

Certains traits persistent dans les populations alors qu'ils semblent « inutiles ». Le mamelon masculin en est un exemple et permet d'illustrer la notion de contrainte de construction de l'organisme.
Pour un public non spécialisé, l’évolution peut être perçue comme allant vers le progrès et la complexification. Ce n’est pas vrai, certaines adaptations peuvent conduire à des régressions et à des simplifications. Par exemple, les pattes arrières des baleines ont disparu et les yeux des taupes et des poissons cavernicoles ont régressé jusqu’à perdre leur fonction visuelle.
La question de la dégénérescence des organes inutilisés a fait l’objet de houleux débats chez les évolutionnistes. La question étant de comprendre pourquoi certains avaient totalement disparu et d’autres, non.
Si, dans un nouvel environnement, avoir une mauvaise vue n’est plus un handicap, les allèles responsables de ce caractère se répandent progressivement dans la population. Mais si des yeux inutiles persistent, c’est que le coût de leur élimination serait supérieur au coût de leur maintien. L’évolution optimise toujours le ratio coût sur bénéfice.
La question du maintien d’un mamelon1 chez le mâle Homo sapiens relève des mêmes interrogations, mais dans un registre totalement différent. Il faut comprendre que les mâles et les femelles ne sont pas des espèces distinctes, ils n’ont pas été façonnés séparément par la sélection naturelle. Leurs embryons sont similaires et les ébauches des organes sexuels sont identiques. Chez l’embryon humain la première différenciation sexuelle a lieu lors de la quatrième semaine au niveau de la crête génitale qui se transformera soit en ovaire, soit en testicule selon des déterminants chromosomiques et génétiques. Chez les mâles (XY), le canal de Wolff se transforme en épididyme à l’arrière et en canal déférent à l’avant. Chez les femelles (XX), le canal de Müller se différencie en oviductes, utérus, canal cervical et vagin. Le gène SRY joue un rôle central dans la masculinisation en induisant la production de testostérone. Ensuite, les hormones modifient le développement des organes. Le testicule produit la testostérone et l’hormone antimüllerienne (AMH) qui entraîne la dégénérescence du canal de Müller. Les faibles taux de testostérone chez les femelles entraînent la dégénérescence du canal de Wolff. Les caractères sexuels secondaires, c’est-à-dire l’apparence « homme » ou « femme » sont également soumis à l’influence de ces hormones produites par les gonades différenciées.
Les lignes mammaires se forment au 11ᵉ jour après fécondation, c’est-à-dire bien avant la différenciation de l’appareil reproducteur. Contrairement aux organes reproducteurs, elles ne sont pas ou très peu soumises aux influences hormonales. Chez le mâle, le développement mammaire embryonnaire précoce est identique à celui de la femelle. La crête mammaire s’épaissit à partir de la 4e semaine, puis laisse place à deux bourgeons mammaires à partir de la 6e semaine. Ce mode de développement explique la possibilité de mamelons surnuméraires, si la crête disparaît en laissant des bourgeons excédentaires.
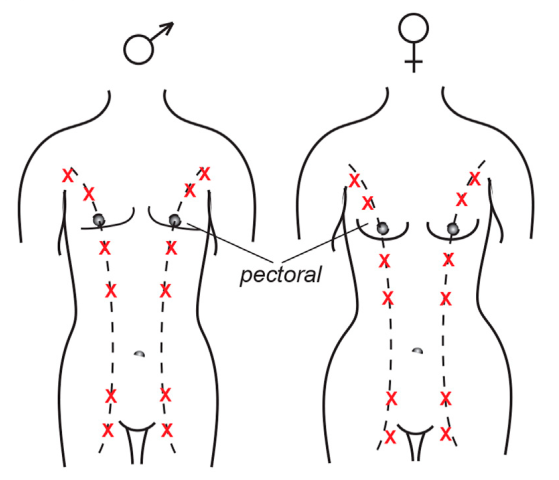
Chez l’être humain, les crêtes mammaires apparaissent entre la 4e et la 5e semaines de gestation. Elles s’étendent des aisselles à l’aine mais régressent pour ne laisser place, chez la plupart des individus, qu’à deux mamelons. Cependant, une fraction non négligeable de la population (allant de 0,2 à 6 % en fonction des régions du monde) présente un ou des mamelons surnuméraires (en particulier aux emplacements indiqués par des croix rouges).
