
Alors qu'ils forment un groupe peu diversifié aujourd'hui, les Crocodyliformes présentaient dans le passé des morphologies, des régimes alimentaires et des milieux de vie beaucoup plus variés qu'actuellement. Contrairement à d'autres taxons occupant les mêmes biotopes, le groupe des Crocodyliformes a été peu affecté par la crise Crétacé-Paléogène. Comment l'expliquer ?
Les crocodiles actuels (au sens large : crocodiles, gavials, alligators…) font partie du grand groupe des Archosauriens, dont seuls subsistent actuellement les Oiseaux et les Crocodiliens1. Ils ont une diversité extrêmement pauvre, avec seulement 24 espèces distribuées principalement dans les zones intertropicales. En effet, leur physiologie poïkilotherme limite leur présence aux climats chauds, leur taille ne leur permettant pas d’hiberner, tout comme elle restreint leur degré d’activité. Leur diversité morphologique est elle aussi extrêmement faible, avec des formes amphibies, dont les déplacements terrestres sont plutôt malhabiles. En effet leurs membres courts, en position transversale (et non érigés sous le corps comme chez les Mammifères), ne leur permettent pas, sauf exception, de courir, et les obligent lors de la marche à un dandinement caractéristique. Les crocodiles occupent principalement les milieux d’eaux douces et les estuaires, seules une ou deux espèces fréquentent assidûment les milieux marins. Le régime alimentaire carné est de mise, avec quelques formes spécialisées dont la longirostrie (museau fin et allongé) et les dents longues et fines indiquent un régime essentiellement piscivore.

La diversité actuelle des Crocodiliens est donc très limitée. Pourtant celle des Crocodyliformes, groupe beaucoup plus large comprenant les Crocodiliens, était quant à elle extrêmement importante, tant par le nombre d’espèces que par leurs formes. Si plusieurs groupes de Crocodyliformes traversent la crise Crétacé-Paléogène (K-Pg, également appelée abusivement crise Crétacé-Tertiaire ou crise Crétacé-Paléocène1), certains de leurs contemporains, les grands Reptiles marins et terrestres, et notamment les Dinosaures non aviens, s’éteignent. Cela rend l’étude de ce groupe et leur comparaison avec l’évolution des Reptiles contemporains particulièrement intéressantes.
Reptiles ou reptiles ?
Le groupe des reptiles était à l’origine défini en opposition aux Mammifères et aux Oiseaux. Il regroupait donc l’ensemble des Amniotes respirant à l’air libre et ne possédant ni poils, ni plumes, ni mamelles. Cela comprend les Lépidosauriens (lézards, serpents), les Chéloniens (tortues), les Crocodiliens, et les Dinosaures, mais exclut les Oiseaux. Or ces derniers sont inclus dans le groupe des Dinosaures. Les exclure des reptiles revient donc à exclure une partie des descendants de l’ancêtre commun de tous les reptiles, qui forment alors un groupe paraphylétique. Le terme Reptilia ne devrait donc plus être utilisé.
Cependant, en 2004, Modesto et Anderson proposèrent une définition phylogénétique de Reptilia, synonyme de Sauropside. Dans cette acception, le terme de Reptile désigne donc un groupe monophylétique qui contient, en plus des taxons précédemment cités, les Oiseaux.
