

Savez vous que la banane que nous mangeons est triploïde ?
Les bananes que l'on trouve sur les étals de nos marchés ne sont guère diversifiées, pourtant les cultivars de bananiers sont très nombreux. Cet article présente leur diversité géographique et leur origine, une intéressante histoire de domestication...
Les bananiers sont cultivés dans plus de 120 pays, sur les 5 continents. En termes de production mondiale, la banane est le quatrième produit agricole (après le riz, le blé et le maïs) et le premier de la production fruitière. Dans les régions tropicales et subtropicales, les bananes sont produites à 85 % pour la consommation locale. Il existe plus de mille variétés de bananiers dont le fruit peut être consommé frais ou en tant que légume cuit, mais peut aussi faire l’objet de nombreuses transformations : chips, frites, beignets, purée, confiture, ketchup, alcool, vin, bière, etc. D’autres parties de la plante peuvent être utilisées comme fibre textile, emballage pour la cuisson, pour la construction d’abris ou la fabrication de couvertures…
L'étonnante diversité phénotypique des bananes
Bananes dessert, bananes à cuire...
Malgré 1000 à 1500 variétés existantes, les bananes proposées sur les marchés sont peu diversifiées. Leur transport délicat en est une des explications principales.
La banane Cavendish est la banane dessert la plus répandue (97 % du commerce mondial de bananes). On distingue la Grande naine (Giant Cavendish) et la petite naine (Dwarf Cavendish), le terme de naine étant relié à la taille du pseudo-tronc. Actuellement elle est cultivée aux Antilles, en Amérique centrale et aux Canaries.

Quatre variétés de banane commercialisées. De gauche à droite : Plantain, Banane rose, Frécinette et Cavendish
La banane frécinette (ou freyssinette), banane « figue sucrée », plus petite que la Cavendish, est répandue dans le monde entier, avec une présence plus forte en Asie et Afrique.
La banane rose (Antilles et Amérique centrale), une banane dessert de taille moyenne, à peau rose-rouge, possède une chair blanche très parfumée.
La banane plantain (plantain vrai, dont le centre de diversité se situe en Afrique occidentale et centrale) est très riche en amidon et doit être cuite avant consommation. On appelle aussi banane plantain d’autres variétés à cuire comme les plantains du Pacifique (par exemple la variété Popoulu), ou les Fé’i de Tahiti.
Remarque: En plus des bananes dessert et des bananes à cuire, on cultive, notamment dans la région des grands Lacs en Afrique, la « banane à bière » qui se caractérise par son amertune. Elle n’est pas exportée.
Comparaison des compositions nutritionnelles des bananes dessert et bananes plantain
|
Composition pour 100 g |
Banane dessert |
Banane plantain |
|
Eau |
74,9 |
65,3 |
|
Énergie (en kcal) |
89 |
122 |
|
Protéines |
1,1 |
1,3 |
|
Lipides |
0,3 |
0,4 |
|
Glucides |
22,8 |
31,9 |
|
Fibres |
2,6 |
2,3 |
|
Sucres |
12,2 |
15 |
La banane plantain présente une densité énergétique bien supérieure à la banane dessert ou encore aux pommes de terre (85 kcal pour 100 g). Sa teneur élevée en amidon (qui nécessite sa cuisson avant consommation) est plus importante que celle de la banane dessert et également supérieure à celle des pommes de terre (19 à 20 g pour 100 g).
Les bananiers : une grande famille
Les bananiers sont des monocotylédones qui appartiennent à l’ordre des Scitaminales ou Zingibérales et à la famille des Musaceae. Cette famille comprend essentiellement le genre Ensete (Afrique, Asie) et le genre Musa (Asie de l’Est).

Cette espèce, de grande taille (6 à 10 m de hauteur), est cultivée dans certaines régions (Éthiopie) pour sa racine comestible qui peut être consommée bouillie (les autres parties de la plante, comme le pseudo-tronc, peuvent être réduites en poudre et fermentées pour produire une farine féculente). Elle est également utilisée comme plante ornementale. Le pseudo-tronc est dépourvu de rejets à la base et l'espèce se reproduit par des graines, grosses et noires, qui germent facilement dans un terreau humide.
