Célèbre pour ses conséquences sur les dinosaures non aviaires, la crise Crétacé-Paléogène, il y a 66 millions d'années, a très fortement affecté le plancton. Très diversifiés au Crétacé supérieur, les foraminifères planctoniques vont brutalement se raréfier à la base du Paléogène. Les modalités de traversée de la crise par ce groupe font toujours débat.
Les foraminifères sont des microorganismes unicellulaires, principalement marins, fabriquant dans leur grande majorité un test calcaire formé de plusieurs loges. Les loges communiquent entre elles par des ouvertures (d’où le nom de foraminifères, dérivé du latin foramen, trou). Le cytoplasme sort par une ou plusieurs grandes ouvertures et recouvre complètement le test. Les extensions du cytoplasme sont utilisées pour se déplacer et se nourrir.
Au cours de leur évolution, les foraminifères ont su exploiter une grande variété de mode de vie, et par suite occuper différentes niches écologiques (benthiques et planctoniques). Ils sont largement employés comme outils biostratigraphiques et de reconstitutions paléoenvironnementales. En effet, leur test minéralisé est généralement très bien conservé et de petites quantités de sédiments suffisent à en extraire un grand nombre.
Depuis la partie moyenne du Jurassique, les foraminifères planctoniques font partie des principaux producteurs de carbonates dans les océans. Après leur mort, leurs coquilles chutent dans la colonne d’eau et se déposent sur le fond pour former une boue carbonatée. De nos jours, les foraminifères planctoniques vivent pour la plupart dans la zone euphotique (entre dix et cinquante mètres de profondeur). Certaines espèces sont, en revanche, capables de descendre dans la tranche d’eau lors de leur cycle de vie et d’atteindre plusieurs centaines de mètres de profondeur. Les espèces à morphologie globuleuse effectuent majoritairement leur cycle de vie dans la zone euphotique alors que les espèces carénées (à une carène) sont présentes dans les masses d’eaux océaniques profondes.
Alors que les foraminifères benthiques semblent avoir été relativement peu affectés par la crise Crétacé-Paléogène, il y a 66 millions d’années, les formes planctoniques connurent quant à elles une crise majeure, que nous présentons dans cet article.