Une incroyable diversité Mésozoïque
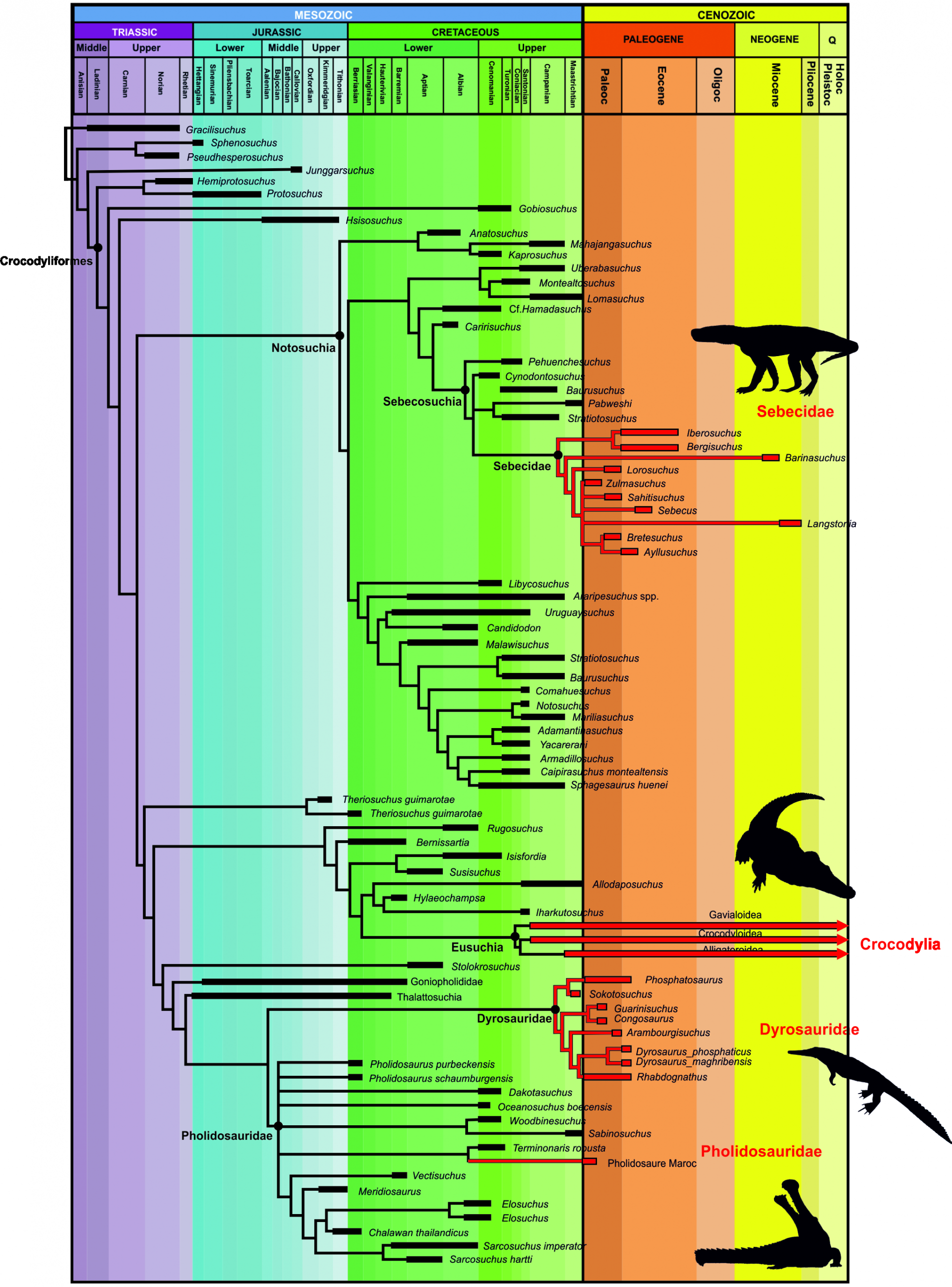
Apparus à la fin du Trias, les Crocodyliformes occupent dès le milieu du Jurassique quasiment tous les milieux, aussi bien marins que continentaux (Wilberg et coll., 2019). À la fin du Crétacé, existent trois grands clades : les Notosuchiens, les Téthysuchiens et les Eusuchiens. Ces trois groupes sont extrêmement différents, et tous les trois traversent la crise Crétacé-Paléogène (Fig. 1).

