Cet article présente deux mécanismes faisant évoluer les fréquences alléliques d'une population au cours du temps : la sélection naturelle et la dérive génétique.
Dérive et sélection
Définitions
Si la sélection s'exerce sur le phénotype, au niveau individuel, la compréhension de la variation des caractères au cours du temps et donc des processus évolutifs demande que l'on s'intéresse à une échelle plus large : des ensembles d'individus (populations).
Population : ensemble d’individus dont tous ont la même probabilité de se croiser entre eux, et qui se reproduisent moins (ou pas du tout) avec les membres d’autres populations. Mais cette reproduction diminuée est due à des critères extrinsèques (spatiaux par exemple), et non à des barrières génétiques entre les groupes : les populations ne sont pas des espèces. Cependant, il peut y avoir une différenciation génétique entre populations, avec des fréquences alléliques, voire des allèles, spécifiques pour chacune (comme par exemple dans le cas des groupes sanguins dans les populations humaines).
La diversité génétique résultant des mutations et remaniements du génome fait qu'à un instant donné, une population naturelle n'est pas homogène d'un point de vue génétique : il existe pour la plupart des séquences, qu'elles soient codantes ou non, plusieurs allèles, dont les fréquences (fréquences alléliques) vont varier au cours du temps. Les généticiens des populations se sont attachés à modéliser ces variations et les paramètres qui les influencent. Un des sujets d'intérêt particulier est l'étude de la manière dont une mutation sur une séquence (donc présente chez un seul individu, sur une seule de ses deux copies du même locus), peut se répandre dans une population, voire un ensemble de populations.
La dérive génétique
Dérive génétique : processus par lequel les fréquences alléliques changent dans les populations à cause de biais aléatoires d’échantillonnage dans la transmission des allèles d'une génération à l'autre.
Si les premiers travaux sur le sujet, à commencer par ceux de Darwin, proposaient que le tri entre les variants soit l'œuvre des sélections naturelle et sexuelle, il a depuis été établi que le hasard joue également un rôle très important dans l'évolution des fréquences alléliques. En 1968, Motoo Kimura introduisit la théorie neutraliste de l’évolution : de nombreux allèles ne sont ni avantageux, ni désavantageux, et donc ne sont pas ou peu soumis à la sélection (allèles neutres : par exemple les allèles résultant de mutation synonymes). Leur devenir ne dépend que du taux de mutation et de ce qui a été appelé la dérive génétique.
Lorsque l'on connaît les allèles portés par les parents, on peut théoriquement évaluer la probabilité de présence de ces allèles dans la descendance (génétique mendélienne). Cependant, la distribution véritable des allèles ne sera pas rigoureusement déterminée, mais fluctuera de manière stochastique, en particulier sur de petites populations, même pour des systèmes simples où n'interviennent qu'un gène et deux allèles. En effet, la descendance réelle est un échantillonnage aléatoire d'un nombre limité de tirages au hasard. Cela signifie que la distribution de deux allèles dans les descendants peut être différente de celle dans la génération parentale.
Un exemple de dérive
Prenons un croisement entre deux souris, une blanche (homozygote g//g) et une grise (hétérozygote G//g), l'allèle G étant dominant et produisant des souris grises. On peut faire une série de tests en employant des haricots de deux couleurs (ou deux types de pièces) en proportions égales, et en faisant des séries de six (par exemple) tirages aléatoires de haricots pour simuler les allèles transmis par la souris grise à six descendants (puisque l’allèle transmis par la blanche sera forcément g). Si, sur un grand nombre de tirages, la moyenne sera de ½ de descendants gris, et ½ de blancs, sur chaque série de six tirages prise indépendamment le nombre de chaque type de descendants pourra varier de 0 à 6 (Figure 1).
À chaque génération, si les descendants sont croisés entre eux (ce qui est le cas dans une population), ce processus se répète, et au bout d'un nombre variable de générations, l'un des deux allèles sera complètement et définitivement éliminé, et tous les individus seront porteurs de l'autre (voir figures 1 et 2 ). À ce moment, le seul allèle restant est dit fixé, et comme c'est le seul présent, tous les descendants en seront porteurs. Mais tôt ou tard de nouvelles mutations se produisent, créant ainsi de nouveaux allèles, et le processus se répète. La probabilité de fixation est (en moyenne) fonction de la proportion de chacun des allèles dans la population de départ. Un allèle présent en un seul exemplaire a donc une probabilité de fixation qui est fonction de la taille de la population : si la population est de deux individus diploïdes, la proportion représentée par l’allèle est de 1/4. Si par contre la population est de mille individus diploïdes, il n’est présent que dans une proportion de 1/2000…
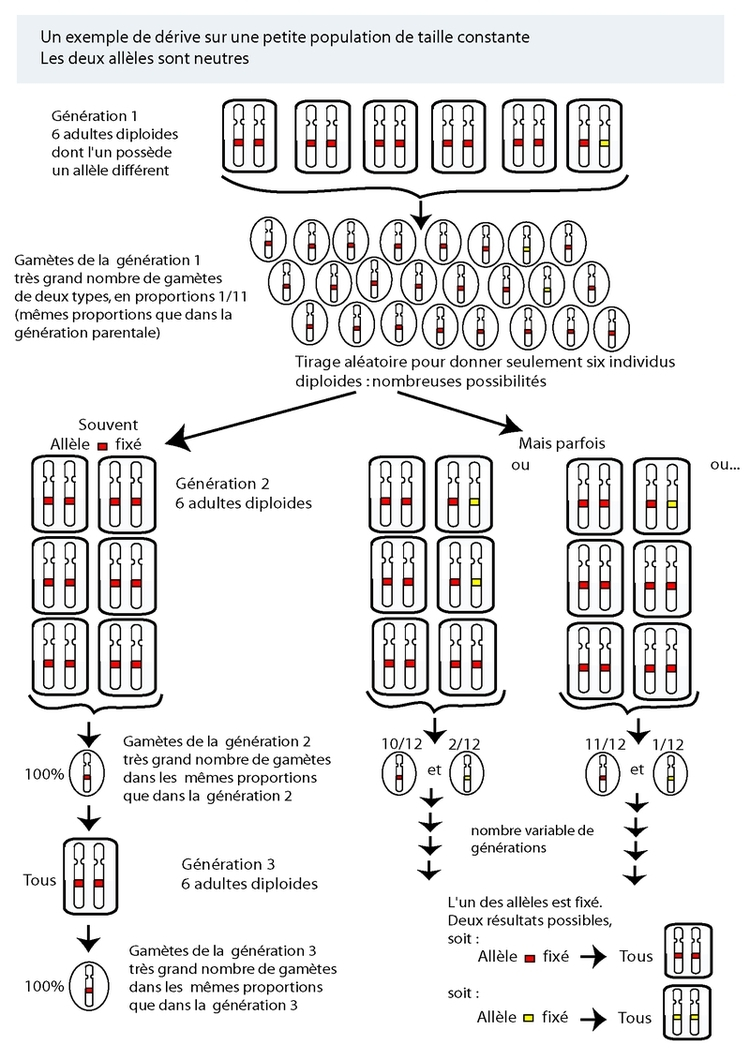
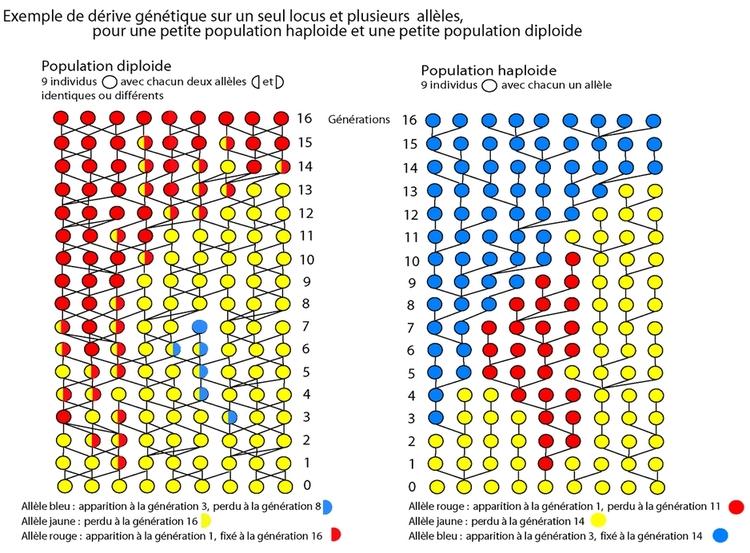
Le temps mis par un nouvel allèle pour envahir une population est fonction de la taille de la population de gènes (n : le nombre d’exemplaires de ce locus existant dans la population d'organismes considérés), et est en moyenne de 2n générations. Ainsi, plus la taille de la population est réduite, plus un allèle peut se fixer rapidement. La plupart des nouveaux allèles seront éliminés par ce processus.