Le genre Ensete s’étend actuellement de l’Afrique à l’Asie du Sud et comprend 9 espèces qui ressemblent à des bananiers mais présentent de très longues feuilles aux nervures généralement cramoisies et des fruits similaires aux bananes, mais qui sont immangeables (secs et remplis de graines). L’espèce Ensete ventricosum est la plus importante dans ce genre. Elle est cultivée en Afrique. Cette plante produit un rhizome utilisé comme nourriture de base par des millions de personnes en Éthiopie.
Le genre Musa provient d’Asie et présente une forte variabilité. Il comprend un très grand nombre d’espèces, notamment celles qui produisent les bananes dessert et les bananes à cuire (dites plantain). Le genre Musa est divisé en sections, selon le nombre de chromosomes et les caractères morphologiques des espèces. Les espèces des sections Eumusa et Rhodochlamys possèdent 22 chromosomes, les espèces des sections Australimusa et Callimusa possèdent 20 chromosomes.
La majorité des bananes comestibles appartient à la section Eumusa du genre Musa et correspond à des hybrides diploïdes ou triploïdes de Musa acuminata (génome A), seul ; ou hybridé interspécifiquement avec Musa balbisiana (génome B).
Classification et répartition géographique de quelques exemples de bananiers cultivés
|
Cultivars obtenus par hybridation de Musa acuminta (genome A) et Musa balbisiana (genome B) |
||
|
DIPLOIDES |
TRIPLOIDES |
TETRAPLOIDES |
|
Groupe AA |
Groupe AAA |
Groupe AAAB |
|
Groupe AB |
Groupe AAB |
|
|
Groupe ABB |
||
Un petit groupe de cultivars incluant la banane à cuire caractéristique de Tahiti, le Fé’i, ne se trouve que dans le Pacifique et dérive d’espèces du groupe Australimusa (2n=10). Le Fe’i, Musa troglodytarum, aurait une origine complexe, impliquant au moins 3 espèces du genre Musa (M. lolodensis, M. maclayi et M. peekelii).

La domestication de la banane
La plupart des cultivars de bananes comestibles dérivent de deux espèces : l’espèce Musa balbisiana et les nombreuses sous-espèces de Musa acuminata. La figure ci-dessous présente la répartition géographique de ces différentes espèces sauvages.
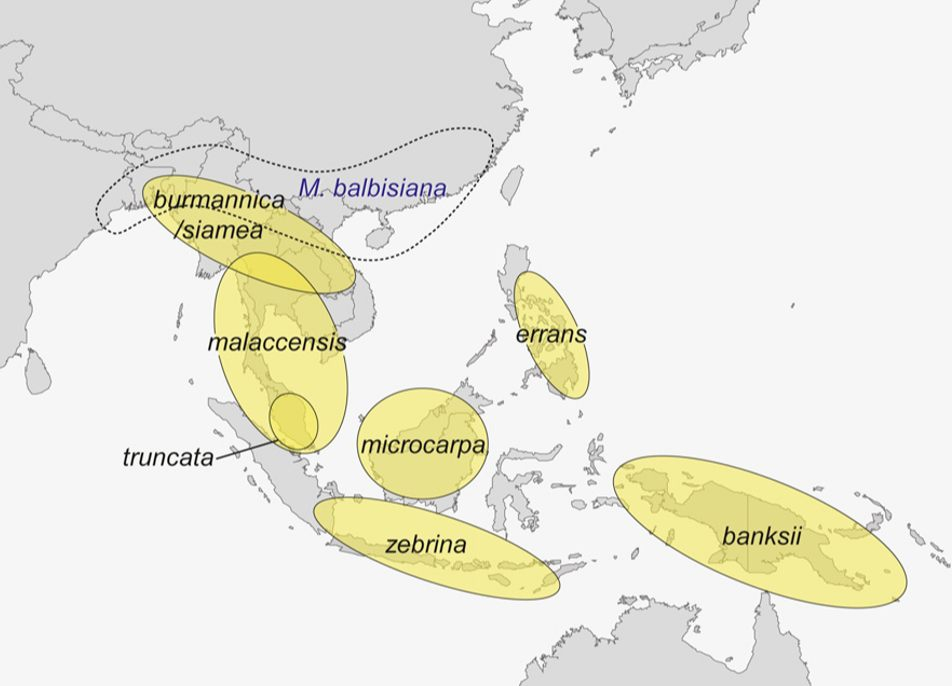
L'analyse des marqueurs nucléaires et cytoplasmiques (génome chloroplastique), ainsi que des isozymes et des anthocyanes permet de différencier plusieurs groupes (en jaune) parmi les diploïdes sauvages de Musa acuminata. Ces groupes correspondent aux sous-espèces préalablement définies sur des critères morphologiques : le groupe banksii de Nouvelle-Guinée; le groupe malaccensis de la péninsule malaisienne; les groupes burmannica et siamea qui ne peuvent être distingués par leur génome nucléaire et forment un complexe qui recouvre le Sud de la Chine, la Thaïlande, le Myanmar le Bangladesh et le Nord-Est de l'Inde (ce complexe est génétiquement proche du groupe malaccensis dont il partage une partie de l'aire de répartition); le groupe zebrina de Java; le groupe microcarpa d'Indonésie; le groupe errans des Philippines et le groupe truncata endémique des hautes terres de la péninsule malaisienne. L'espèce Musa balbisiana présente une aire de répartition plus nordique et, bien que non domestiquée, a été largement dispersée pour ses nombreux usages. On la retrouve sous forme de petites populations spontanées de la Nouvelle-Guinée à l'Est du Sri-Lanka.