DGM 299-R, Departamento Nacional da Produção Mineral, Museu de Ciências da Terra, Rio de Janeiro
Les premiers, les Notosuchiens, forment un groupe quasi exclusivement terrestre, seules quelques formes étant fluviatiles (Pol et Leardi, 2015). Leur morphologie est très éloignée des crocodiles amphibies actuels. Leurs membres sont longs et fins, érigés à la verticale sous leur corps, laissant supposer un mode de vie uniquement terrestre et non aquatique. Leur crâne est plutôt compact, avec un museau court et haut, loin des crânes aplatis des crocodiles actuels. Beaucoup d’entre eux, les Sébécosuchiens, ont de longues dents crénelées à la manière des dinosaures Théropodes, et la forme de leur crâne haut et compact, n’est pas si éloignée de la morphologie de ces derniers (Godoy et coll., 2014) (Fig. 3). Ils étaient probablement de redoutables prédateurs, chassant leurs proies à l’affût et courant aisément sur de courtes distances grâce à leurs longs membres verticaux. Parmi les Notosuchiens, bien qu’ayant une morphologie assez similaire, certaines espèces ont des dents de formes et d’implantation particulièrement étranges, et les paléontologues se sont longtemps interrogés sur le mode alimentaire de ces animaux. Leurs dents portent de nombreux bulbes, à la manière des dents de certains Mammifères avec lesquelles elles ont parfois été confondues, quand d’autres espèces ont des dents triangulaires, avec de grosses crénelures orientées non pas antéro-postérieurement comme chez tous les Crocodyliformes, mais en biais par rapport à l’axe du crâne (Iori et coll., 2013). Cette morphologie dentaire complètement atypique est considérée comme une possible adaptation à un régime alimentaire uniquement phytophage – un trait surprenant pour des Crocodyliformes.
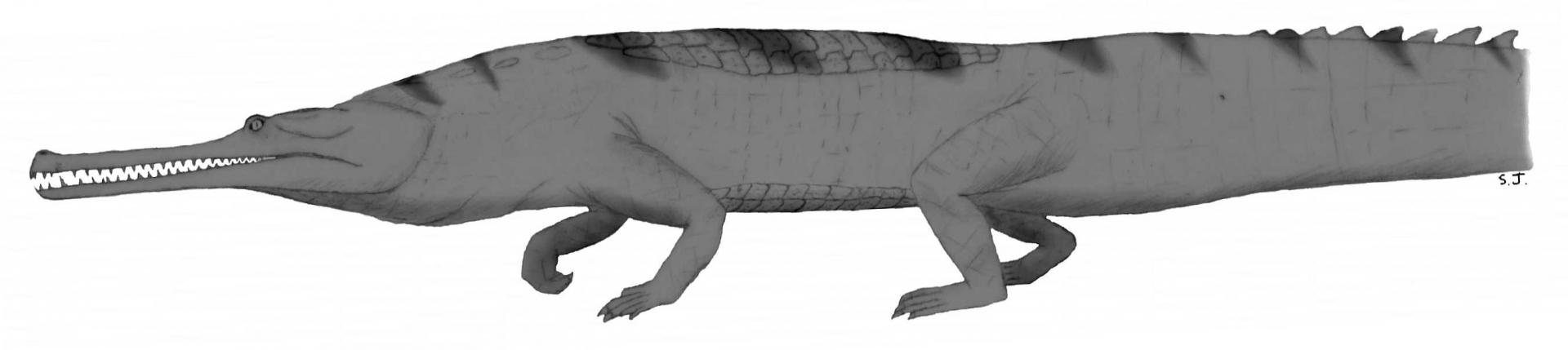
Le second groupe, les Téthysuchiens, est constitué d’espèces uniquement aquatiques. Parmi eux, les Pholidosauridés, présents depuis le Jurassique, comportent des espèces marines et fluviatiles (Martin et coll., 2016). S’ils semblaient disparaître au début du Crétacé supérieur, des découvertes récentes semblent démontrer leur survie à la crise K-Pg. Le second groupe de Téthysuchiens est formé par les Dyrosauridés. Quasi exclusivement constitué de formes vivant dans le milieu marin, ils possèdent une queue très haute et musclée qui devait leur permettre d’être de bons navigateurs (Jouve et coll., 2006) (Fig. 4). Contrairement aux Pholidosauridés dont la diversité est très importante au Crétacé supérieur et s’étiole pour ne laisser qu’une seule espèce à la fin du Crétacé et après la crise K-Pg, les Dyrosauridés semblent peu diversifiés à la fin du Crétacé, et connaissent une véritable explosion après la crise.
