

L’Homme de Luçon, Homo luzonensis, est une espèce d’Hominine récemment décrite à partir de restes fossiles mis au jour dans la grotte de Callao, aux Philippines. L’étude des quelques ossements disponibles montre une combinaison originale de caractères, certains à l’état dérivé, d'autres à l'état ancestral par rapport aux autres représentants du genre Homo. De récentes analyses phylogénétiques permettent de discuter la place d’Homo luzonensis au sein de la lignée humaine. De nombreuses questions restent cependant ouvertes, par exemple sur l’origine de cette espèce, sur son mode de locomotion ou encore sur la façon dont elle a pu arriver sur l’île de Luçon.
Des découvertes spectaculaires, effectuées depuis à peine une vingtaine d’années, ont permis de montrer que la complexité de l’évolution et des migrations humaines en Asie de l’Est et du Sud-Est a été nettement sous-estimée jusqu’à aujourd’hui. Ainsi, des fossiles, mais aussi des séquences génétiques, ont récemment permis d’identifier plusieurs nouvelles espèces du genre Homo aujourd’hui éteintes, mais qui étaient contemporaines de notre propre espèce, Homo sapiens, dont l’origine est estimée à 200 à 300 000 ans environ. En plus d’Homo neanderthalensis, il faut maintenant compter avec les « Dénisoviens » [1], Homo longi1 [2], Homo floresiensis [4] et Homo luzonensis [5]. Ces deux dernières espèces, identifiées respectivement sur les îles de Florès en Indonésie et de Luçon (ou Luzon en Tagalog) aux Philippines étaient, chacune à leur manière, marquées par l’endémisme insulaire et présentaient, en plus de leur très petite taille, de nombreuses caractéristiques morphologiques très surprenantes pour des espèces aussi récentes du genre Homo. Nous vous proposons ici de partir à la découverte de H. luzonensis, de la mise au jour des fossiles lors des fouilles jusqu’à leur analyse et la publication de la nouvelle espèce en 2019 [5], puis nous vous présentons les résultats des toutes premières analyses phylogénétiques menées en 2021 sur les restes fossiles afin de tester les différents scénarios possibles sur la place de cette espèce dans l’arbre des hominines [6].
La découverte des fossiles dans la grotte de Callao, île de Luçon, Philippines
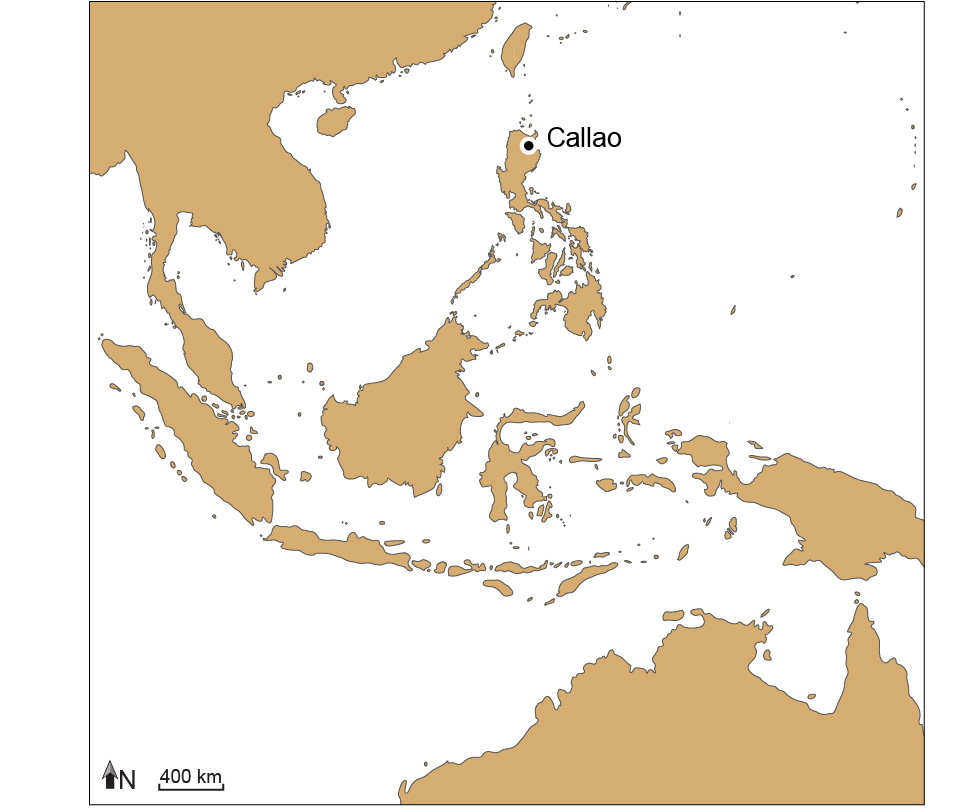
Fond de carte adapté par Florent Détroit à partir de Voris, H. K. 2000. Maps of Pleistocene Sea Levels in South East Asia: Shorelines, River Systems, Time Durations. Journal of Biogeography 27:1153-1167. © 2000 Field Museum of Natural History, Chicago, Illinois USA
Les ossements et les dents fossiles attribués à la nouvelle espèce H. luzonensis ont été découverts lors de plusieurs campagnes de fouilles archéologiques menées depuis 2003 dans la grotte de Callao (Callao Cave), aux Philippines (Figure 2). Callao se trouve dans un massif calcaire d’âge Miocène appelé « massif de Callao » et situé dans la région de la vallée de Cagayan, au nord-est de l’île de Luçon, la grande île où se trouve la capitale actuelle du pays, Manille (Figure 1). Ce petit massif calcaire renferme des centaines de grottes, dont certaines, comme Callao, s’ouvrent le long des falaises creusées par la rivière Pinacanauan.

Plusieurs de ces grottes renferment du matériel archéologique identifié lors de prospections ou de fouilles correspondant à des périodes « récentes » de la préhistoire, c’est-à-dire de la fin de la période Holocène (s’étendant d’il y a environ 12 000 ans à aujourd’hui) et en particulier du Néolithique (période à laquelle les humains domestiquent les animaux et/ou les plantes) et des périodes historiques « précontact » (c’est-à-dire avant l’arrivée au XVIe siècle des premiers colons européens aux Philippines). À ce jour, la grotte de Callao est la seule de la région à avoir livré des vestiges préhistoriques bien plus anciens : des outils de pierre taillée découverts dans un niveau archéologique daté de – 25 000 ans, mais aussi et surtout les restes fossiles humains qui ont permis la description de la nouvelle espèce H. luzonensis. Ces fossiles ont été mis au jour dans une couche sédimentaire située à plus de trois mètres de profondeur et préservée uniquement dans certaines zones de la grotte. Cette couche fossilifère a été fouillée pour la première fois en 2007 et de nombreux fragments d’os et de dents d’animaux y ont été découverts. Parmi les ossements se trouvait un petit os de pied (un métatarsien) de morphologie humaine, mais de petite taille et présentant plusieurs caractères assez différents de ceux d’un Homo sapiens [7]. Cette découverte initiale a été complétée par d’autres fossiles mis au jour lors des fouilles de 2011 et de 2015. Les datations indiquent que ces fossiles sont vieux de plus de 50 000 ans et les descriptions et analyses comparatives des fossiles nous ont permis de les attribuer en 2019 à une nouvelle espèce : Homo luzonensis.
Principales caractéristiques de l’espèce H. luzonensis
L’assemblage fossile
En tout et à ce jour, 13 éléments fossiles différents ont été découverts et attribués à H. luzonensis. Il s’agit de dents, d’os des mains et des pieds et d’un fémur partiel correspondant à au moins trois individus différents. En effet, l’assemblage comporte une diaphyse1 de fémur d’enfant (Figure 3d). Bien que de petite taille par rapport à un fémur de H. sapiens actuel adulte, son aspect extérieur ne permettait pas de savoir s’il appartenait à un H. luzonensis adulte de très petite stature ou à un enfant. Mais l’analyse d’images obtenues par microtomographie (au moyen d’un scanner à rayons X permettant d’obtenir des images à plus haute résolution qu’un scanner médical classique) a confirmé que la structure de cet os correspond bien à celle d’un enfant en cours de croissance. Parmi les autres éléments, deux troisièmes molaires supérieures droites sont présentes (Figures 3a et 3c). Elles appartenaient nécessairement à deux individus différents (car nous n’avons qu’une seule troisième molaire supérieure du côté droit) et ces deux individus étaient adultes car chez les humains (et tous les grands singes en général) les troisièmes molaires apparaissent à la fin de l’adolescence ou au début de l’âge adulte (ce sont, en langage courant, les fameuses « dents de sagesse »). Tous ces éléments fossiles correspondent donc à au moins deux adultes et un enfant.