La domestication des espèces sauvages s’est faite en deux périodes :
- la transition de diploïdes sauvages (avec graines dans le fruit) vers des diploïdes comestibles ;
- la transition des diploïdes comestibles vers des triploïdes comestibles.

Des diploïdes sauvages aux diploïdes comestibles
Les sous-groupes de cultivars diploïdes AA, notés AAcvs, dérivent de l’hybridation entre différentes sous-espèces de M. acuminata dans les îles de l’Asie du Sud-Est et l’Ouest de la Mélanésie, impliquant des mouvements de populations humaines durant l’Holocène (il y a environ 11000 ans). Il est concevable que ces hybridations de sous-espèces fertiles de M. acuminata géographiquement isolées soient dues à des transports par l’homme car ces plantes présentaient des utilisations nombreuses : nourriture, médecine, constructions… Il s’agissait de plants de bananes au stade de la pré-domestication (cultiwild en anglais).
L’hétérozygotie de ces hybrides AAcvs, due à des réarrangements entre chromosomes des sous-espèces parentales de Musa acuminata, a contribué à une stérilité des gamètes. Cette stérilité, associée à la sélection de la richesse du fruit en pulpe conduite par l’homme, a mené à des fruits comestibles et parthénocarpiques, dépourvus de graines.
La parthénocarpie
Le fruit se développe comme si la fleur avait été fécondée, mais les fruits ainsi produits (banane, clémentine...) ne contiennent pas de graines ou les graines ne contiennent pas d'embryons. Dans ces conditions, seule la multiplication végétative permet à la plante de se reproduire (source : Wikipédia).
Des diploïdes comestibles aux triploïdes
Des méioses anormales ont pu, dans ces cultivars AA comestibles, créer occasionnellement des gamètes diploïdes. La fusion d’un gamète diploïde avec un gamète haploïde génère un génotype triploïde stérile. Des triploïdisations spontanées, impliquant presque tous les cultivars AA ont conduit, à travers la propagation végétative de ces bananiers réalisée par l’homme, à la diversité actuelle des cultivars triploïdes, qu’il s’agisse des groupes purs AAA ou interspécifiques AAB et ABB.
Ensuite, la propagation strictement végétative de ces cultivars, par l’homme, au cours de nombreuses générations, a conduit à des variants somatiques qui ont encore augmenté la diversité phénotypique de ces espèces cultivées.
Des étude archéobotaniques ont montré que des cultiwilds dérivant de la sous-espèce Musa acuminata ssp. banksii, qui est à l’origine de nombreux cultivars actuels, existaient en Nouvelle Guinée entre 6900 et 6400 ans avant notre ère. La domestication des diploïdes parthénocarpiques a résulté de transports de ces cultiwilds et hybridation avec des sous-espèces locales, dans au moins trois régions de contact : entre la Nouvelle-Guinée et Java, au Sud, entre la Nouvelle-Guinée et les Philippines vers l’Est, et au sein des Philippines, Borneo et l’Asie du Sud-Est.
Ces cultivars diploïdes se sont ensuite peu répandus en dehors de cette première zone de diversification, à l’exception du groupe AA Mlali qui est à l’origine de la majorité des triploides cultivés actuellement (notamment la banane Cavendish). Ce sous-groupe AA pousse actuellement uniquement sur les cotes Est africaines et les îles proches (Madagascar, Zanzibar, Comores), ce qui suggère un transfert ancien à travers l’Océan Indien.