Une population (théorique) avec un nombre infini de reproducteurs n’est pas soumise à la dérive (un nombre infini de tirages donnera la valeur moyenne), tandis qu’une population avec un nombre très réduit de reproducteurs sera soumise à une dérive forte (nombre limité de tirages : biais d’échantillonnage fort). Étant donné que le processus de création de variabilité, et celui de sa fixation (ou, plus souvent, de son élimination) sont aléatoires, si des groupes (de taille finie) au départ identiques d'individus sont isolés, et maintenus en isolement pendant un temps suffisant, des différences vont se créer et s'accumuler au fil du temps entre les deux groupes. Si aucun processus d'homogénéisation entre les deux groupes n'est présent (échange de migrants pour assurer des échanges d'allèles), après un certain temps les différences seront suffisantes pour que les deux groupes ne soient plus compatibles et interféconds (spéciation).
Cette explication à l'aide de deux allèles est bien sûr schématique. De nouveaux allèles au même locus peuvent apparaître avant que ceux qui étaient précédemment présents dans la population ne soient fixés ou perdus. Ainsi, à un instant donné, dans une population, il existe souvent plusieurs allèles pour un même locus (exemples des groupes sanguins et HLA). Les fréquences observées aujourd'hui dans les diverses populations humaines correspondent à un instantané de ces fréquences à un instant donné : elles ont varié dans le temps, divergeant entre les groupes ou se réhomogénéisant lors d’échanges de migrants, et continuent à varier encore...
La sélection
Qu’est-ce que la sélection ?
La sélection peut agir à tout moment de la vie de l’individu : âge adulte, juvénile, mais également embryon, voire gamètes. La sélection intervient en fonction de la valeur sélective des allèles présents dans la population les uns par rapport aux autres, pas à une valeur absolue. Il existe plusieurs types de sélection : sélection naturelle et, pour certaines espèces, sélection sexuelle.
Sélection naturelle : survie et/ou reproduction différentielle d’organismes qui se distinguent les uns des autres par au moins un caractère héritable. Cette survie ou reproduction différentielle peut être due à l’influence du milieu, à de la compétition à l’intérieur de la population, ou à une compétition avec d’autres espèces (prédation, compétition pour la nourriture…).
Sélection sexuelle : reproduction différentielle due à un comportement reproducteur, soit par une compétition entre les membres d’un même sexe pour l’accès à l’autre sexe (combats, intimidation…), soit (ou et) par sélection par l’un des sexes de ses partenaires à partir de son aspect physique ou d’un comportement particulier.
Adaptation : Une adaptation est un changement qui offre un avantage pour la survie et/ou la reproduction d’un individu. L’origine d’une adaptation est une mutation/modification du génome due au hasard, mais sa dispersion dans la population est partiellement déterminée par sa valeur adaptative par rapport à la forme d’origine (et partiellement stochastique).
Sélection et dérive
La sélection n’a pas un effet immédiat, qui remplacerait automatiquement une version (un allèle) par une autre plus avantageuse, ou éliminerait des allèles à effet délétère. Elle introduit plutôt un paramètre supplémentaire pour la probabilité de fixation des allèles les uns par rapport aux autres (Figure 3). Ce paramètre rend compte du fait que les porteurs de l’un des deux (ou plus) allèles en compétition auront une meilleure probabilité de survie et/ou de meilleurs capacités reproductives, et laisseront donc en moyenne plus de descendants. Plus la différence entre la valeur sélective des deux allèles en compétition est grande, plus l’effet sera fort (Figure 4).