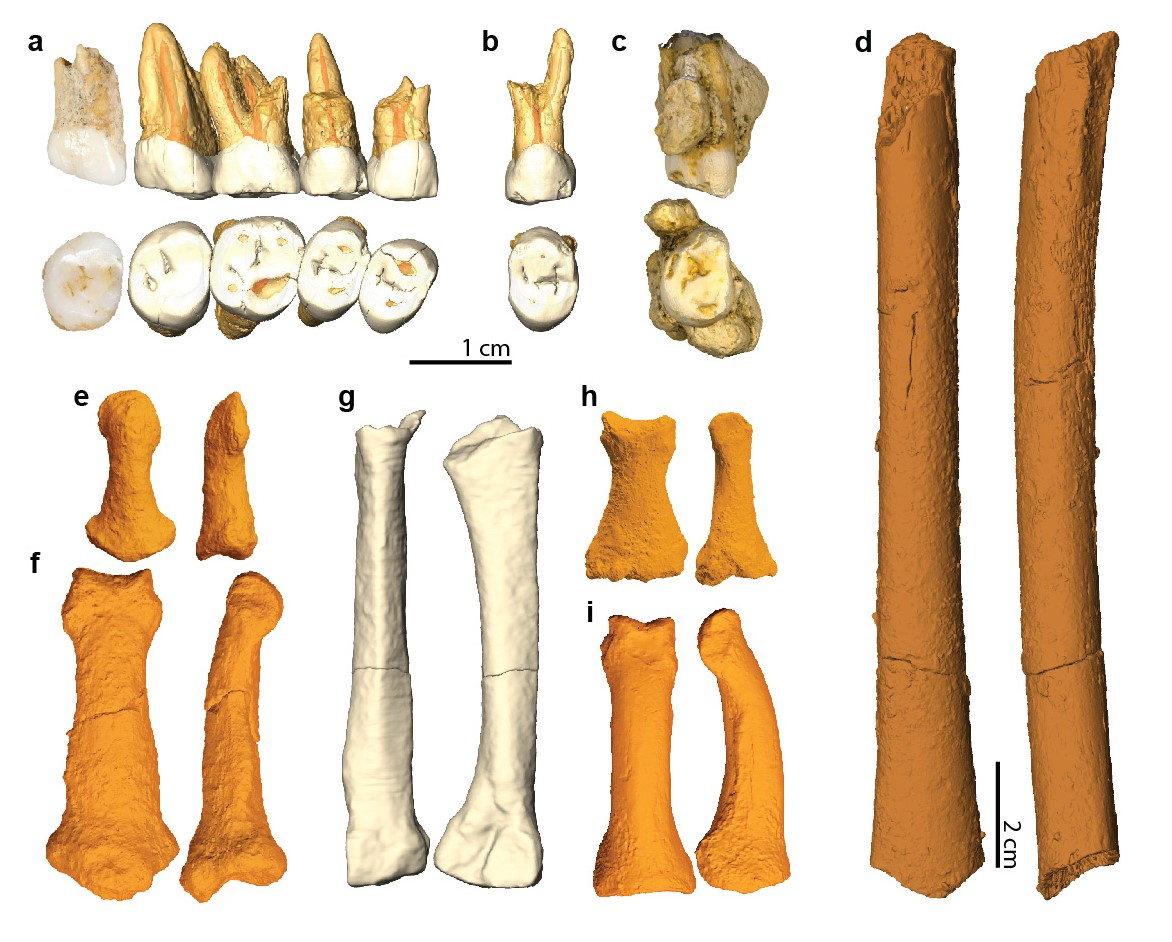
a-c : dents maxillaires (a : prémolaires et molaires CCH6 ; b : prémolaire CCH8 ; c : molaire CCH9) ; d : fémur juvénile CCH7 (incomplet) ; e-f : os des mains (e : phalange distale CCH5 ; f : phalange intermédiaire CCH2) ; g-i : os des pieds (g : métatarsien CCH1 ; h : phalange intermédiaire CCH3 ; i : phalange proximale CCH4).
Les noms donnés aux fossiles (CCH1, CCH2, etc.) signifient Callao Cave Hominin et les numéros correspondent à l’ordre de découverte des spécimens lors des fouilles ou, plus exactement, suite à l’examen des restes après les fouilles. Par exemple, les cinq dents du fossile CCH6 ont été découvertes dissociées, mais l’examen des facettes de ces dents au contact les unes des autres a permis de montrer qu’elles coïncidaient parfaitement.
Alors que tous ces fossiles ont été retrouvés dissociés les uns des autres, nous savons que cinq des dents supérieures droites (deux prémolaires et trois molaires, Figure 3a) appartenaient à un même individu car les facettes de contact entre les dents correspondent parfaitement. Il est en revanche impossible d’aller plus loin et de savoir si, par exemple, la phalange manuelle intermédiaire de main (Figure 3f) et la phalange proximale de pied (Figure 3i) proviennent d’un même individu puisqu’il n’y a bien sûr pas de contact anatomique entre ces deux éléments. Ce sont donc les restes d’au moins trois individus différents qui ont été mis au jour dans la grotte de Callao.
Les dents d’Homo luzonensis
Seuls sont légendés les éléments utiles à la compréhension du texte principal.
Plus de la moitié des éléments fossiles (7 sur 13) attribués à H. luzonensis sont des dents (Figure 4). C’est une situation fréquente en paléontologie dans la mesure où les dents étant composées de tissus très minéralisés (en particulier l’émail dentaire), elles constituent les éléments les plus résistants du squelette des vertébrés, hominines compris. Elles sont donc souvent mieux conservées que les os, plus fragiles. Les dents se fossilisent plus facilement, résistent mieux à l’érosion et aux autres processus géologiques « destructeurs » et elles sont aussi plus aisément reconnues et déterminées par les paléontologues. Dans un ensemble de fossiles, il est en effet plus facile de reconnaître une dent humaine, même si ses racines sont cassées, qu’un os fragmentaire quand les portions les plus reconnaissables sont absentes ou fortement endommagées (il est souvent difficile, voire impossible, de déterminer l’espèce à laquelle appartenait un petit fragment d’os fossile). Les paléontologues spécialistes des vertébrés fossiles, paléoanthropologues compris, considèrent généralement les dents comme le « meilleur matériel fossile » pour étudier, comparer et déterminer de nouvelles découvertes car, celles-ci se conservant mieux, elles sont représentées en très grand nombre dans les collections, pour toutes les espèces. Ainsi, le rythme et les modalités d’évolution sur le temps long de la morphologie externe et interne des dents des hominines sont très bien connues, ce qui en fait le matériel fossile de prédilection de quantité de paléoanthropologues.
Dans le cas de H. luzonensis, nous avons pu comparer les dents découvertes dans la grotte de Callao avec celles de pratiquement toutes les espèces connues des genres Homo (nombreuses espèces fossiles et une seule espèce actuelle, H. sapiens), Paranthropus et Australopithecus (espèces éteintes pour ces deux genres), mais aussi avec celles de nombreux primates non-humains afin d’écarter toute confusion potentielle. Cette étude de la morphologie et de la taille des dents a permis de montrer qu’elles appartenaient, sans aucun doute possible, à un hominine et de relever un ensemble de caractéristiques tout à fait particulières et sensiblement différentes de celles présentées par les espèces du genre Homo connues en Eurasie à la même période ou à des périodes un peu plus anciennes (c’est-à-dire H. sapiens, H. neanderthalensis, « Denisoviens », H. floresiensis et H. erectus). D’une manière générale, les dents de H. luzonensis sont petites, même très petites, et leurs caractéristiques présentent un contraste très intéressant entre les prémolaires et les molaires (Figure 5). Les prémolaires sont beaucoup plus petites que celles de H. erectus, légèrement plus petites que celles de H. sapiens et un peu plus grosses que celles de H. floresiensis. Au niveau de la couronne d’émail et des racines, elles présentent des caractères très primitifs, généralement rencontrés chez des hominines bien plus anciens (Australopithèques et H. habilis) mais absents ou très rares chez les espèces plus récentes, notamment H. sapiens. Les molaires sont très petites : beaucoup plus petites que celles de H. erectus et celles de H. sapiens et même un peu plus petites que celles de H. floresiensis, qui était jusqu’alors l’espèce du genre Homo avec les plus petites dents. Et contrairement à ce qui est observé sur les prémolaires, les molaires présentent des caractères morphologiques très « modernes1 », très dérivés, c’est-à-dire ressemblant fortement à ceux que l’on observe chez H. sapiens.