Les Eusuchiens, groupe dont font partie tous les crocodiles actuels, apparaissent à la fin du Crétacé et sont présents aussi bien en eaux douces que dans les milieux marins côtiers. Relativement bien diversifiés à la fin du Crétacé, ils ne semblent pas véritablement souffrir de la crise K-Pg, et les trois grands groupes, Gavialoïdes, Alligatoroïdes et Crocodyloïdes, déjà présents au Crétacé supérieur, se diversifient lentement au début du Paléogène.
En résumé, les trois groupes de Crocodyliformes sont présents aussi bien avant qu’après la crise. Ils occupent tous les environnements – fluviatiles, terrestres, marins – ont des régimes alimentaires très variés – carnivores, piscivores, phytophages – et des morphologies allant d’animaux amphibies aux membres courts et transversaux et à queue puissante, à des coureurs aux membres longs et fins, verticaux sous le corps. Cette variété rend les Crocodyliformes particulièrement intéressants à étudier dans le cadre de la crise K-Pg, et la comparaison de leur évolution à travers la crise avec les autres Reptiles contemporains, tels les Dinosaures et Reptiles marins est pertinente pour déterminer l’évolution de la biodiversité à travers la crise, et évaluer les causes des extinctions.
Une survie étonnante
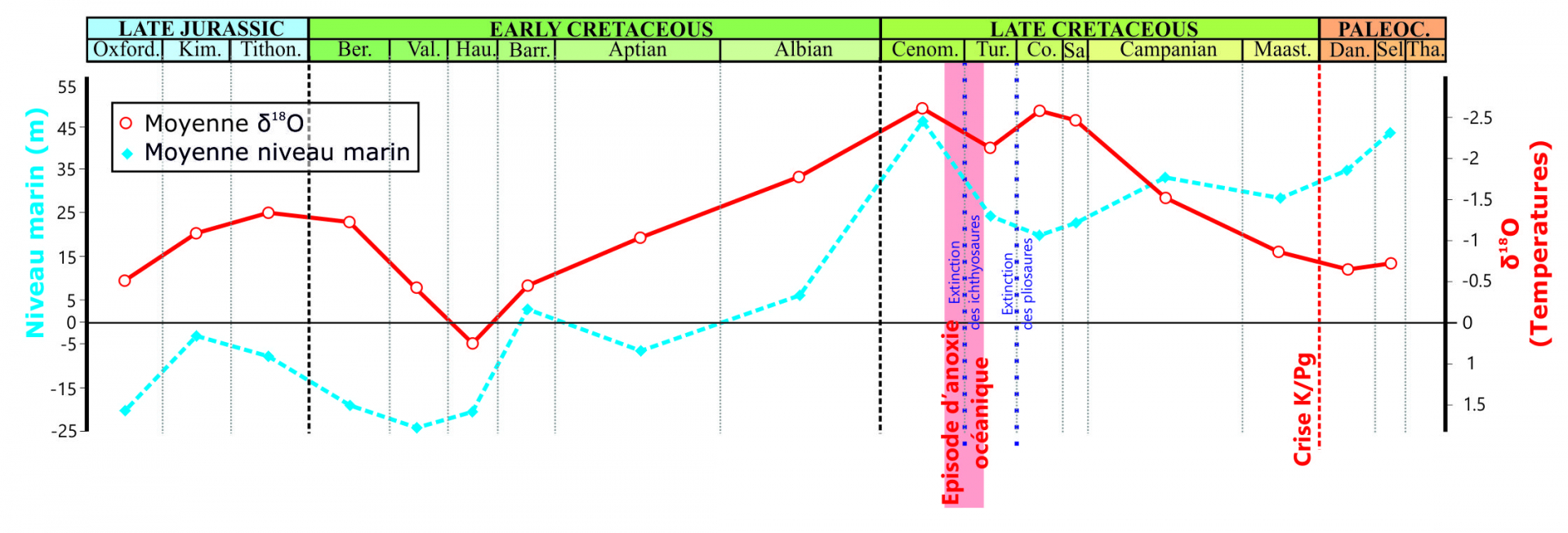
Pour bien comprendre l’évolution des Crocodyliformes, et des Reptiles en général, à travers la crise K-Pg, il faut remonter un peu en arrière pour bien comprendre l’état des différents groupes et leur dynamique lorsque celle-ci survient. Plusieurs phénomènes ont lieu tout au long du Crétacé supérieur. Un fort épisode anoxique (fin Cénomanien-début Turonien) touche les environnements marins, suivi d’une grande régression marine (baisse du niveau marin) au Turonien qui réduit considérablement la surface du plateau continental. La succession des deux phénomènes affaiblit considérablement les écosystèmes marins et en particulier les milieux côtiers (Fig. 5). Les écosystèmes fluviatiles semblent avoir eux aussi été indirectement affectés. Les Pholidosauridés, principalement côtiers et fluviatiles, en subissent les conséquences de plein fouet, et seuls quelques rares taxons survivront jusqu’à la fin du Crétacé. Le climat, très chaud au début du Crétacé supérieur, va ensuite se refroidir progressivement jusqu’au début du Paléogène, impactant considérablement des écosystèmes aquatiques déjà fragilisés, mais atteignant aussi les milieux terrestres. Cette dégradation climatique est probablement à l’origine de la baisse de diversité des Notosuchiens, dont une large partie est éteinte bien avant la crise K-Pg. Seule une partie des Sébécosuchiens (des grands prédateurs), atteignent et traversent la crise K-Pg, avec semble-t-il une radiation au Paléogène (Pol et Leardi, 2015). Les Dyrosauridés et Eusuchiens apparaissent quant à eux à la fin du Crétacé, et connaissent ensuite une lente diversification.
Que se passe-t-il pour les Crocodyliformes lors du passage entre le Crétacé et le Paléogène ? Visiblement, peu de choses négatives ! Les Sébécosuchiens, qui vivent en milieu terrestre, semblent peu affectés et se diversifient fortement en Amérique du Sud après la disparition des concurrents Dinosaures non aviens. Dans le milieu aquatique, les Eusuchiens sont peu perturbés, certaines espèces du Maastrichtien sont même encore présentes au Paléocène. Quant aux Dyrosauridés, leur diversité explose après la crise (Jouve et coll., 2008). Comment expliquer cette disparité entre la survie des Crocodyliformes et la disparition des autres grands Reptiles marins vivant dans les mêmes environnements comme les Dinosaures, Mosasaures et Plésiosaures, des groupes très diversifiés à la fin du Crétacé ?