À la fin du Crétacé, une incroyable diversité
Au Crétacé supérieur, la distribution des foraminifères planctoniques dans la tranche d’eau est supposée similaire à celle observée dans les océans modernes, avec des formes globuleuses à morphologie simple en surface et des formes plus complexes, à une ou deux carènes, en profondeur (Figure 1). La carène est un bourrelet calcitique, présent sur la majorité des loges, et qui permettrait au foraminifère d’alourdir son test pour faciliter la plongée dans la tranche d’eau.
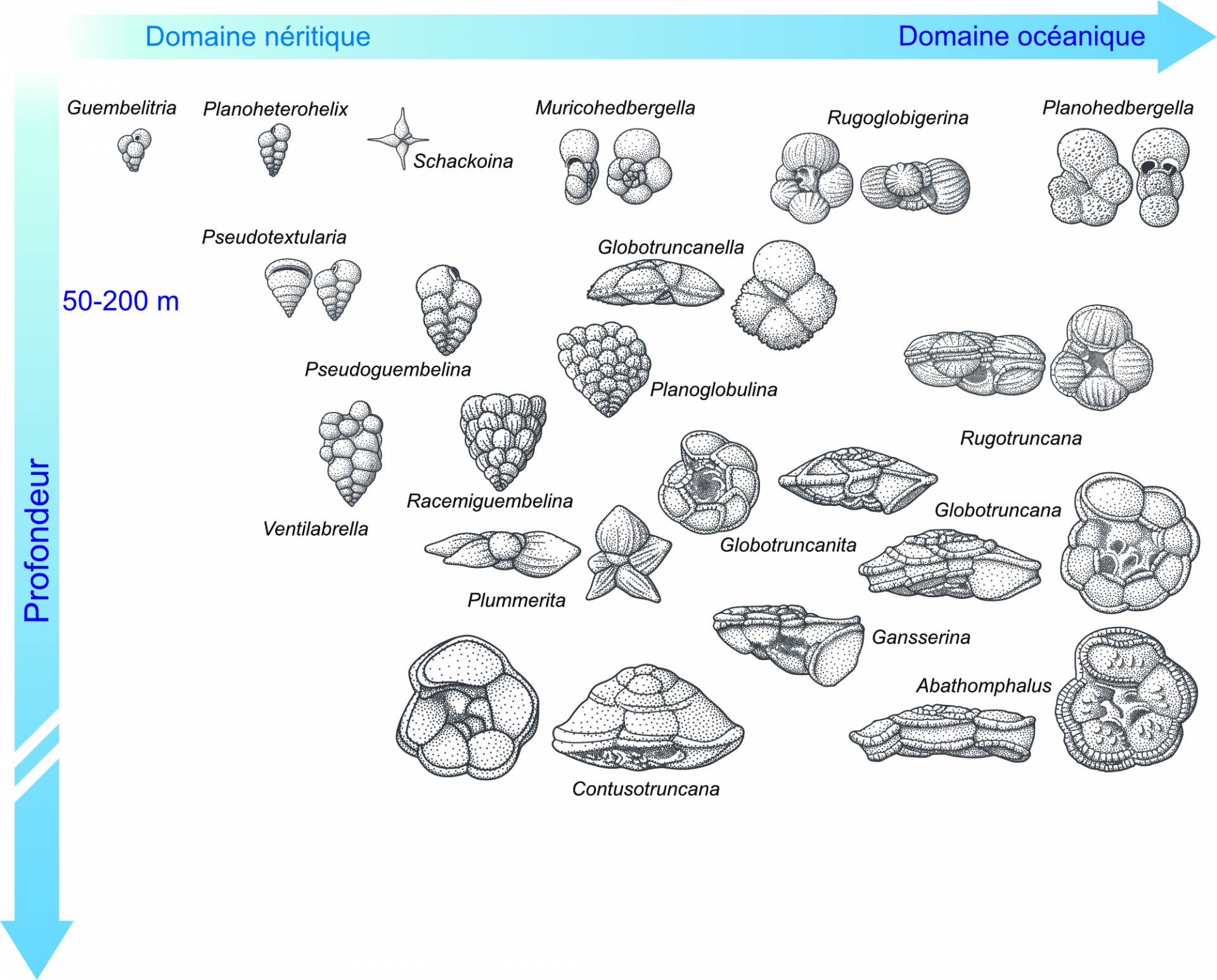
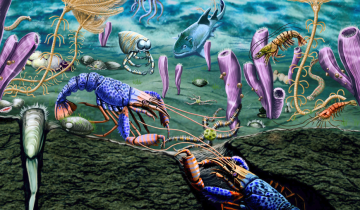
Au Maastrichtien supérieur, c’est-à-dire à la toute fin du Crétacé, les assemblages de foraminifères planctoniques aux basses latitudes sont particulièrement diversifiés. Cela se marque par une incroyable variété de formes (Figures 1 et 2). La fin du Crétacé compte ainsi parmi les intervalles de temps (avec la partie moyenne de l’Éocène) où la diversité des foraminifères planctoniques est la plus forte. Parmi les formes trochospiralées, c’est-à-dire dont la coquille s’organise en spirale non plane (Figure 2), existent des morphotypes globuleux (comme les genres Muricohedbergella ou Rugoglobigerina), des morphotypes à une seule carène (comme les genres Globotruncanita ou Gansserina) et d’autres à deux carènes (comme les genres Contusotruncana et Globotruncana). La présence de deux carènes est d’ailleurs une spécificité du Crétacé supérieur. C’est aussi l’intervalle de temps durant lequel les foraminifères planctoniques atteignent leur plus grande taille, avec Contusotruncana contusa qui pouvait mesurer jusqu’à un millimètre. Cette espèce présentant deux carènes est facilement reconnaissable à sa trochospire très élevée (Figure 2). Les assemblages de la partie terminale du Crétacé ont aussi la particularité de contenir de très nombreuses formes sériées aux morphologies multiples et, parfois, exubérantes (Figures 1 et 2). Parmi ces formes existent des genres bisériées1 (Planoheterohelix ou Pseudotextularia), des formes trisériées (Guembelitria) ou encore plurisériées (Racemiguembelina ou Planoguembelina).