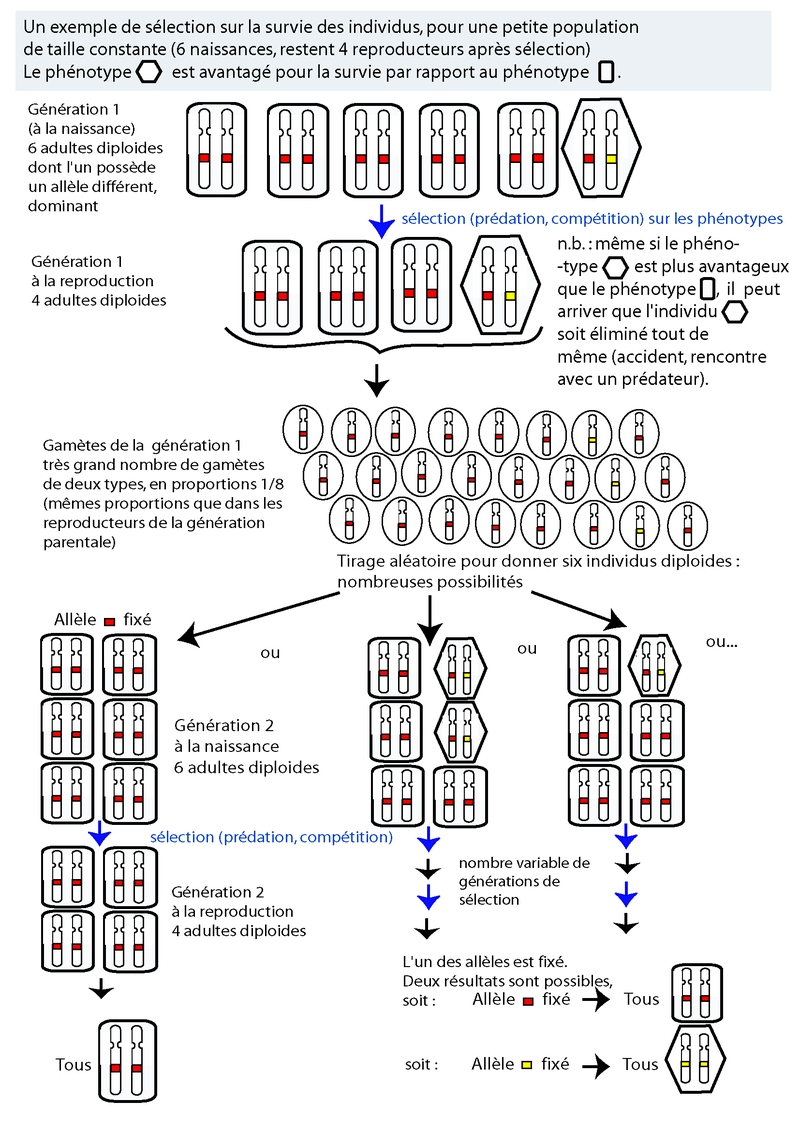
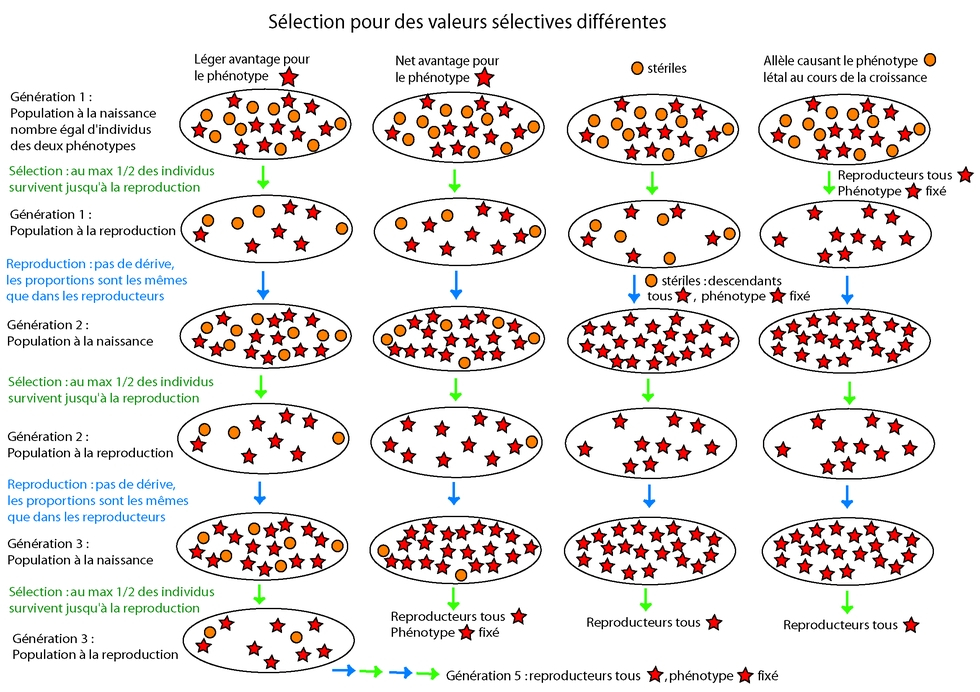
Fixation d’un allèle : Un allèle est dit fixé dans une population lorsque seul cet allèle est encore présent dans la population, les autres ayant disparu.
Pour la discussion sur la dérive, les deux allèles étaient considérés comme étant de valeur équivalente pour les individus. Mais bien que cela soit souvent le cas, il arrive également que le nouvel allèle donne un produit moins efficace (ou, plus rarement, plus efficace) que l'allèle dont il est issu. La probabilité de fixation d'un allèle avantageux est augmentée par rapport à celle de l'allèle avec lequel il est en « compétition ». Ce qui est important, c'est non seulement le taux de survie, mais également le nombre de descendants laissé. Les chercheurs en génétique des populations ont introduit pour cela la notion de fitness des individus. Ce paramètre, également appelé valeur sélective, prend en compte non seulement la probabilité de survie des individus, mais également leur capacité de reproduction, c'est-à-dire de transmission de l'allèle d'intérêt à la génération suivante. Un variant a beau être extrêmement avantageux, s'il ne transmet pas la variation à la génération suivante, il n'aura pas d'impact sur l'évolution à long terme de sa population ou de son espèce.
Des allèles produisant un effet négatif important seront généralement éliminés, mais pas toujours aussi vite qu'on pourrait l'imaginer au premier abord. Par exemple, un allèle même létal, à partir du moment où il est récessif, peut rester présent un temps dans une population (mais pas se fixer, puisqu'il est létal à l'état homozygote). De même, un allèle dominant, mais dont l'effet négatif se fait sentir après que les individus se sont reproduits, peut être maintenu (les allèles à l'origine de la chorée de Huntington en constituent des exemples). D’autre part, il est important de se souvenir ici des mécanismes de crossing-over, dont la fréquence est dépendante de la distance entre les loci. Ainsi, si un allèle à un locus subit une sélection positive, les allèles d’autres gènes situés à faible distance de celui-ci peuvent être « entraînés » par cette sélection et fixés également, même s’ils sont neutres ou légèrement délétères.
Les importances relatives de la sélection, de la dérive et de la vitesse d'apparition des mutations dans le processus évolutif sont encore discutées. Il faut également garder à l’esprit qu’un individu est une combinaison (souvent unique) de très nombreux allèles, et que les effets de divers allèles sont parfois difficilement séparables. Il est donc tout à fait abusif de dire qu’on peut prévoir l’évolution d’une population naturelle dans son milieu naturel (certes on peut prévoir l’avenir d’une population si l’on éradique son milieu !). D’autant plus abusif que la sélection qui s’opère sur ces nombreux allèles est elle-même le fruit de nombreux événements contingents ayant lieu dans le milieu biotique et abiotique. Aucune évolution biologique n’est donc prévisible.
Hasard et déterminisme
Les modifications au niveau du génome se font de manière aléatoire, et non en fonction de l'avantage ou du désavantage que cela procurera à l'organisme. Cependant, il est possible d'évaluer approximativement la vitesse de modification d'une séquence, voire d'un génome1, mais non les sites des prochaines mutations. Cependant, la production de variabilité opère en général par modification de séquences/structures existantes : c'est le « le bricolage » décrit par François Jacob dans Le jeu des possibles. Le bricolage est ce qui reste de contrainte due au fonctionnel et au préexistant, tandis que les essais et erreurs sont eux aléatoires. À un moment donné, dans un individu donné, tout n’est donc pas possible. Ce concept de réutilisation et modification de structures produisant de la nouveauté fonctionne à l’échelle des organes comme à celle des gènes.
Les fréquences alléliques fluctuent de manière stochastique (bien que répondant à une loi de probabilité, dans laquelle interviennent la fréquence des allèles et leur valeur sélective) : il n'est cependant pas possible de prédire si un allèle donné va se fixer ou pas, sauf dans le cas de valeurs sélectives extrêmes. La sélection ramène donc une certaine directionnalité si les conditions des milieux biotiques et abiotiques sont suffisamment constantes. Mais cette directionnalité même si elle se montre parfois très forte, n’est pas absolue. Il reste une composante stochastique.
Processus stochastique : processus au comportement non déterministe : une étape est partiellement (mais pas complètement) déterminée par l’étape précédente.
