminer la taille de fragments d'ADN inconnus séparés par électrophorèse. Pour cela, la distance de migration des bandes de taille inconnue est mesurée sur le graphique. On a procédé de cette façon pour la piste 3 du minigel ci-dessus (ADN de phage lambda digéré entièrement par EcoRI et HindIII) après avoir identifié par analyse de l'image de la piste les bandes invisibles à l'oeil nu comme indiqué précédemment. La figure ci-dessous présente les résultats de cette opération.

Localisation des sites de restriction et prévision des résultats
Les séquences d'ADN conservées dans les banques de données internationales (voir à la fin de l'article) sont accessibles directement à partir des serveurs Web où elles peuvent être téléchargées aisément. Pour les exemples donnés ici, la séquence d'ADN du bactériophage lambda est téléchargeable directement à partir de cette page (au format html ou au format texte .txt utilisable directement avec un traitement de texte) en cliquant sur le lien correspondant.
Le génome du phage lambda comporte 48 502 paires de bases et a une masse moléculaire de 31,5 x 106 daltons.
- Séquence du génome du phage lambda (pour enregistrer le fichier, demander d'enregistrer la cible du lien).
Une fois la séquence chargée, on peut entreprendre de localiser les sites de restriction dans le génome du phage lambda dont l'ADN a été utilisé pour réaliser l'électrophorèse.
L'enzyme EcoRI reconnaît la séquence :
..GAATTC..
..CTTAAG..
L'enzyme HindIII reconnaît la séquence :
..AAGCTT..
..TTCGAA..
La fonction rechercher de tout traitement de texte ou du navigateur permet de localiser les sites de restriction si on lui indique la séquence à retrouver. Noter que la séquence ne doit pas comporter d'espaces entre les nucléotides pour que la fonction "rechercher" puisse trouver tous les sites de restriction. Dans ces conditions, la recherche de la séquence GAATTC conduit à identifier cinq sites de restriction situés respectivement à 21226, 26104, 31747, 39168, 44972 générant six fragments. De la même façon, la recherche de la séquence AAGCTT conduit à identifier six sites de restriction situés respectivement à 23130, 25157, 27479, 36895, 37459, 44141, générant sept fragments. Il est intéressant d'essayer de prévoir le nombre de fragments produits théoriquement par l'action simultanée des deux enzymes et de le comparer au nombre de fragments obtenus lors de l'électrophorèse.
Informations pratiques
Solutions
- Tampon TBE (Tris borate EDTA) pH 8,3
Pour 1000 mL d’eau distillée :
Tris.HCl [tris(hydroxyméthyl)aminométhane] 90 mmol.L-1 : 10,89 g
Acide borique 90 mmol.L-1 : 5,56 g
EDTA 2 mmol.L-1 : 0,74 g (sel disodique de l’acide éthylène diamine tétraacétique)
- Tampon TE (Tris EDTA) pH 7,6
Pour 100 mL d’eau distillée :
- Tris.HCl [tris(hydroxyméthyl)aminométhane] 1 mol.L-1 : 12,1 g
- EDTA (sel disodique de l’acide éthylène diamine tétraacétique) 0,1 mol.L-1 : 3,72 g
- Colorant de charge pH 8
Pour 10 mL d’eau distillée :
- Bleu de bromophénol 3 mmol.L-1 : 0,2 g
- Saccharose 1,5 mol.L-1 : 5,1 g
- Tris. HCl 10 mmol.L-1 : 0,01 g
- Solution de coloration pour les gels
- Solution stock (concentrée 100 fois) à conserver au réfrigérateur : Dissoudre 10 mg de bleu de Nile dans 5 mL d’eau distillée.
- Solution de coloration : 1 mL de la solution stock pour 100 mL d’eau distillée.
Selon la taille de la cuve de coloration, préparer 200 à 300 mL de la solution de coloration.
Approvisionnement
- Cuves à électrophorèse : Pierron, Bio-Rad, Bioblock
- Produits chimiques : SIGMA. L'Isle d'Abeau Chesnes BP 701 - 38297 Saint Quentin Fallavier
- ADN : EUROMEDEX. BP 80 - 67460 Souffelweyersheim
- Séquences d'ADN : EMBL, Genbank (Pour le génome du phage lambda, numéros d'accession : M17233, M24325, V00636)