Les formes aux architectures les plus complexes (Contusotruncana, Racemiguembelina ou Planoguembelina), qui vont s’éteindre au passage Crétacé-Paléogène, n’ont aucun analogue au Cénozoïque. La crise Crétacé-Paléogène ne se réduit donc pas seulement, comme on peut le lire dans certains ouvrages, à la disparition des Globotruncana, mais elle voit la disparition quasi-totale des riches assemblages de foraminifères planctoniques. L’impact de la crise sur les foraminifères planctoniques est bien illustré à El Kef (Tunisie). Ce site correspond au point stratotypique mondial (PSM ou, en anglais, GSSP, Global boundary Stratotype Section and Point), c’est-à-dire au stratotype de référence pour la base du Danien (premier étage du Paléogène) datée à – 66 Ma (Figure 3). Seules, 3 à 4 espèces sur une cinquantaine vont survivre à cette crise. Ainsi, plus de 90 % des espèces s’éteignent.

Micrographies obtenues au microscope électronique à balayage. Les espèces figurées proviennent du Maastrichtien supérieur de la coupe type de Bidart (Pyrénées Atlantiques, France). Spécimens 1 à 3 : Globotruncanidae (formes trochospiralées) illustrés sur le profil (a) et la face spirale (b). 1. Contusotruncana contusa. 2. Globotruncana arca. 3. Globotruncanita stuarti (− 15 cm sous la limite Crétacé-Paléogène). Spécimen 4. Rugoglobigerinidae (forme trochospiralée), Rugoglobigerina rugosa (− 180 cm sous la limite Crétacé-Paléogène). Spécimens 5 à 6 : Heterohelicidae (formes sériées). 5. Pseudotextularia elegans. 6. Racemiguembelina fructicosa (− 180 cm sous la limite Crétacé-Paléogène).
La crise Crétacé-Paléogène, crise du plancton calcaire
L’une des cinq grandes extinctions de masse de l’histoire de la Terre s’est produite à la fin du Crétacé, il y a soixante-six millions d’années. Si cette crise n’est pas la plus sévère en termes de pourcentage de famille et de genre disparus, la disparition de groupes emblématiques tels que les dinosaures non aviaires ou les ammonites a fait d’elle la crise la plus médiatisée. Cette médiatisation a été alimentée par les vifs débats qui ont animé la communauté scientifique concernant la cause de cette crise biologique majeure, en raison de l’occurrence simultanée, près de la limite Crétacé-Paléogène, de l’impact d’un astéroïde et d’un volcanisme intense. Les rôles relatifs de l’impact de l’astéroïde de Chicxulub (plateforme du Yucatan, Mexique) et des trapps du Deccan en Inde en tant que mécanismes d’extinction sont compliqués à démêler du fait des incertitudes inhérentes au registre géologique (enregistrement et chronologie des différents événements et de leurs conséquences respectives). Toutefois, des travaux récents de modélisation tendent à démontrer quantitativement que cette extinction aurait été principalement induite par l’impact de l’astéroïde 12. Ces nouveaux modèles et simulations suggèrent même que les effets d’un volcanisme actif pourraient avoir favorisé la récupération après l’extinction de masse induite par l’impact hivernal.