Les environnements génétiques nouveaux créés par des associations d'allèles originaux sont encore souvent imprévisibles pour les chercheurs, du fait de leur complexité. Un environnement génétique particulier va être déterministe, mais nous ne maîtrisons pas suffisamment les éléments pour être capables de formuler des prédictions. Gardons aussi à l’esprit que ce déterminisme a lui-même ses limites : dans le dernier quart du XXe siècle est apparue l’idée que les gènes ne contrôlent pas tout ; et qu’une partie de la variabilité phénotypique est épigénétique.
À une plus grande échelle, celle de l'histoire des organismes, si la théorie générale de l'évolution explique partiellement le cours effectif des événements, cette explication est loin d'être complète. Dans ce cas, il ne s'agit pas seulement de notre incapacité à retrouver/modéliser des interactions et des circonstances complexes, mais également d'une composante contingente. L'histoire des organismes est un processus historique, dont le déroulement a souvent dépendu de la présence d'une population avec une composition génétique donnée, dans un endroit donné et des conditions données (dépendance sur les conditions initiales, et pas seulement les lois). Si n'importe laquelle de ces conditions initiales est différente (or plusieurs dépendent de processus aléatoires ou stochastiques pour leur mise en place), l'issue sera radicalement différente, et influera sur les populations et les circonstances suivantes, de sorte que toute l'histoire sera modifiée. Ce type d'effets est bien connu en histoire : c’est la notion de « processus historique », où chaque étape improbable conditionne la survenue d’étapes suivantes a priori improbables. Ainsi le cours de nombreux événements de grande importance apparaît a posteriori avoir été influencé par les actions de peu d'individus, des problèmes matériels, ou bien des circonstances météorologiques, et où toute modification mineure aurait eut des résultats considérablement différents. Il existe d'ailleurs chez les historiens une tradition d'essais sur « ce qui aurait pu se produire si... », et la majorité s'accorde à dire que si l'histoire de l'humanité était rejouée, il est fort probable que les événements se produirait de manière différentes, puisque de petites modifications peuvent produire de très grands changements. Cependant, ce type de récits ne fonctionne bien que sur la description d'un intervalle de temps restreint après le changement dans l'histoire : s'il est possible de d'extrapoler sur ce qui se serait, par exemple, produit immédiatement après l'invasion de l'Angleterre si l'Armada espagnole avait réussi en juillet-août 1588, les cascades d'effets inconnus, et l'accumulation d'autres événements contingents rendent les prédictions à plus de quelques mois du changement complètement impossible.
Notons qu’une succession d’événements qui s’enchaînent causalement, tous aussi improbables soient-ils, conduit toujours à un résultat. L’un des défauts de la rétrospective humaine est d’oublier la notion de processus historique en inversant les causes et les effets. Constatant que, si à l’origine les conditions avaient été très légèrement différentes, la face du monde en eût été bouleversée, on en conclut (à tort) que ces conditions furent réglées pour que le monde fût tel que nous le voyons. Sur le plan épistémologique, c’est une erreur de raisonnement : le monde présent ne saurait imposer logiquement au monde passé le recours à une programmation. De plus, nous n’avons aucune explication rationnelle pour interpréter comment un passé pourrait être programmé à devenir le présent. Sauf en sortant du champ scientifique pour évoquer des entités surnaturelles ou des interventions providentielles. C’est une posture répandue dont Voltaire déjà s’était moqué, présente non seulement dans le public, mais aussi chez les astronomes, sous le nom de « principe anthropique », et chez les paléoanthropologues teilhardiens, entre autres.
Pour plus d'éléments sur ces questions, le texte de J. Gayon Hasard et évolution (L'évolution, Le Guyader, H.), et le livre de S. J. Gould La vie est belle, sont de bons points de départ. La revue Ciel et Espace traite du principe anthropique dans un numéro Hors Série d’octobre 2006 intitulé L’univers a-t-il besoin de Dieu ?
Génotype et phénotype
Il reste encore beaucoup à découvrir au niveau des liens entre le phénotype et le génotype. Or, la sélection se fait au niveau du phénotype, mais la création de variabilité transmissible et sa fixation se font au niveau du génotype. Comment s’opèrent les changements phénotypiques de plus large ampleur que des variations sur un seul allèle, comme la couleur des yeux, ou le mélanisme ? Comment ont pu se mettre en place à partir d’un ancêtre commun des organismes différent autant par leur organisation que ceux observés dés le Cambrien ?
Plan d’organisation : organisation d’un être virtuel, idéal, supposé représenter ce que l’on trouve généralement dans un groupe d’organismes. Il y a le plan d’organisation des arthropodes, celui des chordés… Le rang taxonomique « phylum » est défini comme l’ensemble des organismes partageant un plan d’organisation. Il s'agit d'une notion uniquement pratique (une division artificielle) destinée à faciliter l’apprentissage de la zoologie et de la botanique, qui n'est pas compatible avec une compréhension évolutionniste du vivant. En effet, les êtres vivants sont conçus par les évolutionnistes comme des mosaïques de traits primitifs ou dérivés, et dont les modifications ne se produisent pas toutes en même temps. En figeant la nature dans une conception idéaliste, en utilisant la métaphore ambiguë du « plan », le plan d’organisation créé davantage de blocages épistémologiques qu’il n’aide à la pédagogie.
Une complexité irréductible ?
L’un des arguments favoris de ceux qui nient l’existence de l’évolution est que certains organes, ou certaines cascades enzymatiques, sont bien trop complexes pour pouvoir être apparus simplement « par hasard » (ce qui n'est pas ce que dit la théorie générale de l'évolution d'ailleurs). À quoi peut servir un œil partiel ?
Il y a de fait deux niveaux de malentendu différents dans ce type de question.
- L'un est en relation avec la notion de contingence et de hasard : l'improbabilité d'apparition d'une telle structure, qui contient lui même deux problèmes différents.
L'un d'entre eux est que nous raisonnons à postériori : nous observons un résultat final, sans avoir accès aux circonstances intermédiaires, ni à des informations sur le résultat final si nous refaisions le déroulement de l'histoire de la vie d'un bout à l'autre. Si on tire dans un jeu de 52 cartes dix cartes au hasard, la probabilité de l'agencement particulier de cartes obtenu est extrêmement faible. Et pourtant, c'est bien celui qui a été tiré. Si notre univers n'est qu'un seul parmi des millions de possibles, la probabilité initiale que les constantes physiques soient juste comme il faut pour permettre le développement de la vie était extrêmement faible. Mais, même si c'est vrai, le tirage a déjà été fait, et si son résultat avait été autre, nous ne serions tout simplement pas là à nous poser la question... La probabilité d'un événement passé est de 1 : il est présent, et il ne changera pas, quel qu'ait été la faiblesse de la probabilité de chacune des étapes ayant conduit à celui ci.
D'autre part, il y a l'idée d'influence de chaque étape sur l'étape suivante : il s'agit d'un processus historique, comme discuté dans la partie précédente. Pour l'évolution de l'oeil comme pour de nombreux systèmes enzymatiques intégrés, d'ailleurs, il est particulièrement intéressant de comparer divers organismes, qui sont arrivés à des résultats d'efficacité comparables en utilisant des éléments différents. Cependant, une fois qu'une voie est empruntée, les itérations d'essai et erreur se font à partir de celle ci, pour conduire à des résultats très différents.