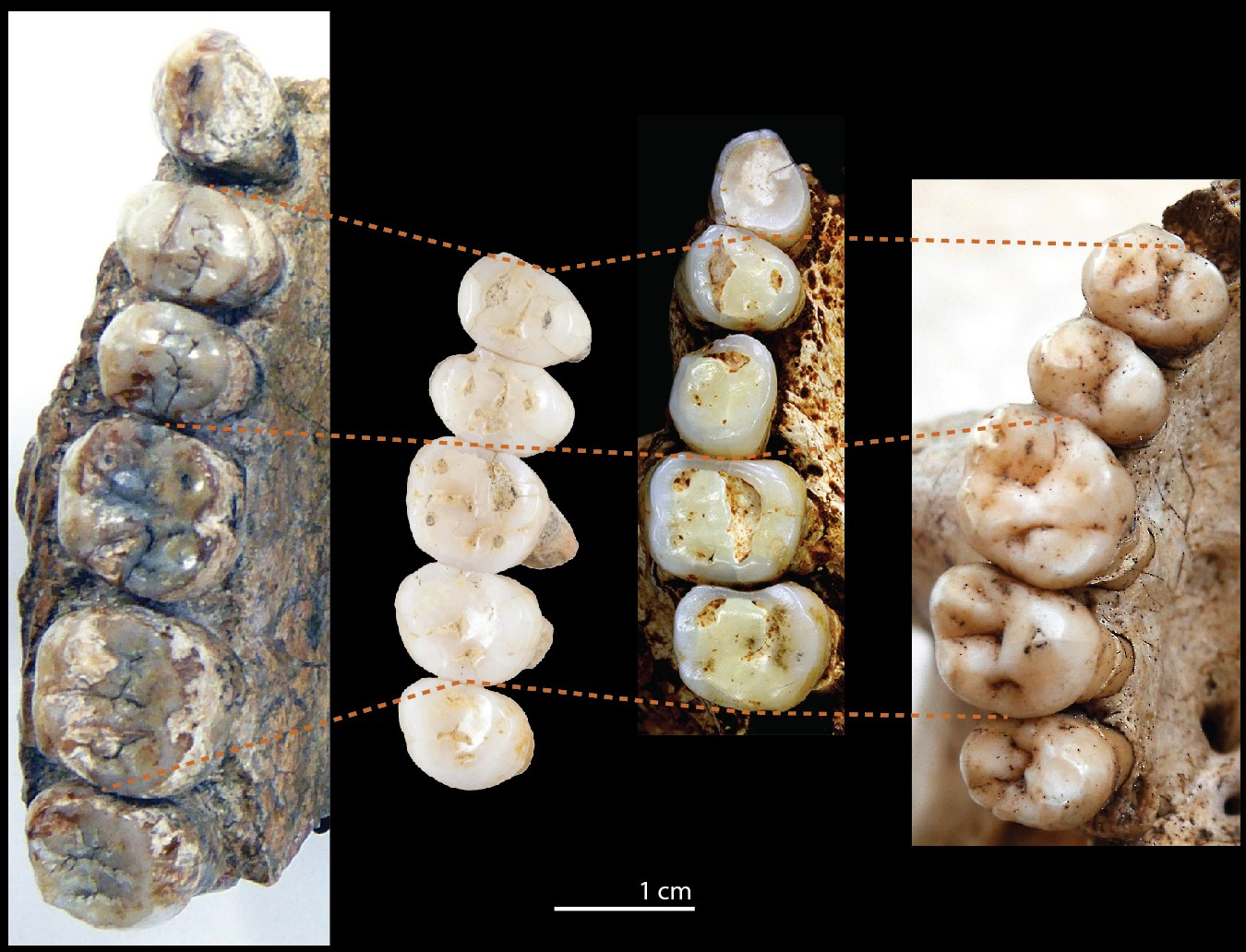
De gauche à droite : Homo erectus, Homo luzonensis, Homo floresiensis et Homo sapiens. Homo erectus de Java : Sangiran 4, de la canine à la troisième molaire ; Homo luzonensis : CCH6, de la première prémolaire à la troisième molaire ; Homo floresiensis : LB1 (vue en miroir des dents gauches), de la canine à la deuxième molaire (remarque : cet individu présente une rotation anormale à 90° de sa deuxième prémolaire) ; Homo sapiens : individu récent, de la première prémolaire à la troisième molaire. Le trait pointillé orangé permet de comparer les proportions relatives des deux prémolaires et des deux premières molaires, conservées chez tous les spécimens.
La face occlusale des dents est celle qui, directement en contact avec les aliments, permet de les triturer.
Crédits : montage : F. Détroit ; photographies : Senckenberg Research Institute et Natural History Museum Frankfurt, C. Zanolli ; National Museum of the Philippines et Callao Cave Archaeology Project, F. Détroit ; Pusat Penelitian Arkeologi Nasional, Creative Commons Kaifu et al. 2015 ; C. Zanolli.
Ce contraste étonnant entre les caractéristiques primitives des prémolaires et dérivées des molaires est d’autant plus intéressant que l’on sait qu’il n’est pas dû à un simple polymorphisme (c’est-à-dire à des différences marquées d’un individu à un autre) puisqu’une partie de ces dents appartenaient bien à un seul et même individu. Nous avons ensuite fait appel à la « paléoanthropologie virtuelle » pour aller plus loin dans les comparaisons et notamment pour analyser la morphologie de la cavité pulpaire et de la jonction émail-dentine. L’utilisation de la microtomographie permet en effet d’enlever virtuellement la couronne d’émail, porteuse de caractères très intéressants mais souvent endommagée ou usée, afin d’accéder à la surface de la jonction entre l’émail et la dentine. Celle-ci, protégée par la couronne d’émail, est toujours mieux conservée et on sait depuis plusieurs années qu’elle présente des caractéristiques de morphologie et de forme très précieuses car très pertinentes pour les comparaisons et les analyses taxonomiques et phylogénétiques. Les résultats obtenus sur la morphologie interne des dents de H. luzonensis sont très complémentaires de ceux obtenus sur la morphologie externe puisqu’ils indiquent des affinités plus marquées avec les H. erectus asiatiques plus anciens qu’avec leurs contemporains H. sapiens et H. neanderthalensis. Cette observation a d’ailleurs été renforcée par les résultats de nouvelles analyses de la morphologie dentaire que nous avons publiés en 2022 [8].
Les os des mains et des pieds
Plusieurs os des mains (deux phalanges) et des pieds (deux phalanges et un métatarsien) de H. luzonensis ont été découverts dans la grotte de Callao (Figure 3 e-i). Ces os de petites dimensions sont assez peu fréquemment retrouvés lors de fouilles et, à l’inverse des dents, ils ne sont pas très abondants dans les collections. La morphologie des mains et des pieds de plusieurs espèces d’hominines reste donc mal, voire très mal connue, alors que ce sont pourtant des ensembles anatomiques clés pour mieux comprendre leurs aptitudes locomotrices et manipulatrices. La détermination de ces aptitudes est fondamentale en paléoanthropologie puisqu’une bipédie stricte et une forte dextérité manuelle sont généralement considérées comme caractéristiques du genre Homo.
Les os du pied de H. luzonensis sont particulièrement intéressants car ils présentent des attributs morphologiques d’apparence primitive, jusqu’alors inconnus dans le genre Homo. Ainsi, le métatarsien est de petite taille et présente des proportions très différentes de celles rencontrées chez H. sapiens. Cet os présente aussi une facette articulaire proximale (donc du côté de la cheville) qui est légèrement convexe alors qu’elle est rectiligne chez H. sapiens. Ce caractère pourrait permettre le mouvement de dorsiflexion s’effectuant au milieu du pied de nombreux primates non-humain (permettant de soulever le talon indépendamment du reste du pied), mais qui est absent ou extrêmement réduit chez H. sapiens dans la mesure où notre pied est très « rigide » du talon à la base des orteils, optimisant la stabilité au sol et la propulsion lors de la marche.