La crise Crétacé-Paléogène affecte particulièrement les producteurs carbonatés pélagiques, à savoir les coccolithophoridés sensu lato et les foraminifères planctoniques. La crise est ainsi marquée par l’effondrement de la production de carbonate de calcium par le plancton marin. La limite Crétacé-Paléogène voit donc une extinction brutale (en un laps de temps très court) et le renouvellement quasiment complet du microplancton et du nannoplancton calcaire. Les foraminifères planctoniques sont affectés à un haut rang taxinomique puisque 2/3 des familles (4 sur 6) disparaissent, à l’échelle globale, à la limite (ou peu après), dont la fameuse famille des Globotruncanidae regroupant la plupart des genres à deux carènes (Globotruncana et Contusotruncana). D’une manière générale, toutes les grandes formes, à l’architecture complexes, carénées et/ou ornementées sont exterminées (Figure 3). Les survivants avérés sont des petites formes opportunistes de surface, à la morphologie simple, appartenant aux deux familles de foraminifères planctoniques sériés (Heterohelicidae et Guembelitriidae). Il s’agit des deux espèces : Planoheterohelix globulosa et Guembelitria cretacea. Cette dernière espèce prolifère peu de temps avant la limite et est particulièrement abondante à la base du Danien 3. Bien représentée pendant l’intervalle de crise, cette petite forme trisériée est désignée comme une espèce « désastre » : les Guembelitriidae sont considérées comme des formes résistantes aux stress environnementaux ; leur prolifération est donc un marqueur de ces intervalles de crises (Keller & Pardo, 2004). D’autres petites formes globuleuses de surface (par exemple Muricohedbergella monmouthensis et Muricohedbergella holmdelensis) auraient pu survivre quelque temps (Figure 3). Des formes maastrichtiennes, plus complexes, se retrouvent également dans les sédiments de la base du Danien. Le statut de ces taxons (survivants ou remaniés, voir partie 4) a été, et est encore, source de débat (Figure 3).
Le schéma classique proposé pour la reconquête de la tranche d’eau par les foraminifères planctoniques est celui d’une recolonisation initiale par les quelques survivants à la crise Crétacé-Paléogène, suivie de deux phases de diversification 4. La première phase, dans le premier demi-million d’années suivant la limite, conduit au retour des morphologies basiques. Le nombre de genres augmente, mais le nombre d’espèces dans chaque genre reste relativement faible. La seconde phase de diversification, qui aura lieu près de 4 millions d’années après la limite Crétacé-Paléogène, voit la mise en place des principales lignées du Paléogène et le retour des formes carénées (à une carène). La diversité spécifique augmente alors rapidement pour atteindre une quarantaine d’espèces.
Les travaux de Morard et ses collaborateurs remettent toutefois en question l’interprétation classique suggérant que les foraminifères planctoniques mésozoïques survivants ont donné naissance aux communautés modernes 5. Leurs données de phylogénie moléculaire montrent que les nouveaux clades cénozoïques auraient émergé d’ancêtres benthiques qui auraient colonisé la niche planctonique après la crise. Une telle hypothèse avait déjà été avancée par des travaux antérieurs 67 illustrant que la classification traditionnelle des foraminifères, séparant les foraminifères planctoniques dans le sous-ordre des Globigerinina et les foraminifères benthiques dans d’autres sous-ordres, ne reflétait probablement pas la phylogénie réelle de ces taxons. En effet, des preuves génétiques surprenantes montrent que dans les océans actuels, Streptochilus globigerus, planctonique bisérié, appartient à la même espèce biologique qu’une forme benthique avérée : Bolivina variabilis 8. S. globigerus / B. variabilis est donc une espèce écologiquement flexible qui peut occuper à la fois les domaines pélagiques et benthiques. Un tel mode de vie, qualifié de tychopélagique, constitue un avantage écologique car il permet la recolonisation rapide du domaine pélagique à partir du benthos après les événements de crise.