De nouveau, pour employer l'analogie avec le paquet de cartes, si nous faisons un premier tirage de vingt cartes, dans lequel nous tirerons ensuite les dix cartes, les probabilités de tirage de la main particulière ont changé. Les circonstances initiales du second tirage sont différentes, et certains agencements sont devenus totalement impossibles, du fait de l'absence de certaines cartes dans le lot dans lequel sera fait le tirage... (voir également J. S. Pierre p. 624 dans Les matérialismes (et leurs détracteurs), Syllepse, 2004). - D'autre part, pour en revenir à la compréhension des étapes intermédiaires possibles de l'évolution de l'œil ou de cascades protéiques, il n’est en effet pas possible de répondre à ces questions si on ne s’intéresse qu’à l’humain, ou qu’aux mammifères. Par contre, l’étude comparée de groupes beaucoup plus éloignés ainsi que l'étude du développement ont donné lieu à des centaines de publications et surtout ont fourni des éléments de réponse convaincants sur un certain nombre d’exemples, même si la combinaison de ces deux champs d’étude n’a réellement pris son essor qu’au cours des vingt dernières années.
Enfin, un aspect particulièrement intéressant dans l'évolution de structures complexes est l'existence de « recyclage » de structures pour de nouvelles fonctions, autre que celles pour lesquelles elles ont été sélectionnées à l'origine (exaptation). Il en existe plusieurs exemples très célèbres, comme les plumes, dont le rôle à l'origine n'était pas du tout l'aide au vol, ou la langue, qui n'a pas été sélectionnée pour permettre la parole, mais de très nombreux exemples existent au niveau moléculaire.


Évolution, développement, et régulation de l’expression
Pendant presque tout le XXe siècle, l’étude du développement et celle de l’évolution ont été découplés. Mais eu cours des vingt dernières années, d’importantes découvertes combinant le développement et le moléculaire, puis la comparaison entre différents organismes servant de modèle, sont venus bouleverser cet état de choses. Aujourd'hui l'évo-dévo (évolution-développement) est l'une des approches les plus fructueuses en ce qui concerne la compréhension des liens entre le génotype et le phénotype. Pour comprendre des différences morphologiques, les chercheurs s’attachent à décrire et comprendre les modalités de la mise en place du phénotype au cours du développement, et le pourquoi et comment des différences entre ces modalités pour différents organismes.
Il existe de telles différences morphologiques et développementales entre les divers phylums eucaryotes que de nombreux chercheurs avaient manifesté un certain scepticisme quand à ce que l'étude du développement de modèles animaux aussi éloignés que la drosophile pourrait bien apporter à notre compréhension du développement des vertébrés, et plus particulièrement des mammifères. L'accumulation de divers mutants plus ou moins étranges pouvait sembler n'avoir que peu de portée pour éclaircir les zones sombres de notre compréhension de l'évolution.
La découverte de gènes homologues extrêmement conservés entre la souris et la drosophile, et qui semblaient impliqués à un niveau fondamental dans le développement vint donc comme une surprise. Plus surprenant encore, certains gènes homologues sont impliqués dans des fonctions clairement équivalentes chez la drosophile et les mammifères. Par exemple, le gène eyeless de la drosophile dont l'inactivation produit une perte des yeux, est l'homologue du gène Small eye de la souris, où les mutants présentent une perte ou une réduction des yeux (ces deux gènes sont également connus sous le nom de Pax-6)... La conservation est telle qu'il est possible, en introduisant dans des larves de drosophile le gène qui déclenche la formation d'yeux chez la souris, de déclencher la formation à cet endroit d'un oeil sur le corps de l'insecte. Mais l'oeil ainsi formé est un oeil de drosophile, pas de souris : le produit du gène ne donne pas les instructions pour la construction de l'oeil, il déclenche une cascade d'expressions de gènes variés qui vont avoir cet effet. Pax-6 n'est pas le seul gène à avoir des équivalents impliqués dans des fonctions comparables dans des groupes éloignés : par exemple, les gènes Distal-less, impliqué dans la formation d'appendices, et tinman, impliqué dans la formation du coeur chez la drosophile, ont également leurs homologues et équivalents chez les vertébrés (et même les échinodermes et les vers marins pour Distal-less). Il apparaît donc que les gènes présents chez l'ancêtre commun de ces groupes ont été non seulement conservés en séquence, mais également au niveau de certaines de leurs fonctions de pilotage d’expression de gènes pour former les structures, tandis que les organes assurant ladite fonction ont évolué jusqu'à en devenir très différents.
Tous ces gènes appartiennent à des familles différentes, mais partagent la possession d'homéoboîtes capables de se fixer à des séquences spécifiques de l'ADN génomique, et d'en contrôler l'expression. Ils sont connus sous le nom de gènes homéotiques, ou parfois gènes architectes, car toute inactivation (intégrale ou partielle) de l'un de ces gènes causera des défauts (ou du moins des modifications) plus ou moins radicales du développement.
Ce qui est encore plus intéressant pour la compréhension des différences majeures entre les organismes, c'est que l'expression de ces gènes est clairement délimitée dans le temps et dans l'espace au cours du développement des individus. Souvent, des structures particulières (appendices, limites de segment, organes...) coïncident avec les zones d'expressions de l'un de ces gènes, avec les zones où cesse l'expression de l'un de ces gènes, ou avec les zones dans lesquelles sont combinées les expressions de plusieurs de ces gènes. La modulation de la position où un tel gène est exprimé, ou du moment du développement auquel il intervient, peuvent permettre de « construire » des organismes très différents à partir du même set d'instructions, en utilisant parfois un même gène homéotique à plusieurs reprises. En effet, les changements et modulations interviennent plutôt par des modifications dans les régions régulatrices que sur la séquence même du gène, ce qui permet une régulation très fine. Les sites de régulation de ces gènes vont dicter où et quand au cours du développement ils seront exprimés et déclencheront l’activation de la transcription d’une cascade d’autres gènes. Les régions régulatrices peuvent être multiples pour un seul gène, et la transformation d’un site de régulation permet des modifications sur l’un des épisodes d’expression du gène (décalage dans le temps ou l’espace) sans toucher à ses autres épisodes d’expression.
Des décalages dans les zones d'expressions de divers gènes homéotiques permettent ainsi par exemple des changement dans le nombre de segments d'un type particulier (et donc du nombre et de la distribution des divers types d'appendices sur l'organisme). Voir fig. 3.