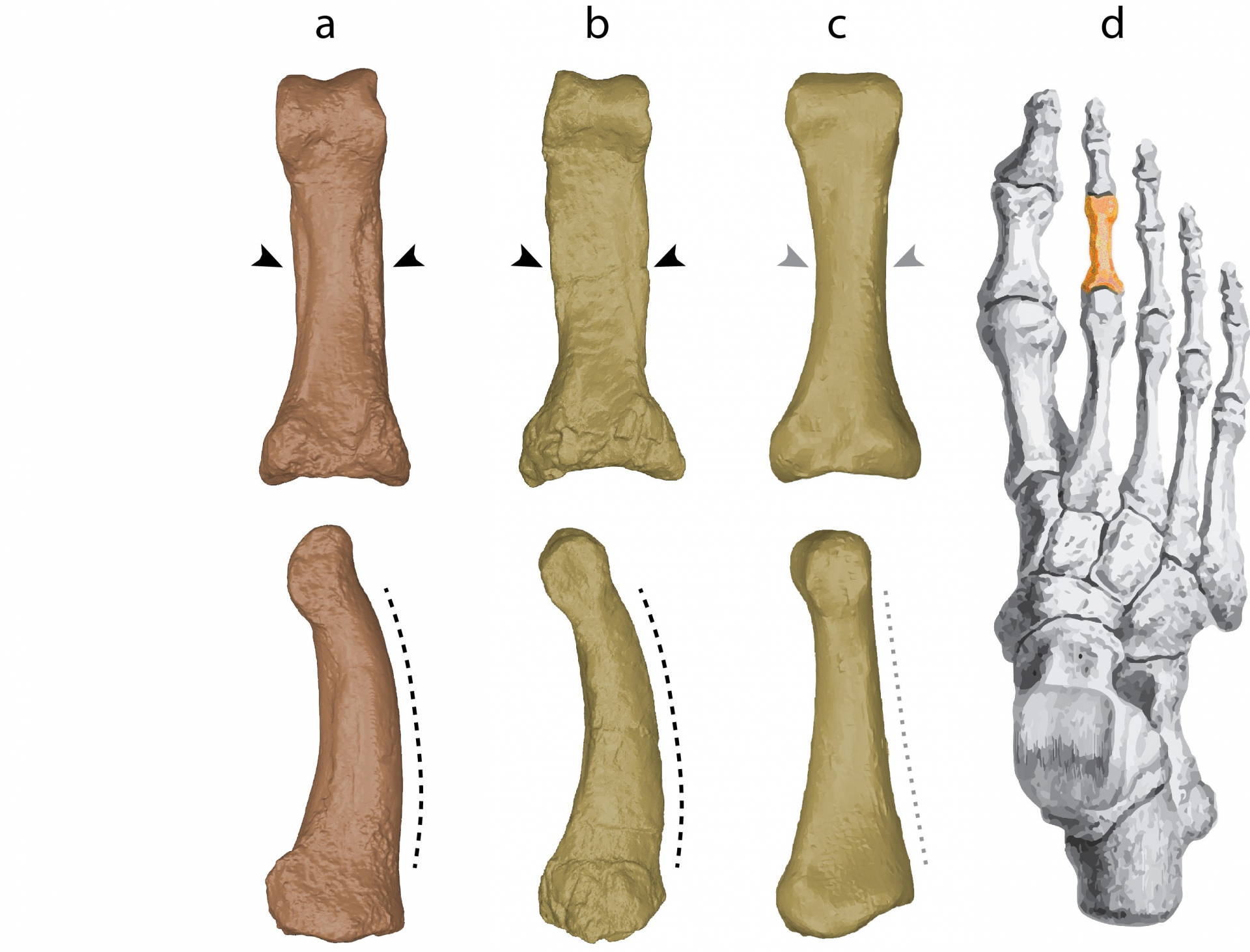
Les principales ressemblances entre H. luzonensis et A. afarensis sont indiquées en noir au niveau des flèches (forme générale en vue plantaire et développement des insertions pour les muscles fléchisseurs des pieds) et du trait pointillé (courbure longitudinale marquée en vue latérale) ; H. sapiens se distingue de ces deux espèces par une forme dite « en sablier » avec des insertions très réduites pour les muscles fléchisseurs des pieds en vue plantaire et un bord dorsal rectiligne en vue latérale. Ces trois phalanges sont des phalanges de pied de la rangée proximale (du doigt 2, 3 ou 4) dont l’emplacement est indiqué en orange (d).
Crédits : acquisition des modèles 3D et montage : Florent Détroit ; spécimens originaux et moulages du National Museum of the Philippines et du Muséum national d’Histoire naturelle.
Parmi les deux phalanges de pied qui ont été mises au jour, la phalange proximale (appartenant à la rangée de phalanges la plus proche de notre cheville) est la plus étonnante (Figure 6). Chez H. sapiens, cet os est en forme de sablier et présente un bord supérieur rectiligne, correspondant également à une anatomie du pied optimisée pour la marche bipède. D’après le registre fossile, il semble que cette morphologie soit commune à toutes les espèces du genre Homo. Pourtant, la phalange de H. luzonensis est incurvée longitudinalement et elle n’est pas en forme de sablier. En effet, sa portion centrale présente du côté de la voûte plantaire de larges zones d’insertions pour les muscles fléchisseurs des pieds. Ces caractéristiques (et d’autres comme la faible obliquité de la facette articulaire proximale) se retrouvent chez le genre Australopithecus et, de manière encore plus accentuée, chez les grands singes non-humains et on considère généralement qu’elles correspondent à des modes locomoteurs incluant une part plus ou moins importante de déplacements arboricoles. En effet, la locomotion des Australopithecus devait inclure une part de marche bipède au sol et une part de déplacements dans les arbres, dans des proportions qui restent très débattues et probablement variables d’une espèce d’australopithèque à l’autre [9,10]. Il est donc extrêmement surprenant de retrouver de tels caractères d’apparence archaïque sur les os des pieds de H. luzonensis. Il faut toutefois rester prudent quant à leur interprétation et il est impossible de savoir si H. luzonensis pratiquait une part de grimper dans les arbres avant d’avoir développé plus de travaux sur la biomécanique de son pied. Une chose semble cependant assez évidente : avec un pied constitué de la sorte, H. luzonensis n’était certainement pas un « excellent bipède », ou, pour l’exposer de façon plus rigoureuse, il devait pratiquer une marche bipède assez différente de celle de ses contemporains H. sapiens et H. neanderthalensis.
Les deux phalanges de main de H. luzonensis présentent également des informations précieuses. La petite phalange distale (située tout au bout d’un doigt) est de petite taille et présente une morphologie proche de celle que l’on retrouve chez les autres espèces du genre Homo. En revanche, la phalange manuelle intermédiaire (située au milieu d’un doigt) possède des caractéristiques assez singulières puisque sa morphologie et sa forme présentent à la fois des ressemblances et des différences avec celles de toutes les autres espèces d’hominines connues. La particularité la plus marquante de cette phalange est qu’elle est aplatie, mais aussi qu’elle est longue. Bien qu’il ne soit pas possible de savoir si cette phalange correspond à l’index, au majeur ou à l’annulaire (les trois plus grands doigts de la main), sa longueur importante est assez étonnante quand on la rapporte aux petites dimensions des autres éléments, notamment ceux des pieds. Tant que de nouveau fossiles n’ont pas été découverts, il ne faut toutefois pas déduire de cette observation qu’H. luzonensis avait nécessairement des petits pieds et de grandes mains, car ces différences de dimensions peuvent aussi être liées à des différences de taille entre individus.