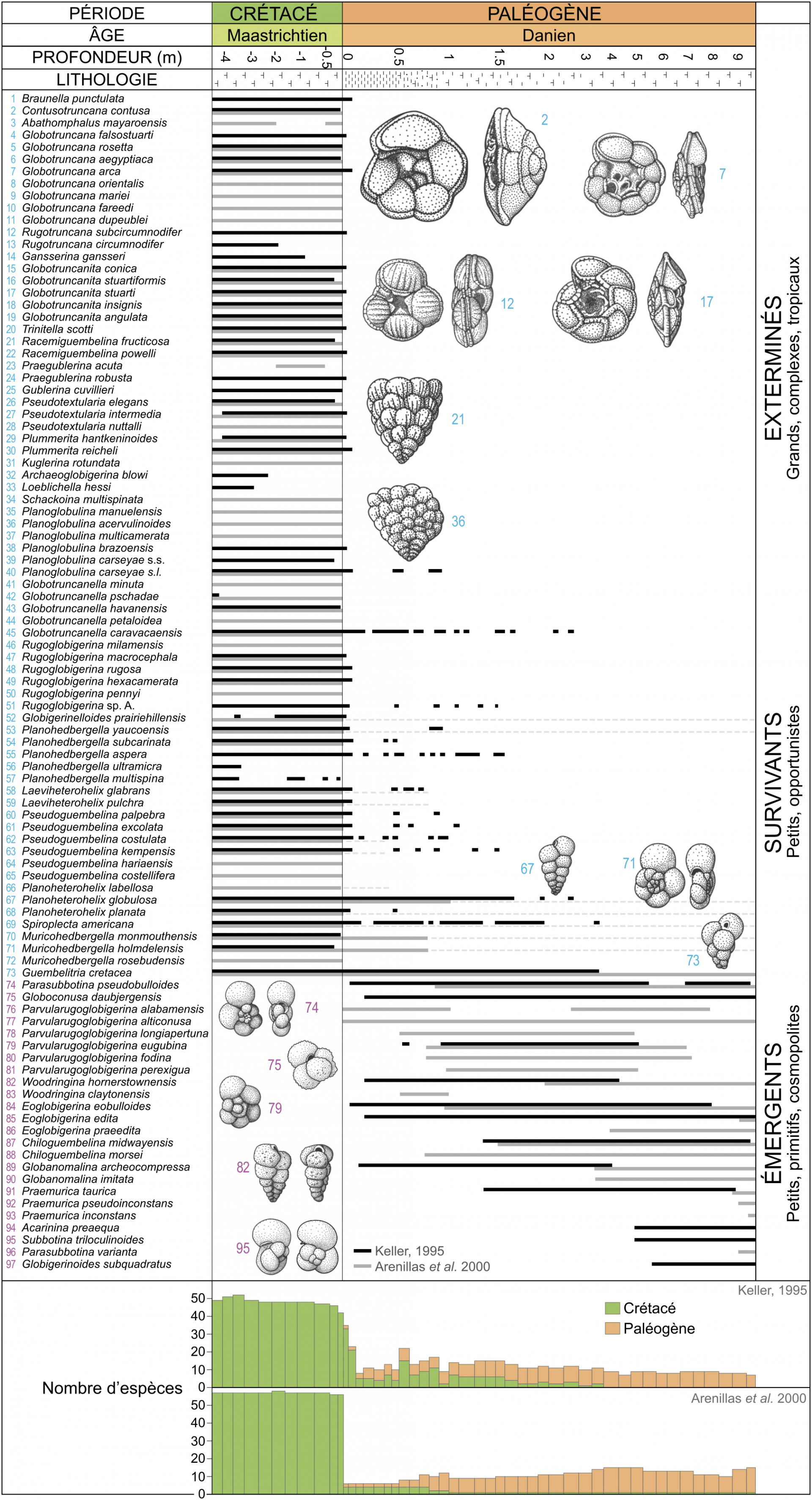
Les traits noirs indiquent les extensions stratigraphiques proposées par Keller (1995). Les traits gris correspondent aux extensions stratigraphiques d’Arenillas et coll. (2000) ; les pointillés gris correspondent aux occurrences considérées comme remaniées par ces mêmes auteurs. Les deux dénombrements des espèces (diversité) ont été effectués en suivant les travaux des deux équipes (Keller, 1995 vs Arenillas et coll., 2000), l’une (Keller, 1995) considère que les formes crétacées présentes au début du Paléogène sont des formes survivantes alors que l’autre (Arenillas et coll., 2000) estime qu’il s’agit majoritairement de spécimens remaniés.
Une limite qui se voit bien sur le terrain
Sur le terrain, lorsque les coupes sont complètes, la limite Crétacé-Paléogène est souvent marquée par de brusques changements lithologiques ; les séries carbonatées sont ainsi interrompues par un fin niveau argileux, dépourvu de carbonates (crise des producteurs) et contenant l’anomalie en iridium. C’est notamment le cas pour la coupe de référence de la Contessa, située dans la région de Gubbio en Italie (Figures 3 et 4). Le nom de cette coupe est connu de tous les géologues car c’est sur ce site que les Alvarez, père et fils, accompagné de Frank Asaro et d’Helen Michel ont mis en évidence dans les années 1980 la fameuse anomalie en iridium, permettant d’avancer une origine extraterrestre pour expliquer la crise Crétacé-Paléogène.