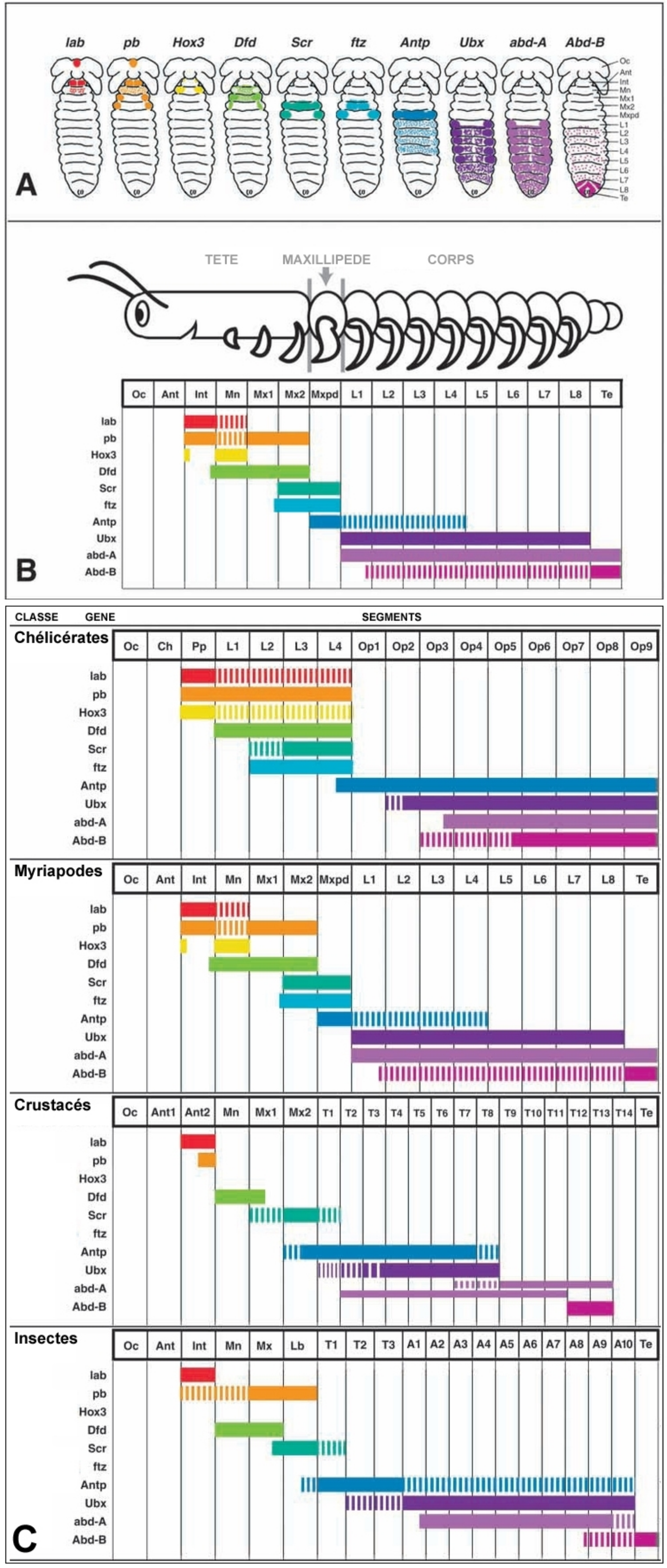
(A) et (B) Patrons d'expression de dix gènes Hox chez les Centipèdes (Arthropodes Myriapodes) présenté sur un schéma d'embryon (A) et sous forme de diagramme (B). (C) Patron d'expression de dix gènes Hox chez différents sous-embranchements d'Arthropodes. Les différentes espèces d'Arthropodes ont souvent un nombre différent de segments. Les diagrammes présentés sont basés respectivement sur les araignées Cupiennius et Achaearanea (Chélicérates), le centipède Lithobius à l'éclosion (Myriapode), le cloporte Porcellio (Crustacé) et le lepisme Thermobia (Insecte).
Les barres pleines indiquent une forte expression, les barres tiretées une expression faible ou transitoire. Chaque barre résume les informations d'expression spatiale et temporelle de plusieurs espèces et constitue donc une version très simplifiée des résultats connus.
Répétition et spécialisation
De nombreux métazoaires sont construits sur un modèle de répétition sérielle (que ce soit par exemple des segments, comme chez les arthropodes, ou des structures comme les vertèbres et les myomères chez les vertébrés). Ici aussi, la redondance due à la répétition des structures permet des modifications de quelques unes, qui pourront mener à une spécialisation de certaines d’entre elles.
Des additions de nouvelles régions régulatrices à certains gènes homéotiques ont permis leur utilisation à différents niveaux au cours du développement, d’une première fonction générale à des endroits et des spécialisations de plus en plus fines. Par exemple, le gène Distal-less évoqué plus haut est également activé à certains endroits lors de la formation des ailes de papillons, et initie la formation des taches imitant des yeux sur les ailes.
Spéciation
Que les mutations et modifications génétiques soient fixées par la dérive, ou par la sélection n’est pas forcément important pour comprendre les mécanismes de spéciation. Par contre, il est important de se souvenir que même des populations parfaitement identiques au départ, maintenues séparées dans des conditions semblables, vont progressivement diverger les unes des autres, à cause de la nature aléatoire de la création et de la fixation de la variabilité. C’est ce qu’on appelle le principe de divergence ; qui provient du simple fait que les individus varient aléatoirement au cours du temps.
L’espèce
Une espèce est définie par une communauté d'ascendance (filiation). A l’intérieur d’une espèce, les relations sont de type réseau (reproduction et échange de matériel génétique : continuum de flux généalogique), tandis qu’entre espèces, les relations vont former des branches distinctes (un arbre). La raison en est simple : si une barrière empêche des individus d’échanger leurs gènes, le principe de divergence va aboutir tôt ou tard à ce que les individus ne se reconnaîtront plus comme partenaires sexuels, jusqu’à l’interstérilité, voire directement à une interstérilité. Une espèce est donc définie comme un continuum généalogique, c’est-à-dire un internœud sur un arbre généalogique théorique (voir fig. 4) : d'une rupture dans le flux généalogique jusqu’à la rupture suivante. Au sein de ce continuum, tout individu est interfécond avec les autres et peut potentiellement donner une descendance fertile.