Tout d’abord, dans les milieux aquatiques, l’évolution des poissons montre une différence assez importante entre les écosystèmes fluviatiles et marins. Quand les milieux et la base des chaînes alimentaires marines souffrent considérablement lors de la crise K-Pg, les écosystèmes fluviatiles semblent ne pas souffrir du tout. Les deux écosystèmes reposent sur une base trophique différente. Quand les milieux marins reposent essentiellement sur les microorganismes planctoniques, eux-mêmes largement dépendant de la photosynthèse et donc de la luminosité, la base de la chaîne alimentaire des écosystèmes fluviatiles s’appuie principalement sur les apports détritiques de matière organique en provenance du continent (Cavin, 2002). Dans le cas d’un impact d’un bolide météoritique, la baisse de luminosité produite par les matières vaporisées a une incidence sur les chaînes trophiques dépendantes de la photosynthèse. Au contraire, celles basées sur les apports de débris organiques seront peu touchées, en particulier par un évènement brutal et court. Les débris organiques continueront à alimenter les milieux fluviatiles pendant des années, même après l’arrêt ou le ralentissement de la production primaire. Or, les Mosasaures et Plésiosaures, à quelques exceptions près, sont des animaux vivants exclusivement dans le milieu marin quand les Eusuchiens n’ont pas de préférences et sont largement présents dans les milieux fluviatiles. Les Dyrosaures quant à eux, s’ils sont essentiellement marins, ont une distribution très restreinte au Maastrichtien, plutôt rare dans les localités où sont présents les Mosasaures. De plus, dans certains gisements marins où les restes de Dyrosaures sont présents en nombre, les restes de juvéniles sont quasi absents. Les pouponnières des Dyrosaures se seraient-elles trouvées en milieux fluviatiles, étant ainsi moins impactées par la crise ? Ceci pourrait expliquer la disparité entre la survie des Dyrosauridés et celle des autres grands Reptiles marins. Une seconde hypothèse pourrait être une différence physiologique. Il a été suggéré que certains Mosasaures auraient pu développer une certaine forme d’homéothermie, alors que les Dyrosaures sont probablement poïkilothermes, comme les crocodiles actuels. Cette différence physiologique pourrait être à l’avantage des Crocodyliformes, puisque leur physiologie plus économe nécessite des apports énergétiques moins importants et aurait donc pu leur permettre de survivre à une crise de courte durée. D’ailleurs la disparition des autres grands Reptiles marins a permis une diversification et une dispersion géographique des Dyrosaures (Jouve et coll., 2008).