Homo luzonensis dans la phylogénie
Hypothèses sur l’origine de H. luzonensis et de ses caractères
Les fossiles découverts et attribués à H. luzonensis présentent, comme nous venons de le décrire, une mosaïque de caractères anatomiques indiquant clairement qu’il s’agit bien d’un hominine et plus précisément d’une espèce du genre Homo. Mais comme nous l’avons vu, certains de ses caractères sont d’apparence très primitive chez les hominines et ils sont rares, voire absents, des autres espèces du genre Homo. Comment expliquer qu’une espèce du genre Homo qui vivait aux Philippines il y a à peine plus de 50 000 ans présente de telles ressemblances avec des espèces bien plus anciennes, y compris du genre Australopithecus qui vivaient en Afrique il y a plus de 3 millions d’années ? Deux grandes hypothèses phylogénétiques sont envisageables.
La première est que ces caractères archaïques ont été hérités directement de membres du genre Australopithecus, ce qui impliquerait que ces derniers seraient arrivés en Asie il y a plus de 2 millions d’années sans laisser de trace puisqu’à ce jour, sa présence hors d’Afrique n’est pas connue. Il ne faut cependant jamais oublier que dans nos disciplines, l’absence de preuve ne constitue pas la preuve de l’absence et que peut-être un jour prochain, des fossiles d’australopithèques seront découverts en Asie, même si cela semble aujourd’hui relativement peu probable étant donnés les très nombreux sites fouillés et l’abondance des fossiles mis au jour.
La seconde hypothèse est que les ancêtres de H. luzonensis se trouveraient du côté des H. erectus, présents partout en Asie de l’Est et du Sud-Est depuis au moins 1,6 millions d’années et jusqu’à environ 100 000 ans ainsi qu’indiqué par les très nombreux fossiles découverts en Chine et en Indonésie. Dans cette hypothèse, les caractères primitifs observés chez H. luzonensis, notamment au niveau des pieds, correspondraient à des réversions vers un état ancestral alors qu’ils auraient été hérités de H. erectus dans un état dérivé (le pied de H. erectus ressemblait bien plus au nôtre que le pied de H. luzonensis).
Pour tester ces hypothèses, des analyses phylogénétiques ont été menées en 2021, dans le cadre du travail de recherche de master 2 effectué par Pierre Gousset au MNHN [6].
L’approche cladistique
De nombreuses méthodes sont utilisées en paléoanthropologie pour tenter de reconstituer les relations de parentés entre taxons. L’approche cladistique a pour avantage de présenter de solides bases empiriques. En effet, la cladistique a pour principe de hiérarchiser les états de caractères en ancestraux et dérivés, et d’ainsi regrouper les taxons par les caractères qu’ils partagent à l’état dérivé, et qui résultent supposément d’une histoire commune plus longue entre membres de ces taxons qu'avec d'autres taxons [11].
Traitement des caractères
L’analyse cladistique commence par la construction d’une matrice, constituée de 66 caractères observables chez H. luzonensis ainsi que dans 15 autres taxons d’Hominines. Quatre espèces de grands singes ont été considérées comme groupes externes (ou extragroupes), permettant ainsi de polariser les caractères en fonction de leur état supposément ancestral.
Certains des caractères utilisés pour l’analyse phylogénétique étaient des caractères discrets (par exemple le nombre de racines sur une dent) qui étaient donc codés, dans la matrice, par un nombre limité d’états (notés 0, 1, 2, etc.). D’autres, comme les ratios de mesures anatomiques, constituent des caractères continus qui présentaient donc un nombre potentiellement infini d’états, mais codés, pour des raisons informatiques, en un nombre limité d’états notés de 0 à 65 000. La redondance des caractères entre eux a ensuite été testée. Dans les cas où deux caractères montrent le même signal, un des deux a été supprimé. Par ailleurs, certains caractères peuvent être partiellement redondants entre eux. Pour en tenir compte, nous avons testé les effets de différents codages des caractères ASUDAS (voir encadré), produisant ainsi différentes matrices et donc, in fine différents arbres.
Les caractères ASUDAS
Parmi les caractères discrets analysés, certains concernent la morphologie dentaire. Or, la forme des dents n’est pas strictement identique entre individus d’une même espèce. Au contraire, les dents présentent des variations intraspécifiques et l’ensemble de morphologies existantes est décrite par le système ASUDAS (Arizona State University Dental Anthropology System). Ces caractères sont utilisés en médecine légale depuis de nombreuses années pour identifier le groupe ethnique d’un défunt [12]. Ils ont par ailleurs montré une capacité à reconstituer une généalogie au sein des populations de H. sapiens actuelles [13], et ont depuis été réutilisés chez des espèces fossiles pour faire de la phylogénie [14–16]. Dans notre étude, les caractères ASUDAS ont reçu une pondération deux fois plus faible que les autres parce qu’on suppose, sans pouvoir le tester, qu’ils sont partiellement redondants entre eux (notion d’intégration de caractères). En effet, les dimensions et la morphologie des différentes dents d’un individu étant fortement liées d’une dent à l’autre, il faut éviter de donner trop d’importance à des caractères qui évoluent de concert.
Résultats
À partir des différentes matrices de caractères obtenues1, un même programme informatique permet de générer l’arbre le plus parcimonieux. Pour chaque matrice, nous n’avons pas étudié que cet arbre le plus parcimonieux, mais analysé l’ensemble des 12 à 45 arbres les plus parcimonieux obtenus. L’analyse des différents arbres obtenus montre deux résultats remarquables. D’une part, ces arbres proposent une phylogénie cohérente avec celle déjà connue et obtenue à partir d’autres jeux de caractères chez les hominines (Figure 7). Cette observation est importante parce qu’elle donne du poids aux résultats de l’analyse, mais aussi parce qu’elle nous indique qu’un nombre limité de restes, quand ces derniers sont informatifs, peut permettre de faire une analyse phylogénétique de qualité.
D’autre part, les arbres obtenus fournissent des résultats homogènes qui peuvent être regroupés en deux topologies différentes : dans l’une, H. luzonensis est basal à tous les représentants du genre Homo (Figure 7A) et dans l’autre il est dérivé par rapport à H. erectus (Figure 7B).
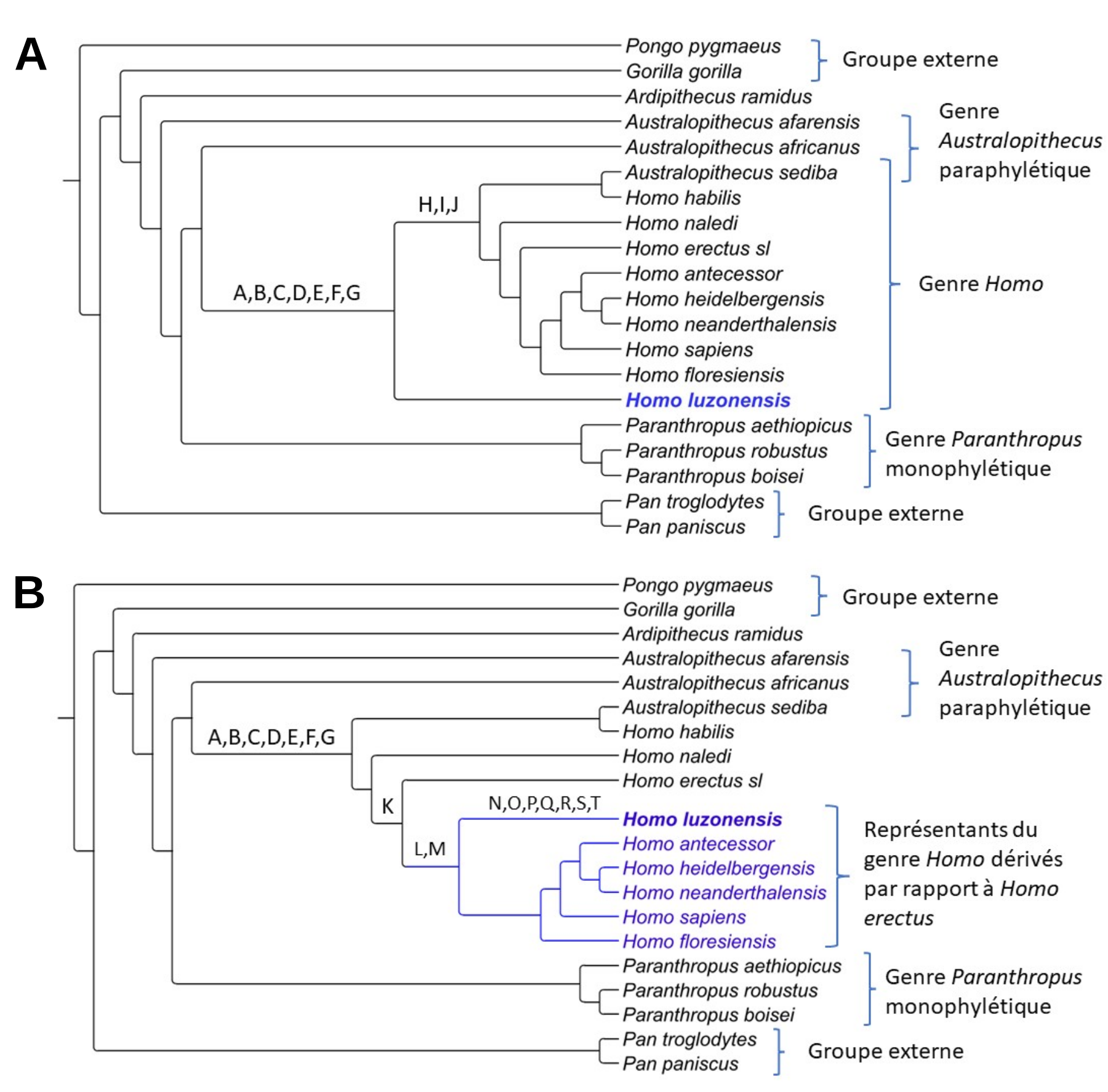
Les lettres A à G (images du haut et du bas) correspondent aux synapomorphies suivantes : bords dorsaux plats des phalanges intermédiaires de pied et du 3e métatarsien, hauteur dorso-palmaire de la base de la phalange distale de main réduite par rapport à sa longueur proximo-distale, réduction des dimensions de la cuspide1 de Carabelli sur la M3 (troisième molaire), réduction de la 5e cuspide sur la M2 et la M3 et réduction de la 6e cuspide sur la M3.