À la Contessa, une fine couche d’argiles sépare les calcaires rose clair de la fin du Crétacé des calcaires rose foncé du début du Paléogène. L’observation des calcaires à la loupe, de part et d’autre de la couche d’argiles, permet de visualiser l’impact de la crise sur les foraminifères planctoniques. Avec une loupe de terrain, ce que l’on observe est analogue à la première colonne de la figure 5. Ainsi dans l’échantillon crétacé, de nombreux petits grains sont visibles à la loupe. Ce sont des foraminifères planctoniques. Sans être spécialiste, on peut noter la variété de forme et de taille de ces grains qui rend compte de la diversité des assemblages de la partie terminale du Crétacé. Un centimètre au-dessus de la couche d’argiles, l’échantillon du Paléogène parait homogène et, on y perçoit difficilement quelques grains. En quelques centimètres seulement, les riches assemblages du Maastrichtien disparaissent brutalement. Ce constat est encore plus flagrant à fort grossissement, au microscope. L’observation des lames minces montre des foraminifères planctoniques qui sont jusqu’à 10 fois plus petits à la base du Danien (Figure 5).


L’assemblage du Crétacé est entièrement constitué de foraminifères planctoniques, seuls deux foraminifères benthiques sont visibles (notés Bth). Quelques centimètres au-dessus les formes planctoniques sont très rares (un seul spécimen, noté Plc, visible sur le cliché).
La coupe de Bidart (Pyrénées-Atlantiques) est aussi un site de référence pour l'étude de la limite Crétacé-Paléogène. Dans les séries marno-calcaires qui affleurent le long de la côte, au sud de Biarritz, les lavages (tamisages) des sédiments permettent de récolter facilement les microfossiles. L’observation des résidus de tamis de part et d’autre de la limite (Figure 6) permet également d’illustrer cette crise. En effet, les assemblages sont totalement différents. Pour la même quantité de sédiments tamisés, la quantité de microfossiles collectés est beaucoup moins importante à la base du Danien. Les derniers centimètres du Maastrichtien sont principalement composés de foraminifères planctoniques alors que dans les sédiments de la base du Danien il ne reste plus que des foraminifères benthiques, les formes planctoniques sont anecdotiques (Figure 6).
Débat : survivants ou remaniés ?
Les travaux sur les foraminifères planctoniques au passage Crétacé-Paléogène sont nombreux mais contrastent à la fois sur l’interprétation des modalités d’extinction (graduelle ou catastrophique) mais aussi sur le nombre d’espèces survivantes. En effet l’observation d’une espèce donnée dans des roches du Danien peut s’expliquer de deux façons : soit l’espèce était réellement présente à cette période (espèce survivante) soit son fossile, déposé au Maastrichtien, s’est retrouvé plus tard mélangé aux sédiments du Danien, par exemple à cause d’un phénomène de bioturbation (déplacement de sédiments sous l’action d’organismes vivants). Une difficulté pour les équipes de recherche est donc de discriminer les espèces survivantes des espèces remaniées.
La figure 3 confronte les dénombrements d’espèces obtenus sur la coupe d’El Kef, GSSP de la base du Danien, par deux équipes différentes 12. La première observation qui peut être faite, c’est que, pour une même coupe, le nombre d’espèces dénombrées diffère selon les auteurs. Cela peut être lié au pas d’échantillonnage, aux méthodes d’extraction et d’observation des foraminifères, mais aussi aux interprétations taxinomiques qui diffèrent selon les auteurs. Si Keller considère que les formes crétacées présentes au début du Paléogène sont des formes survivantes 3, Arenillas et ses collaborateurs estiment qu’il s’agit en grande majorité de spécimens remaniés 4. Pour les sites comme ceux de la région de Gubbio, où des lames minces peuvent être facilement réalisées dans des calcaires lithographiques, le statut de ces formes peut être vérifié. Par exemple, dans la lame présentée sur la figure 7, les occurrences de formes crétacées dans le Danien sont visibles dans un terrier. Cet exemple illustre que la bioturbation est un phénomène commun qui peut remanier du matériel crétacé dans les premiers centimètres de sédiments daniens. Dans les séries marneuses, comme pour la coupe d’El Kef où les lames minces sont difficilement réalisables, la mise en évidence de bioturbation est moins évidente.