Dans les milieux terrestres, la même différence au niveau de la régulation de la température corporelle pourrait, elle aussi, expliquer la disparition des Dinosaures non aviens, homéothermes, et la survie des Sébécosuchiens. La physiologie de ces derniers, testée seulement sur une seule espèce du Cénozoïque, aurait été plus proche de celle des grands lézards, c’est-à-dire poïkilotherme, et donc plus lente et économe en énergie (Cubo et coll., 2017). Reste à expliquer la survie des Dinosaures aviens, les Oiseaux, pourtant homéothermes. Leur plus petite taille et leur plumage isolant auraient-ils contribué à leur sauver la mise ?
Conclusion
La crise Crétacé-Paléogène a fait l’objet d’une attention soutenue de la part des paléontologues, et notamment des spécialistes des Dinosaures. Si de nombreuses théories ont été développées pour expliquer leur disparition, il semble que celle de l’impact météoritique tienne actuellement la corde. Les autres hypothèses, comme les épanchements volcaniques du Deccan (Inde) et les variations climatiques de la fin du Crétacé ont probablement eu un impact sur l’évolution de ces différents groupes, et fragilisé les écosystèmes à la fin du Crétacé supérieur, mais les observations faites sur les Crocodyliformes plaident plutôt pour un événement brutal et de courte durée, congruent avec l’hypothèse de l’impact météoritique comme acteur principal de la crise K-Pg.
Toutes ces hypothèses font encore l’objet d’études actuellement. Il convient donc de les considérer avec prudence, et c’est probablement le croisement des données disponibles sur l’ensemble des taxons qui permettra d’apporter des arguments solides en faveur des différentes hypothèses, et de connaître leur impact sur la biodiversité.
L’évaluation délicate de la diversité fossile et de son évolution
La diversité fossile est estimée par le nombre d’espèces présentes en un lieu et à un moment donné. Cette évaluation permet de tracer l’évolution des différents taxons au cours du temps. Loin d’un simple comptage, cette estimation est complexe et se heurte à de nombreux écueils. En effet, les fossiles proviennent des roches sédimentaires dont la distribution spatiale et temporelle est très variable, produisant des biais très importants quant à la qualité de l’enregistrement, et donc de l’estimation de la diversité fossile qui en résulte (Benton et coll., 2003). Certaines périodes ont ainsi fourni peu d’affleurements géologiques, alors qu’au contraire, d’autres ont fourni quantité de gisements d’une grande richesse fossilifère. De même, si l’exploration paléontologique est particulièrement intense dans certaines parties du monde, ce n’est pas le cas sur certains continents. Il y a donc une hétérogénéité de l’échantillonnage fossile introduisant des biais importants. La taxonomie apporte aussi son lot de biais potentiels, puisque c’est la base sur laquelle s’appuie l’évaluation de la diversité. Chez certains groupes, un grand nombre d’espèces ont été nommées au cours du temps, parfois sur des restes assez fragmentaires, et il peut s’avérer que beaucoup d’entre elles appartiennent en fait à une seule et même espèce. Cela peut provoquer la surestimation ou sous-estimation de la diversité (Jouve et coll., 2017 ; Tennant et coll., 2018). Bien sûr, les paléontologues tentent d’appliquer des corrections aux données de diversité brute, afin de minimiser le plus possible ces biais, mais il peut s’avérer que la découverte de nouvelles espèces ou la révision de groupes mettant des espèces en synonymie modifie les évaluations de diversité, et les conclusions qui en découlent.