Comme le montre la figure ci-dessous, le groupe AA Mlali correspond à des hybrides de deux sous-espèces sauvages de Musa acuminata : Musa acuminata ssp. banksii (Nouvelle-Guinée) et Musa acuminata ssp. zebrina (Java).
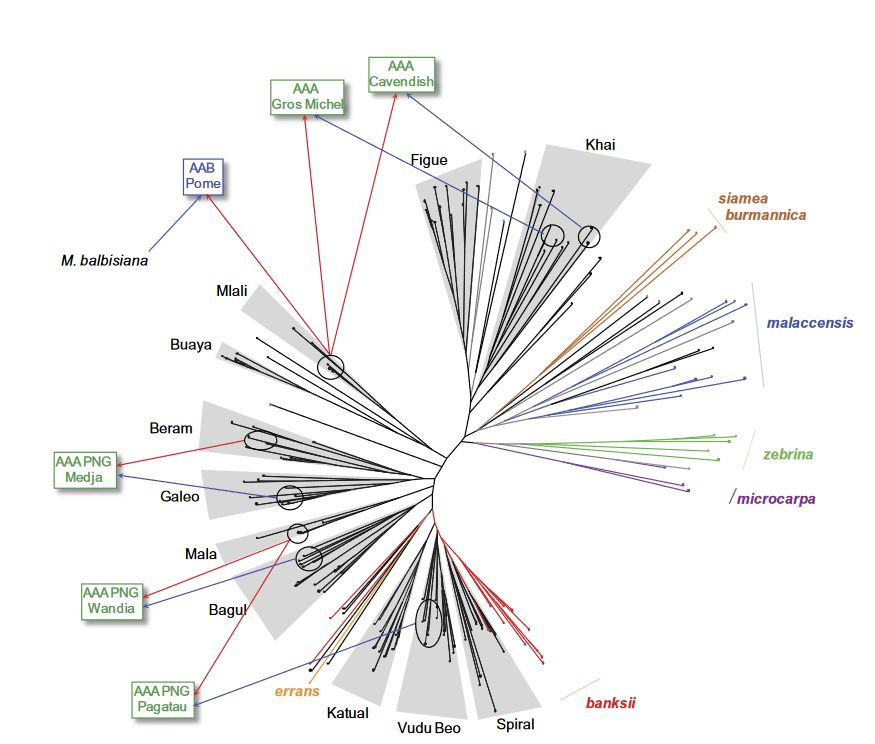
Cet arbre se base sur les différences génétiques décelées au niveau de 22 marqueurs (séquences répétitives), sur 41 diploïdes sauvages AA et 131 cultivars AA.
Les sous-espèces sauvages de Musa acuminata sont en couleur (banksii, errans, siamea, burmannica, malaccensis, zebrina et microcarpa) et les groupes de cultivars sont en noir (nommés par un représentant).
Les groupes de AAcvs, de Spiral à Beram, collectés en Nouvelle-Guinée, sont proches de la sous-espèce banksii, dont c'est la zone d'origine.
Au-delà de ces premiers groupes, la contribution du génome banksii diminue rapidement au profit d'une augmentation de celui de zebrina/microcarpa, en parallèle avec un accroissement de l'hétérozygotie.
La contribution de ces cultivars AA aux triploïdes, en tant que donneur 2n (flèches rouges) ou donneur 1n (flèches bleues), est indiquée pour quelques cultivars AAA et AAB.
La triploïdisation est apparue, ensuite, indépendamment dans diverses zones de contact entre les sous-espèces diploïdes, avec des combinaisons parentales variées. Cette triploïdisation a vraisemblablement commencé très tôt après l’émergence des cultivars AA et continue encore (on trouve de nombreux génomes AAA en Nouvelle-Guinée).
Trois de ces sous-groupes triploïdes sont cultivés actuellement très loin de leur région de naissance : les AAA Mutika Lujugira africains, les AAB plantains africains et les AAB plantains du Pacifique. Ces bananiers triploïdes ne sont pas cultivés en Asie, à l’exception de quelques plantains africains qui ont dû être introduits en Asie du Sud avec le commerce des esclaves de l’Afrique de l’Est.