Les lettres H, I et J (image du haut) correspondent aux synapomorphies suivantes : incurvation réduite du bord dorsal des phalanges proximales de pied, « rainures buccales » réduites, et une excroissance en partie proximo-dorsale de la phalange intermédiaire de main réduite.
La lettre K (image du bas) symbolise la synapomorphie suivante : aire de la surface occlusale de la M1 plus grande que celle de la M3. Les lettres L et M (image du bas) symbolisent les synapomorphies suivantes : bord dorsal plat des phalanges intermédiaires de pied et crêtes accessoires distales absentes ou faiblement développées sur la P3 (troisième prémolaire). Enfin, les lettres N à T symbolisent les synapomorphies suivantes, qui sont aussi des réversions vers un état ancestral : fort développement des « rainures buccales » sur les prémolaires, trois racines à la P3, forte courbure de la phalange proximale de pied, zones d’insertions pour les gaines des muscles fléchisseurs fortement développées tout le long du corps de la phalange proximale de pied, faible angle d’inclinaison dorsale2 sur la phalange proximale de pied, phalange intermédiaire de main allongée et base du métatarsien basse par rapport à sa longueur. Arbre réalisé sur le logiciel Treegraph2.
Homo erectus sl désigne les Homo erectus asiatiques et africains.
1 Les cuspides correspondent aux saillies coniques de la face occlusale des dents, c’est-à-dire de la face qui triture les aliments.
2 L’inclinaison dorsale correspond à l’angle entre la face proximale et la face dorsale de la phalange.
Pour confronter les deux topologies obtenues, nous nous sommes intéressés au taux d’homoplasies (convergences ou réversions) de chaque arbre, qui permet d’évaluer la qualité du signal phylogénétique contenu en son sein [21]. Le taux d’homoplasies étant le plus faible pour l’arbre où H. luzonensis est dérivé par rapport à H. erectus (Figure 7B), c’est donc cet arbre qui rend compte de l’histoire évolutive la plus probable.
La mise en cohérence de cet arbre avec les données paléontologiques laisse supposer que H. luzonensis est probablement issu d’une population de H. erectus, qui, sous l’effet de l’évolution insulaire, a connu des réversions pour un certain nombre de caractères. H. luzonensis a cependant conservé suffisamment de caractères dérivés pour qu’on le retrouve proche des représentants du genre Homo les plus récents dans les arbres.
Importance relative des caractères dentaires et postcrâniens d’Homo luzonensis pour l’analyse phylogénétique
À l’inverse des caractères postcrâniens (ceux qui concernent les os du pied et de la main dans notre cas), les caractères dentaires sont majoritairement dérivés chez H. luzonensis : c’est-à-dire qu’ils montrent le même état chez H. luzonensis que chez les espèces récentes du genre Homo, par exemple H. sapiens, H. neanderthalensis ou H. heidelbergensis. Or, les caractères dentaires sont souvent considérés comme de bons caractères en phylogénie, dans la mesure où leur évolution se fait, pour une partie importante d’entre eux, de façon neutre, et non pas sous l’influence du milieu où évolue le taxon [17,18]. Des études récentes ont réussi, uniquement à partir des caractères dentaires, à reconstituer les mêmes relations de parenté que celles habituellement déduites à partir d’un plus grand nombre de caractères anatomiques, et même d’argumenter en faveur ou en défaveur d’hypothèses jusque-là débattues [15,16].
Pour les caractères postcrâniens, H. luzonensis présente plus souvent des états ancestraux, c’est-à-dire identiques ou proches à ceux présents chez les membres de l’extragroupe. Ces caractères ancestraux sont également retrouvés chez les australopithèques ou les paranthropes. Les caractères postcrâniens et particulièrement ceux montrant un état ancestral chez H. luzonensis ont la particularité d’être des caractères présentant une forte composante fonctionnelle. Autrement dit, il s’agit de caractères pour lesquels les études d’anatomie fonctionnelle et d’anatomie comparée permettent de dire qu’ils remplissent une fonction précise. Cependant, de telles déductions ne sont valables qu’à l’échelle d’un groupe d’espèces et ne permettent pas de tirer de conclusion pour une espèce précise. En effet, une espèce donnée peut présenter un caractère (par exemple des phalanges courbes) sans que celui-ci ne soit forcément lié à la fonction portée globalement dans le groupe d’espèces (l’escalade dans les arbres). Ainsi, la présence de phalanges courbes chez Homo luzonensis peut tout aussi bien être héritée d’ancêtres arboricoles sans que cette espèce ne le soit elle-même, qu’être un trait absent chez ses ancêtres récents mais réapparu au cours de son histoire et sélectionné car facilitant l’escalade dans les arbres. Autrement dit, comme les caractères fonctionnels peuvent aussi bien provenir d’ancêtres communs qu’être le résultat de convergences évolutives, ils peuvent compliquer les reconstitutions phylogénétiques, même si cela est discuté [19,20].