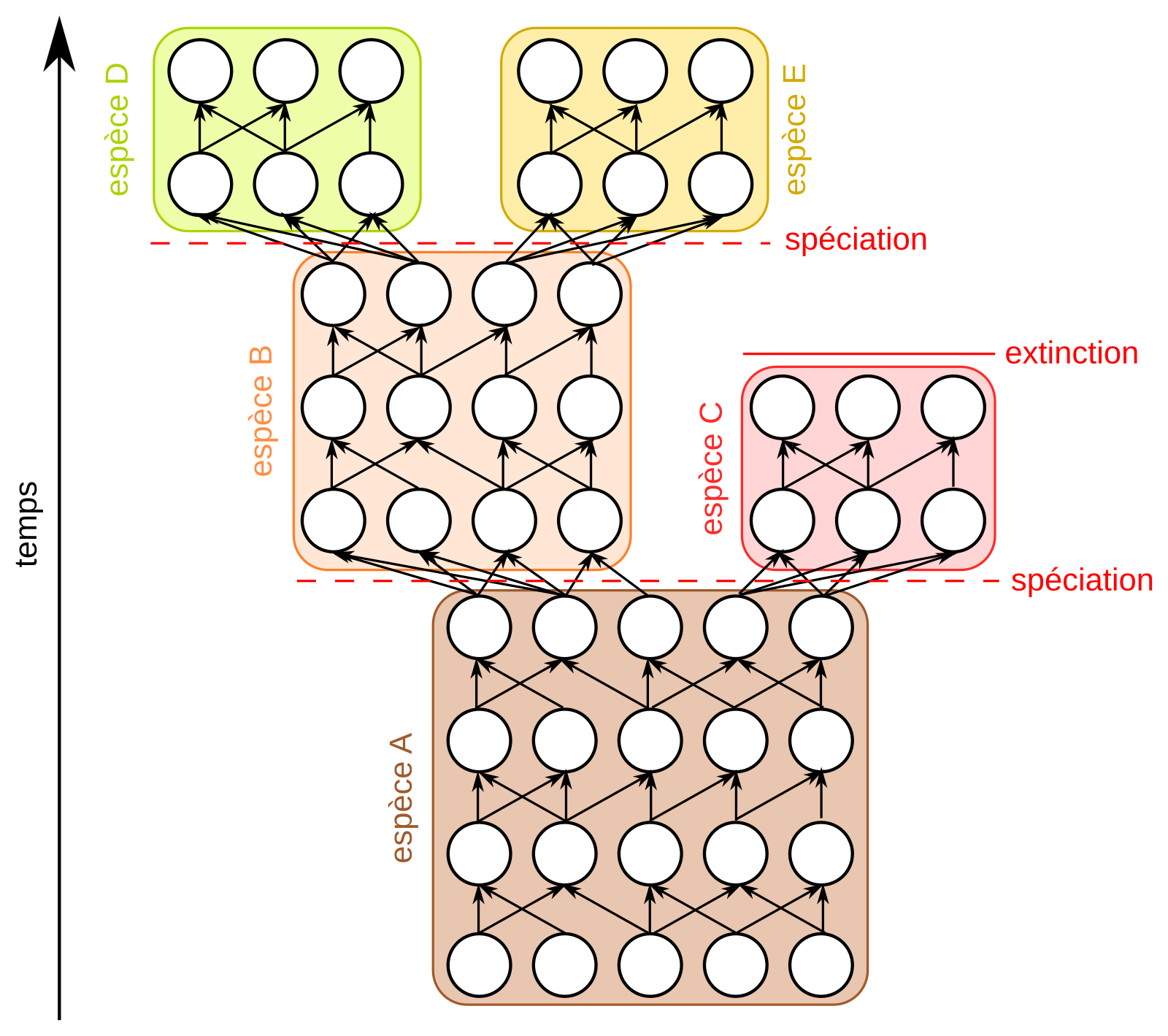
Chaque rond représente un individu. Les flèches représentent les liens de parenté entre individus. Si deux populations ne peuvent plus se reproduire, on considère qu’il y a formation de deux nouvelles espèces.
Bien que l’espèce soit la seule catégorie taxinomique qui peut être considérée comme ayant des bases biologiques, aucun des critères biologiques pour la reconnaître n’est applicable facilement à tous les groupes d’êtres vivants, et certains ne sont utilisables que pour des groupes restreints d’organismes. Il n’y a donc pas une seule, mais de très nombreux critères de reconnaissance de l’espèce (Lherminier et Solignac 2005), et les limites de l’application de chacun font aujourd’hui encore l’objet d’un intense débat.
Un seul critère de reconnaissance ne suffit pas, du fait de la diversité des modes de reproduction et des modes d’échange de matériel génétique, et surtout de la variabilité des niveaux auxquels lesdits échanges cessent/deviennent impossibles. Il n’y a pas un niveau unique de divergence, qu’elle soit morphologique ou moléculaire, qui pourrait servir à définir une espèce, ou à prédire si des individus appartiennent à des espèces différentes. Il n'est pas possible de dire : il y a tant de pourcentages de différence au niveau de la morphologie ou du génome, donc il s'agit de la même espèce (ou d'espèces différentes).
Même si la difficulté d'application des critères peut rendre difficiles voire impossible les
Pour le programme de terminale, deux critères ont été retenus. La plupart des critères de reconnaissance rentrent dans l’une des trois catégories suivantes : les critères biologiques, les critères morphologique, et les critères phylogénétiques. (voir "Les critères de reconnaissances de l'espèce'").
Types et mécanismes de spéciation
Il existe au moins deux types de spéciation.
Spéciation allopatrique
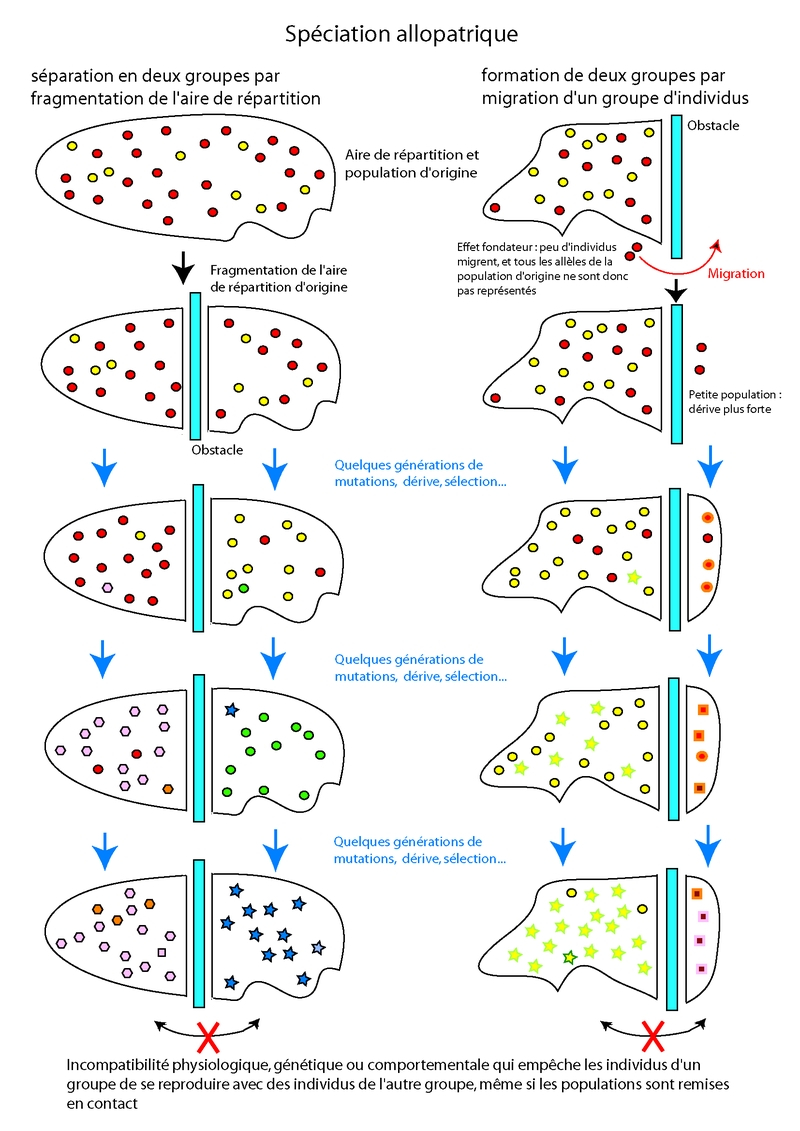
La spéciation allopatrique est probablement la plus courante. Elle se produit entre des populations qui ont été isolées dans l’espace, par fragmentation de l’habitat (une zone de distribution est séparée en deux par l’apparition d’un obstacle) ou migration, et qui du fait de ces obstacles ne peuvent plus échanger de matériel génétique. Cet isolement reproductif du à des circonstances externes permet une évolution indépendante et une divergence des populations, qui finissent par acquérir un isolement reproductif intrinsèque (l’interfécondité étant compromise par une ou plusieurs différences morphologiques, génétiques ou comportementales).Voir fig. 5.