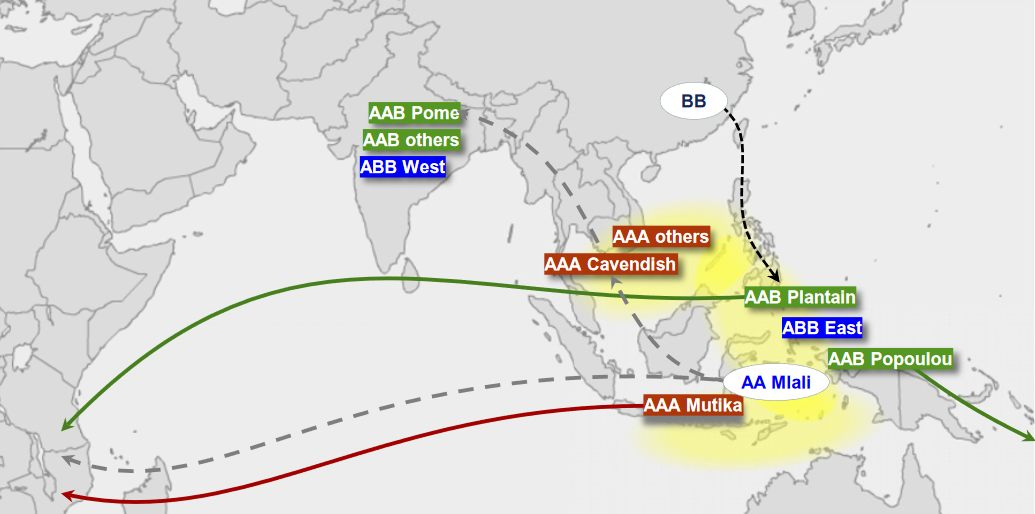
Les flèches entières indiquent les migrations préhistoriques des cultivars triploïdes vers l'Afrique et les îles du Pacifique.
Les pointillés gris indiquent la migration du groupe diploïde Mlali vers l'Asie du Sud-Est, où il a contribué génétiquement à la variété Cavendish (AAA), puis vers l'Inde où il a rencontré Musa Balbisiana (BB) et a donné la variété Pome (AAB), ainsi que vers l'Afrique.
Les pointillés noirs indiquent la migration de Musa balbisiana du Sud de la Chine vers Taïwan et les Philippines.
Les cultivars du groupe AAA sont représentés en rouge, ceux du groupe AAB en vert et ceux du groupe ABB en bleu.
En juillet 2012, après 10 ans de travail, le génome de la banane a été séquencé par le Cirad et le CEA-Genoscope de Montpellier. Il comprend 36 000 gènes. Ces données seront fort utiles pour l’amélioration des variétés, difficile chez ces espèces à faible fertilité, telle qu’une augmentation de la résistance à la fusariose (maladie de Panama due à un champignon, Fusarium oxysporum, responsable d'une grave épidémie dans les années 1950).
Références
- La banane : de son origine à sa commercialisation
http://www.pressesagro.be/base/text/v13n4/575.pdf - Multidisciplinary perspectives on banana (Musa spp.) domestication
http://www.pnas.org/content/pnas/early/2011/06/27/1102001108.full.pdf - Banana and plantain : an overview with emphasis on Pacific island cultivars
http://www.agroforestry.net/images/pdfs/Banana-plantain-overview.pdf - Composition nutritionnelle de la banane dessert
https://ndb.nal.usda.gov/ndb/foods/show/2159 - Composition nutritionnelle de la banane plantain
https://ndb.nal.usda.gov/ndb/foods/show/2351
Pour aller plus loin
- Histoire et amélioration de 50 plantes cultivées
C. Doré et F. Varoquaux, coordinateurs, INRA Collection Savoir-faire, 2006 - La légende du fé’i de Tahiti
https://www.tahitiheritage.pf/fei-tahiti-banane-plantain-montagne/ - L'espèce Ensete ventricosum en images
http://tropical.theferns.info/image.php?id=Ensete+ventricosum - Qualité de la banane plantain
http://www.cirad.fr/nos-recherches/resultats-de-recherche/2015/banane-plantain-aptitude-a-la-cuisson-et-qualite-nutritionnelle
http://www.wikiwand.com/fr/Banane_plantain - Impact fusarium
http://www.fao.org/in-action/banana-varieties-resistant-to-fungus-are-identified-using-mutation-induction/fr/ - Conférences des Nations Unies sur le commerce et le développement, 2016 : La banane (aspect commercial, différents types de maladie impactant le production) http://unctad.org/fr/PublicationsLibrary/INFOCOMM_cp01_Banana_fr.pdf