Traversée de bras de mer et endémisme insulaire
Conjointement aux passionnantes perspectives de recherche que la découverte de H. luzonensis ouvre, une grande question se pose : quand et surtout comment sont-ils arrivés sur l’île de Luçon ? Débutant il y a 2,58 millions d’années, le Quaternaire est marqué par des alternances de périodes glaciaires et interglaciaires qui ont régulièrement bouleversé non seulement le climat, mais aussi les caractéristiques géographiques des continents. En effet, les glaces qui s’accumulent en milieu continental lors des épisodes glaciaires se forment à partir de l’eau des océans et provoquent donc un abaissement du niveau marin global qui modifie, parfois de façon importante, les contours et l’étendue des terres émergées. Ainsi, l’île de Java en Indonésie, sur laquelle de très nombreux fossiles de H. erectus ont été mis au jour, a été reliée au continent asiatique à de multiples reprises au cours du Quaternaire, expliquant l’arrivée de ces hominines il y a plus de 1,6 millions d’années, à pied sec. Mais ce n’est pas le cas de l’île de Luçon où ont été découverts les restes fossiles de H. luzonensis, ni celui de l’île de Florès où H. floresiensis a été mis en évidence. Ces deux îles sont situées à l’Est de la fameuse « ligne de Wallace » (Figure 8), ligne imaginaire décrite au XIXe siècle par A.R. Wallace et distinguant les faunes et les flores présentes sur le continent asiatique de celles présentes sur le continent australien [25]. Ainsi, ces îles n’ont jamais été accessibles à pied sec et leur colonisation par n’importe quel organisme, hominines compris, implique la traversée de plusieurs bras de mer. Pourtant, des sites d’occupation humaine encore plus anciens que ceux de H. luzonensis et de H. floresiensis y ont été mis au jour [26,27]. Même si l’identité des hominines ayant occupé ces lieux reste méconnue, leur datation de 700 000 ans à 1 million d’années et les quelques dents fossiles découvertes à Florès indiquent qu’il devait s’agir de H. erectus [28]. La question des modalités et de l’intentionnalité des traversées effectuées par ces probables ancêtres de H. luzonensis et de H. floresiensis reste très débattue. S’agit-il de traversées accidentelles, de quelques individus à la dérive sur des « radeaux » naturels constitués d’amas de débris végétaux arrachés des terres lors de tsunamis, fréquents dans la région ? Ou s’agit-il de traversées intentionnelles sur des embarcations rudimentaires (dont nous ne retrouverons probablement jamais les traces, car faites de matériaux périssables) ? Cette seconde hypothèse semble difficile à envisager pour de nombreux chercheurs car jusqu’à présent, la traversée intentionnelle de bras de mer reste une innovation propre à notre espèce, H. sapiens. Mais même si nous sommes encore loin d’en comprendre tous les détails, les nombreuses découvertes effectuées en Asie du Sud-Est insulaire ces dernières années éclairent ce débat sous un nouveau jour. En effet, l’île de Célèbes (Sulawesi en indonésien) a aussi livré récemment des sites d’occupation humaine datés de 100 à 200 000 ans [29], donc bien antérieurs à la date d’arrivée de H. sapiens dans la région, estimée à environ 65 000 ans [30]. Rejeter l’hypothèse de traversées intentionnelles par toute autre espèce que H. sapiens implique donc de considérer qu’au moins trois îles de la région ont été colonisées indépendamment de façon accidentelle. Et si l’on prend en compte le fait que la colonisation de chaque île a forcément nécessité des arrivées répétées et successives de plusieurs individus jusqu’à pouvoir fonder une population viable (et dont nous avons retrouvé les traces plusieurs dizaines de milliers d’années plus tard), il n’est pas certain que l’hypothèse de traversées accidentelles soit réellement la plus parcimonieuse.
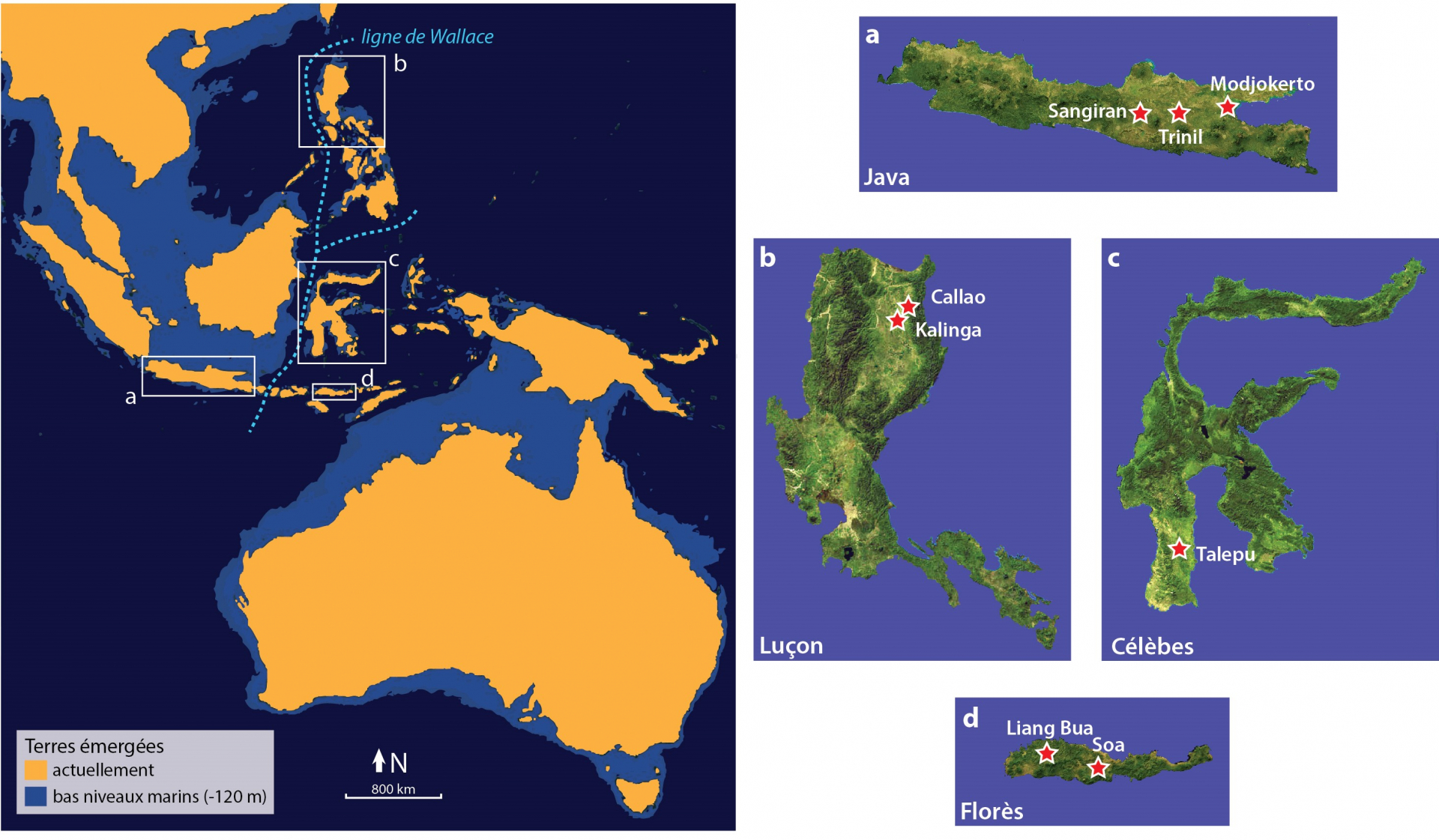
a. Java : les sites de Sangiran, Modjokerto et Trinil ont livré des fossiles d’Homo erectus datés d’environ 1,6 à 0,8 million d’années. b. Luçon : la grotte de Callao a livré les fossiles d’Homo luzonensis et le site de Kalinga a livré des outils de pierre taillée et des ossements d’animaux portant des marques de boucherie datés d’environ 700 000 ans. c. Célèbes : le site de Talepu a livré des outils de pierre taillée datés de 100 à 200 000 ans environ. d. Flores : la grotte de Liang Bua a livré les fossiles d’Homo floresiensis et les sites du bassin de la Soa ont livré des outils de pierre taillée datés d’environ un million d’années et quelques fossiles d’hominines daté d’environ 700 000 ans. (Crédits : infographie : F. Détroit ; vues détaillées des îles : NASA Visible Earth)
Il sera très difficile d’obtenir un jour la réponse définitive à cette question, mais une chose semble d’ores et déjà certaine : H. floresiensis et H. luzonensis sont de magnifiques illustrations des effets de l’endémisme insulaire. Ces effets, liés à une perte de diversité génétique des populations insulaires combinée à des ressources limitées, sont reconnus depuis très longtemps en paléontologie et ils ont été décrits sur de nombreuses espèces de vertébrés ayant évolué sur des îles [31,32]. Les effets les plus connus de l’insularité sont le nanisme, affectant les espèces de grande taille, et le gigantisme, affectant les espèces de petite taille. Le registre fossile regorge d’exemples spectaculaires, notamment dans la famille des éléphants : des espèces naines ont été découvertes sur de nombreuses îles dans le monde, notamment à Florès, mais aussi plus proche de nous puisque des éléphants mesurant à peine plus d’un mètre au garrot au stade adulte vivaient encore sur les îles de Méditerranée il y a à peine plus de 10 000 ans [33]. S’il fallait encore le démontrer, la découverte de H. luzonensis vient confirmer, après celle de H. floresiensis, que les grands principes de l’évolution biologique s’appliquent bien aux hominines, y compris lorsque ceux-ci sont soumis aux pressions de sélection très particulières des milieux insulaires.