Cependant, quel que soit le statut de ces rares formes crétacées encore présentes au début du Danien, cela ne change en rien la sévérité de la crise pour les foraminifères planctoniques puisque dans les deux cas de figure, on passe d’une cinquantaine d’espèces à la fin du Maastrichtien à moins d’une dizaine à la base du Danien (Figure 3).
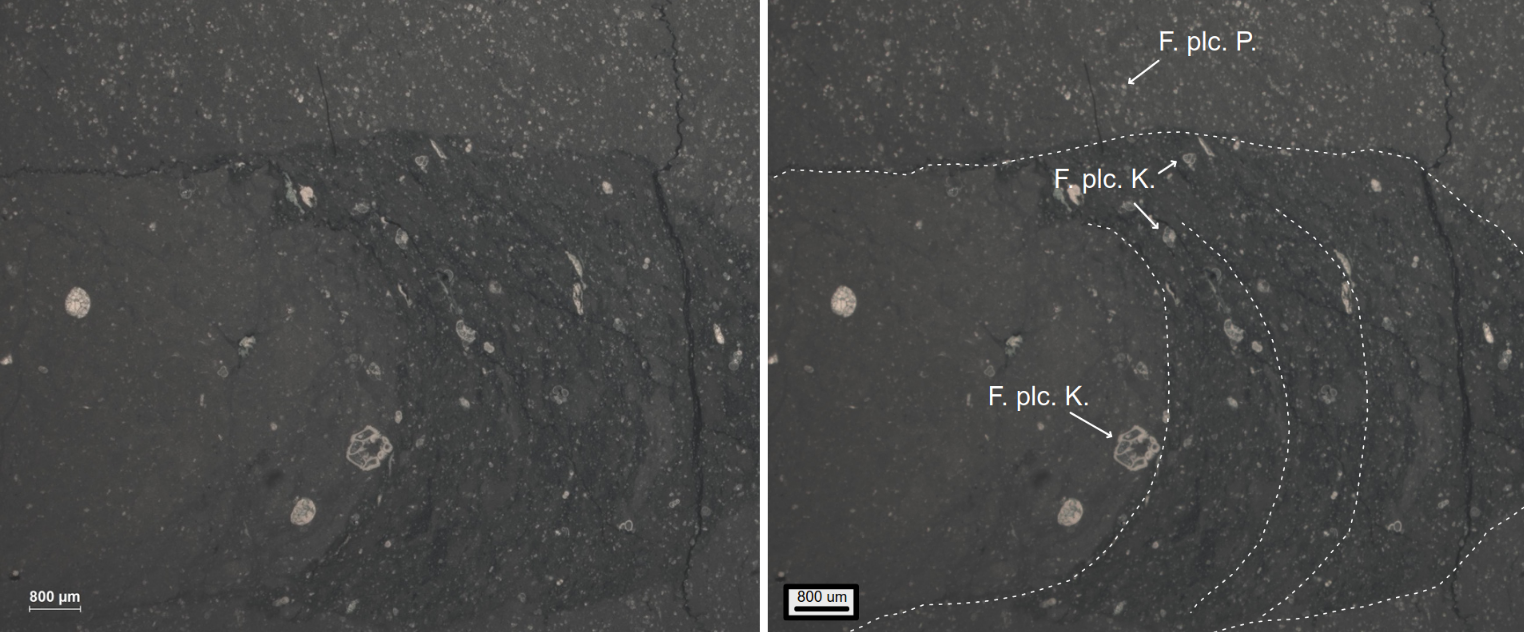
Cette lame mince illustre bien le remaniement de grandes formes crétacées (F. plc. K.) dans les sédiments paléogènes exclusivement composés de petits formes (F. plc. P.).