Spéciation sympatrique
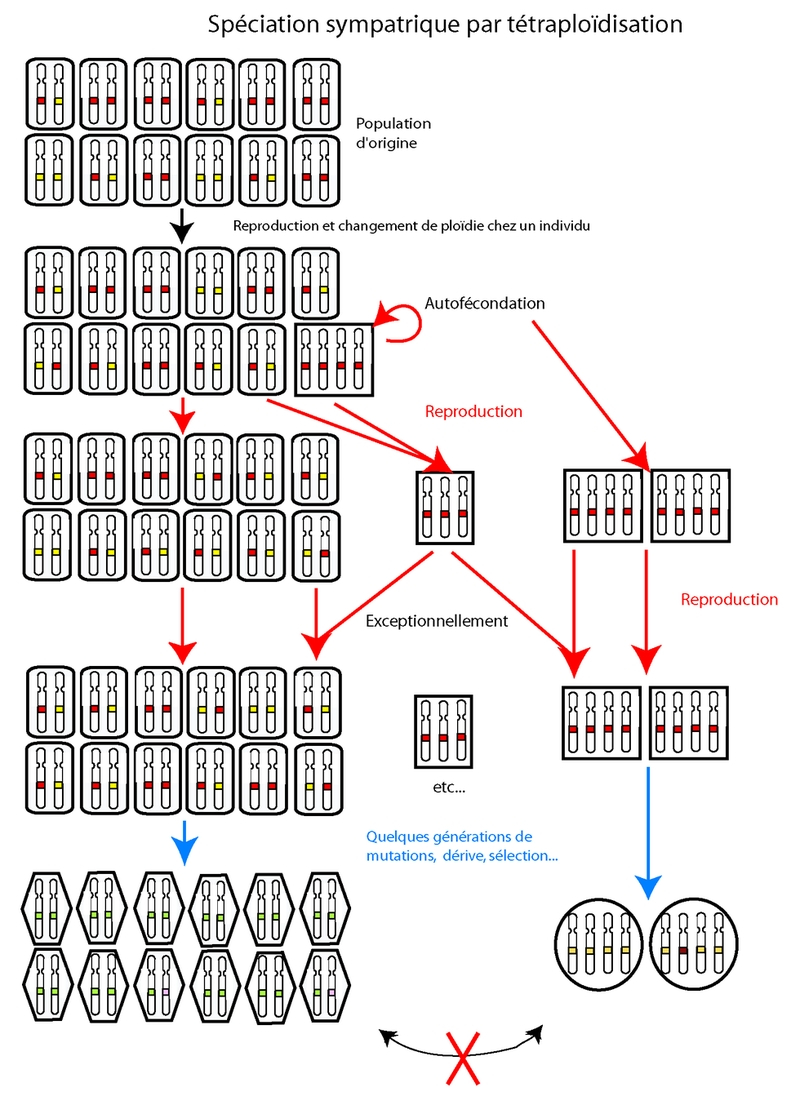
La spéciation sympatrique se fait par apparition d’une nouvelle espèce sur une aire de répartition chevauchante avec celle de l’espèce d’origine. Ce type de spéciation suppose qu’une barrière intrinsèque à la reproduction se mette en place très rapidement, puisque les deux groupes sont en contact. Les changements de ploïdie, par exemple, peuvent servir de telles barrières (un tétraploïde qui apparaît dans une population diploïde, par exemple). Dans les cas d’hybrides de parents avec des nombres chromosomiques différents, une tétraploïdisation (allotétraploïde) peut permettre la survie et la fertilité de l’hybride. Un certain nombre d’exemples existent chez les plantes à fleur et les fougères actuelles (certains chercheurs ont estimé qu’ils sont responsables de 2 à 4 % des événements de spéciation chez les plantes à fleur, et de 7% chez les fougères ; Otto et Whitton 2000). L’un des problèmes rencontrés par les polyploïdes nouvellement formés est le manque de partenaires pour la reproduction : un croisement entre un individu diploïde et un individu tétraploïde donnera des triploïdes, qui ont généralement une fécondité très réduite du fait de problèmes de ségrégation des chromosomes. Il arrive cependant que les triploïdes produisent des gamètes euploïdes (haploïdes, diploïdes et triploïdes) en très faible proportion. Ils peuvent alors produire avec l’individu tétraploïde quelques descendants tétraploïdes ; les organismes capables d’autofertilisation peuvent également contourner ce problème. La polyploïdisation peut produire des changements importants dans la morphologie. Voir fig. 6.
Extinction
Les archives fossiles sont remplies d’exemples qui montrent que des espèces et des groupes d’espèces disparaissent, souvent sans laisser de descendants. On peut citer ainsi les trilobites, la plupart des groupes de dinosaures, les placodermes, les belemnites…
Extinction : disparition complète d’une espèce ou d’un groupe d’espèces. Une espèce est considérée comme éteinte lorsque le dernier individu meurt.
Les extinctions peuvent avoir de nombreuses raisons, et il est souvent difficile de déterminer laquelle s’applique à un cas particulier. D’une manière générale, l’extinction se produit lorsqu’une espèce qui ne peut plus survivre ou se reproduire dans un milieu, et est incapable de gagner un nouveau milieu qui lui conviendrait.
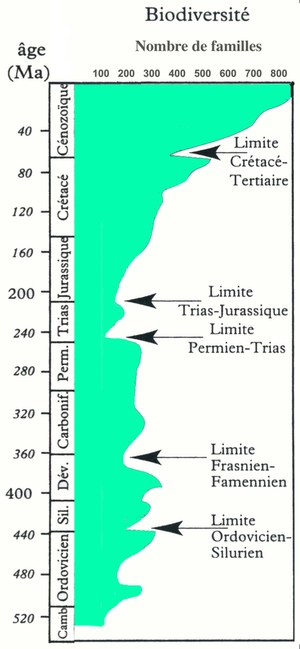
Les limites sont mises en évidences par la diminution rapide du nombre de familles.
Elles peuvent être dues par exemple à des changements brutaux du milieu (c’est le cas de nombreuses espèces de grenouilles, de nos jours), de la prédation (on peut évoquer le cas du dodo), ou de la compétition (certaines espèces introduites se multiplie rapidement, au détriment d’espèces locales : grenouille taureau, caulerpe…). Des maladies peuvent également être impliquées (la grippe aviaire qui a récemment causé la mort de plusieurs tigres vivant en liberté), ainsi que des causes génétiques, qui frappent particulièrement les petites populations. Par exemple la consanguinité et/ou la dérive peuvent être responsables de la fixation de mutations délétères. Enfin, l’extinction peut également être due à la disparition ou extinction d’une ressource dont dépendait l’espèce pour sa survie ou sa reproduction : pollinisateur, proie… Bien entendu, il est également possible que plusieurs facteurs se combinent…
Un grand nombre d’extinctions se sont produites au cours de l’histoire des êtres vivants. Nombre d'entre elles ont été retenues comme marqueurs des limites entre les différentes ères géologiques (voir fig. 7). Nous vivons actuellement la sixième grande extinction.