Remerciements
Les auteurs souhaitent remercier tout particulièrement Isabelle Rouget et Jérémie Bardin (UMR 7207 CR2P, Muséum national d’Histoire naturelle), Armand S. B. Mijares (Archaeological Studies Program, University of the Philippines) et tous les collègues participants activement au Callao Cave Archaeology Project, ainsi que le National Museum of the Philippines.
Sur le même sujet
La plateforme pédagogique du Muséum national d'Histoire naturelle propose une page dédiée à Homo luzonensis dans sa formation sur l'évolution humaine.
Références
[1] D. Reich et al., « Genetic history of an archaic hominin group from Denisova Cave in Siberia », Nature, vol. 468, no 7327, p. 1053‑1060, déc. 2010, doi: 10.1038/nature09710.
[2] Q. Ji, W. Wu, Y. Ji, Q. Li, et X. Ni, « Late Middle Pleistocene Harbin cranium represents a new Homo species », The Innovation, vol. 2, no 3, p. 100132, août 2021, doi: 10/gm5hdj.
[3] X. Ni et al., « Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage », The Innovation, vol. 2, no 3, p. 100130, août 2021, doi: 10/gnh6d4.
[4] P. Brown, T. Sutikna, M. J. Morwood, et R. P. Soejono, « A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia », Nature, vol. 431, no 7012, p. 1055‑1061, 2004.
[5] F. Détroit et al., « A new species of Homo from the Late Pleistocene of the Philippines », Nature, vol. 568, no 7751, p. 181‑186, avr. 2019, doi: 10/gfx84m.
[6] P. Gousset, « Analyse phylogénétique d’Homo luzonensis : taxon, caractères, phylogénie et évolution insulaire », Mémoire de Master 2, Muséum National d’Histoire Naturelle, Paris, 2021.
[7] A. S. Mijares et al., « New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines », J. Hum. Evol., vol. 59, no 1, p. 123‑132, juill. 2010, doi: 10.1016/j.jhevol.2010.04.008.
[8] C. Zanolli et al., « Further analyses of the structural organization of Homo luzonensis teeth: Evolutionary implications », J. Hum. Evol., vol. 163, p. 103124, févr. 2022, doi: 10.1016/j.jhevol.2021.103124.
[9] J. T. J. Stern et R. L. Susman, « The locomotor anatomy of Australopithecus afarensis », Am. J. Phys. Anthropol., vol. 60, no 3, p. 279‑317, 1983, doi: 10/dm9xtr.
[10] B. Latimer, J. C. Ohman, et C. O. Lovejoy, « Talocrural joint in African hominoids: Implications for Australopithecus afarensis », Am. J. Phys. Anthropol., vol. 74, no 2, p. 155‑175, 1987, doi: 10.1002/ajpa.1330740204.
[11] D. Strait, F. E. Grine, et J. G. Fleagle, « Analyzing Hominin Hominin Phylogeny: Cladistic Approach », in Handbook of Paleoanthropology, W. Henke et I. Tattersall, Éd. Berlin, Heidelberg: Springer Berlin Heidelberg, 2015, p. 1989‑2014. doi: 10.1007/978-3-642-39979-4_58.
[12] G. R. Scott, C. G. Turner II, G. C. Townsend, et M. Martinón-Torres, The Anthropology of Modern Human Teeth: Dental Morphology and Its Variation in Recent and Fossil Homo sapiens, 2e éd. Cambridge: Cambridge University Press, 2018.
[13] G. R. Scott et J. D. Irish, Human Tooth Crown and Root Morphology. Cambridge University Press, 2017.
[14] M. Martinón-Torres et al., « Dental evidence on the hominin dispersals during the Pleistocene », Proc. Natl. Acad. Sci., vol. 104, no 33, p. 13279‑13282, 2007.
[15] J. D. Irish, D. Guatelli-Steinberg, S. S. Legge, D. J. de Ruiter, et L. R. Berger, « Dental Morphology and the Phylogenetic “Place” of Australopithecus sediba », Science, vol. 340, no 6129, p. 1233062‑1233062, avr. 2013, doi: 10.1126/science.1233062.
[16] J. D. Irish, S. E. Bailey, D. Guatelli-Steinberg, L. K. Delezene, et L. R. Berger, « Ancient teeth, phenetic affinities, and African hominins: Another look at where Homo naledi fits in », J. Hum. Evol., vol. 122, p. 108‑123, sept. 2018, doi: 10/gdxrhk.
[17] A. Gómez-Robles et P. D. Polly, « Morphological integration in the hominin dentition: evolutionary, developmental, and functional factors », Evolution, vol. 66, no 4, p. 1024‑1043, avr. 2012, doi: 10.1111/j.1558-5646.2011.01508.x.
[18] H. Rathmann, H. Reyes-Centeno, S. Ghirotto, N. Creanza, T. Hanihara, et K. Harvati, « Reconstructing human population history from dental phenotypes », Sci. Rep., vol. 7, no 1, p. 12495, déc. 2017, doi: 10/gb23b3.
[19] F. S. Szalay, « Functional analysis and the practice of the phylogenetic method as reflected by some mammalian studies », Am. Zool., vol. 21, no 1, p. 37‑45, 1981, doi: 10/csg42f.
[20] I. Tattersall et N. Eldredge, « Fact, theory, and fantasy in human paleontology », Am. Sci., vol. 65, no 2, p. 204‑211, avr. 1977.
[21] P. Darlu, P. Tassy, C. d’Haese, et R. Zaragüeta i Bagils, La reconstruction phylogénétique. Editions Matériologiques, 2019.
[22] P. A. Goloboff, « Estimating character weights during tree search », Cladistics, vol. 9, no 1, p. 83‑91, 1993, doi: 10/ffq8j2.
[23] P. Y. Sondaar, « Paleoecology and evolutionary patterns in horses and Island mammals », Hist. Biol., vol. 8, no 1‑4, p. 1‑13, oct. 1994, doi: 10/bw8jnb.
[24] L. W. van den Hoek Ostende, « Cladistics and insular evolution, an unfortunate marriage? Another tangle in the Deinogalerix analysis of Borrani et al. (2017) », Cladistics, vol. 34, no 6, p. 708‑713, déc. 2018, doi: 10/gdwrzz.
[25] A. R. Wallace, The Malay Archipelago, vol. 1989. (1989) Singapore, Oxford, New York: Oxford University Press, 1869.
[26] T. Ingicco et al., « Earliest known hominin activity in the Philippines by 709 thousand years ago », Nature, vol. 557, no 7704, p. 233‑237, mai 2018, doi: 10/cn87.
[27] A. Brumm et al., « Early stone technology on Flores and its implications for Homo floresiensis », Nature, vol. 441, no 7093, p. 624‑628, juin 2006, doi: 10.1038/nature04618.
[28] G. D. van den Bergh et al., « Homo floresiensis-like fossils from the early Middle Pleistocene of Flores », Nature, vol. 534, no 7606, p. 245‑248, juin 2016, doi: 10.1038/nature17999.
[29] G. D. van den Bergh et al., « Earliest hominin occupation of Sulawesi, Indonesia », Nature, vol. 529, no 7585, p. 208‑211, janv. 2016, doi: 10/bbwg.
[30] C. Clarkson et al., « Human occupation of northern Australia by 65,000 years ago », Nature, vol. 547, no 7663, p. 306‑310, juill. 2017, doi: 10.1038/nature22968.
[31] A. A. E. van der Geer, M. V. Lomolino, et G. A. Lyras, « ‘Island Life’ before man: biogeography of palaeo-insular mammals », J. Biogeogr., vol. 44, no 5, p. 995‑1006, 2017, doi: 10/f95znh.
[32] J. Vos, L. Ostende, et G. Bergh, « Patterns in insular evolution of mammals: a key to island palaeogeography », in Biogeography, time, and place: distributions, barriers, and islands, 2007, p. 315‑345.
[33] A. Athanassiou, A. A. van der Geer, et G. A. Lyras, « Pleistocene insular Proboscidea of the Eastern Mediterranean: a review and update », Quat. Sci. Rev., vol. 218, p. 306‑321, 2019, doi: 10/gf43nv.
[34] H. K. Voris, « Maps of Pleistocene sea levels in Southeast Asia: shorelines, river systems and time durations », J. Biogeogr., vol. 27, no 5, p. 1153‑1167, 2000.