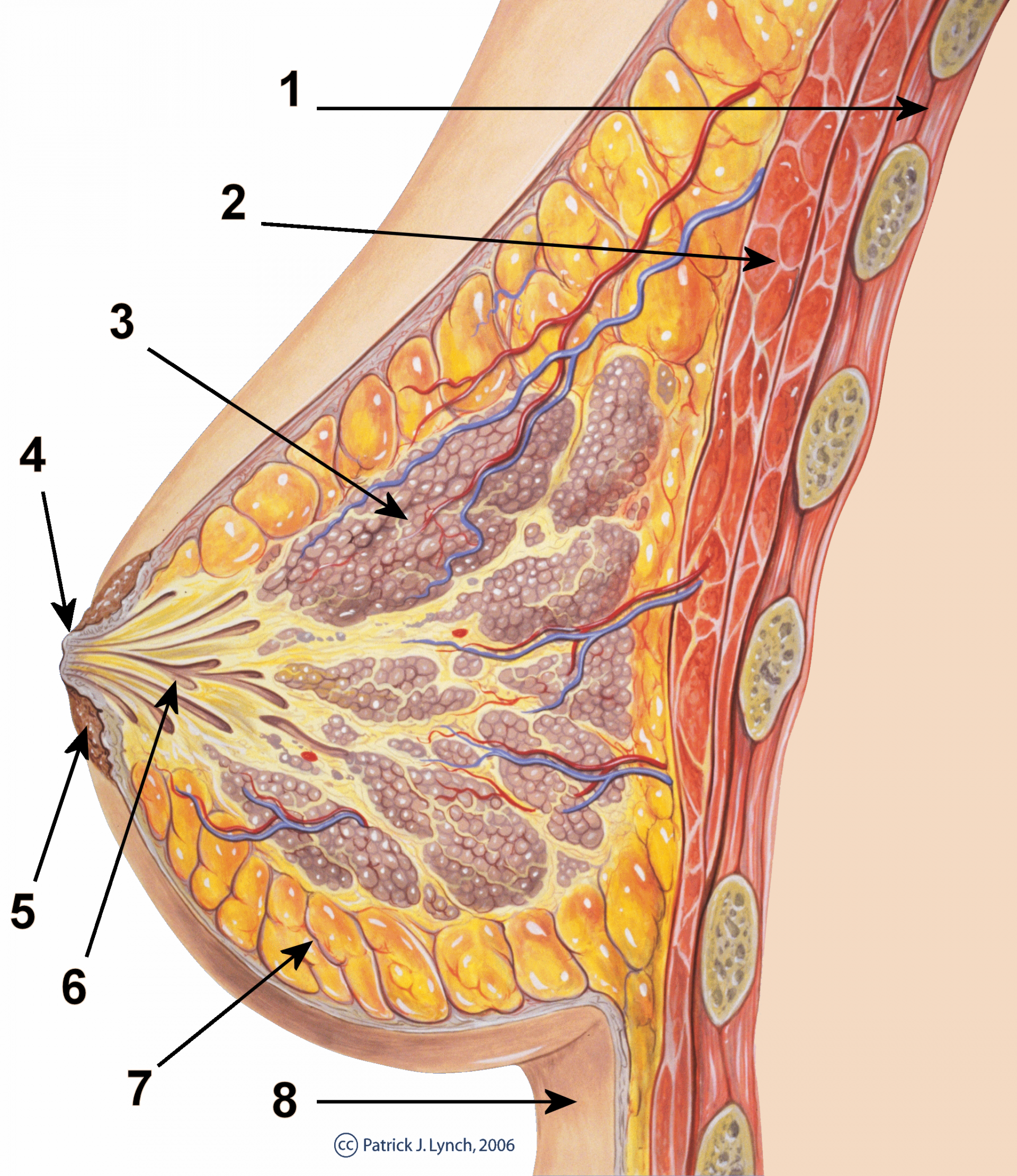
1 : cage thoracique ; 2 : muscles pectoraux ; 3 : lobules ; 4 : surface du mamelon ; 5 : aréole ; 6 : conduit galactophore ; 7 : tissu adipeux ; 8 : peau.
La glande mammaire reste quiescente jusqu’à la puberté. Et la structure anatomique qui permet l’allaitement n’est atteinte qu’après le premier accouchement. Le sein de la femme est véritablement un organe à développement très tardif en termes évolutionnistes. Avant la puberté, la glande mammaire subit une croissance très lente par ramifications successives de galactophores (canaux de la lactation) non fonctionnels.
En biologie de l’évolution, on dit qu’un trait est « robuste » lorsque sa différenciation est très ancienne dans la phylogenèse, et que sa structure et son adaptation semblent stables et achevées. Les gonades mâles et femelles, ainsi que l’utérus sont des traits robustes. Les hommes n’auront jamais de grossesse quelles que soient les prouesses chirurgicales que l’on puisse imaginer. Inversement, le sein et le mamelon ne sont pas robustes. Sous l’effet de produits chimiques ou de certaines hormones, notamment œstrogène et prolactine, ou d’une baisse de la testostérone, le glande mammaire des hommes peut se développer ; c’est ce que l’on nomme une gynécomastie.
Les nouveau-nés masculins ont très souvent une gynécomastie liée au passage des œstrogènes placentaires dans la circulation. Par ailleurs, la moitié des garçons présentent ce trait, qui peut durer parfois deux ans, au moment de la puberté. Plusieurs médicaments, ainsi que le cannabis peuvent provoquer des gynécomasties. Ce caractère s’observe également chez les hommes âgés ou traités avec des anti-androgènes pour un cancer de la prostate.
Il n’est pas tout à fait irréaliste d’envisager qu’un homme puisse allaiter. Il faudrait par exemple injecter de la prolactine pour stimuler les canaux galactophores. Il existe au moins une espèce de mammifères où les mâles allaitent, il s’agit de la chauve-souris Dyacopterus spadiceus.
En résumé, la présence d’un mamelon et, plus encore, d’une glande mammaire chez l’homme adulte résulte d’une contrainte de construction puisque, lors du développement embryonnaire, il est mis en place bien avant la différenciation sexuelle.
Références
- Gould, Stephen Jay. La foire aux dinosaures: réflexions sur l’histoire naturelle. Éd. du Seuil, 1997.
- Veltmaat, Jacqueline M., et al. « Mouse Embryonic Mammogenesis as a Model for the Molecular Regulation of Pattern Formation ». Differentiation, vol. 71, nᵒ 1, janvier 2003, p. 1‑17. https://doi.org/10.1046/j.1432-0436.2003.700601.x.