Bibliographie
- Benton, M.J., 2003. The quality of the fossil record. Telling the evolutionary time: Molecular clocks and the fossil record 66–90.
- Cavin, L., 2002. Effects of the Cretaceous-Tertiary Boundary Event on Bony Fishes, in: Buffetaut, E., Koeberl, C. (Eds.), Geological and Biological Effects of Impact Events. Springer Berlin Heidelberg, Berlin, Heidelberg, pp. 141–158. https://doi.org/10.1007/978-3-642-59388-8_6
- Cubo, J., Köhler, M., de Buffrénil, V., 2017. Bone histology of Iberosuchus macrodon (Sebecosuchia, Crocodylomorpha). Lethaia 50, 495–503. https://doi.org/10.1111/let.12203
- Godoy, P.L., Montefeltro, F.C., Norell, M.A., Langer, M.C., 2014. An Additional Baurusuchid from the Cretaceous of Brazil with Evidence of Interspecific Predation among Crocodyliformes. PLoS ONE 9, e97138. https://doi.org/10.1371/journal.pone.0097138
- Iori, F.V., Marinho, T.D.S., Carvalho, I.D.S., Campos, A.C. de A., 2013. Taxonomic reappraisal of the sphagesaurid crocodyliform Sphagesaurus montealtensis from the Late Cretaceous Adamantina Formation of São Paulo State, Brazil. Zootaxa 3686, 183. https://doi.org/10.11646/zootaxa.3686.2.4
- Jouve, S., Iarochène, M., Bouya, B., Amaghzaz, M., 2006. A new species of Dyrosaurus (Crocodylomorpha, Dyrosauridae) from the Early Eocene of Morocco: phylogenetic implications. Zoological Journal of the Linnean Society 148, 603–656.
- Jouve, S., Bouya, B., Amaghzaz, M., 2008. A long-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Palaeocene of Morocco : Phylogenetic and Palaeobiogeographic implications. Palaeontology 51, 281–294.
- Jouve, S., Mennecart, B., Douteau, J., Jalil, N.-E., 2017. Biases in the study of relationships between biodiversity dynamics and fluctuation of environmental conditions. Palaeontologia Electronica 20, 1–21.
- Martin, J.E., Raslan-Loubatié, J., Mazin, J.-M., 2016. Cranial anatomy of Pholidosaurus purbeckensis from the Lower Cretaceous of France and its bearing on pholidosaurid affinities. Cretaceous Research 66, 43–59. https://doi.org/10.1016/j.cretres.2016.05.008
- Pol, D., Leardi, J.M., 2015. Diversity patterns of Notosuchia (Crocodyliformes, Mesoeucrocodylia) during the Cretaceous of Gondwana. Publicación Electrónica de la Asociación Paleontológica Argentina 12. https://doi.org/10.5710/PEAPA.10.06.2015.108
- Tennant, J.P., Chiarenza, A.A., Baron, M., 2018. How has our knowledge of dinosaur diversity through geologic time changed through research history? PeerJ 6, e4417. https://doi.org/10.7717/peerj.4417
- Wilberg EW, Turner AH, Brochu CA. Evolutionary structure and timing of major habitat shifts in Crocodylomorpha. Scientific Reports 9:514 doi:10.1038/s41598-018-36795-1

{kind=link}
_female_head.jpg){kind=link}
{kind=link}