Présentation de la structure de l'œil des Vertébrés et de sa mise en place lors du développement embryonnaire.
La structure de l’œil
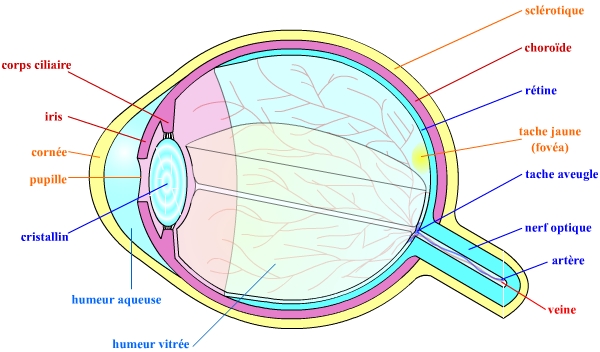
L’œil, ou globe oculaire, est une structure creuse de forme globalement sphérique. Il se compose de différentes tuniques, d’un cristallin et de liquides.
La tunique la plus externe est la sclérotique, qui est formée d’un tissu conjonctif dense et peu vascularisé et qui remplit ainsi un rôle de protection de l’œil. Du côté antérieur, cette sclérotique est remplacée par la cornée transparente. C’est cette cornée qui permet la pénétration des rayons lumineux dans le globe oculaire. Elle est riche en fibres nerveuses nociceptives : le contact avec un objet induit le clignement de l’œil et la sécrétion lacrymale (« larmes ») protégeant ainsi l’œil des contacts et des attaques bactériennes.
Plus interne que ces enveloppes essentiellement protectrices se trouvent la tunique vasculaire formée de trois parties : la choroïde, le corps ciliaire et l’iris. La choroïde est une membrane fortement vascularisée et pigmentée en brun par des mélanocytes. Le corps ciliaire est essentiellement formé de muscles lisses qui, grâce à leurs contractions, modifient la forme du cristallin et permettent ainsi l’accommodation. L’iris est la partie colorée et visible de l’œil. Composé de muscles lisses, il permet de contrôler la taille de la pupille (qui est son ouverture centrale) et donc les rayons lumineux pénétrant dans le globe oculaire : il joue un rôle de diaphragme.
La tunique la plus interne est la rétine, composée de deux couches. La couche pigmentaire (externe) empêche la lumière de diffuser dans l’œil. La couche interne est une structure nerveuse composée de nombreux photorécepteurs (cônes et bâtonnets) et de cellules traitant et acheminant l’information visuelle vers le cerveau. Cette structure fait partie du système nerveux central.

Accéder à une coupe légendée d’une tête de têtard de xénope. En cliquant sur l’œil, vous pouvez accéder à un détail sur l’œil et la tache aveugle.
Document du dossier sur le développement du xénope de Biologie et multimédia.
Les fibres nerveuses efférentes sortent de l’œil par le nerf optique. Au niveau de ce point de sortie, la rétine est tout naturellement interrompue : c’est la tache aveugle (car ne pouvant capter aucun stimulus lumineux de part l’absence de photorécepteurs). À proximité de cette tache aveugle se trouve la tache jaune (comportant une fossette centrale, la fovéa) qui est le point de la rétine avec la meilleure acuité visuelle : c’est là que les rayons lumineux arrivent directement, avec le moins d’interférences, et c’est là que la densité de photorécepteurs est la plus importante.
Le cristallin est une capsule mince et élastique composée de cellules anucléées et de fibres spécifiques. Elle joue le rôle d’une lentille biconvexe.
Le cristallin et son ligament suspenseur divisent le globe oculaire en deux chambres. La chambre antérieure est remplie d’un liquide, l’humeur aqueuse, qui est continuellement renouvelée. La chambre postérieure est remplie d’une substance gélatineuse transparente, le corps vitré ou humeur vitrée. Le corps vitré contribue à la pression intra-oculaire et ainsi à la forme de l’œil.
Origine embryonnaire de l’œil : observation
L’œil des Vertébrés dérive de formations issues de plusieurs feuillets embryonnaires :
- De l’ectoderme dérivent des tissus épidermiques (le cristallin, la cornée, l’iris), et des tissus neurodermiques (la rétine).
- Du mésoderme dérivent la sclérotique, la choroïde, ainsi que les muscles oculomoteurs (qui permettent les mouvements de l’œil).
La formation de la structure complexe qu’est l’œil est particulièrement intéressante, car elle fait appel à un grand nombre de mécanismes embryologiques, et en particulier à une cascade d’inductions. Pour des raisons expérimentales, les exemples suivants sont tirés de divers Vertébrés (xénope et souris), mais pas directement de l’homme ; ils y sont toutefois transposables.
Mise en place de l’œil : schémas
La neurulation permet la formation d’un tube nerveux dorsal chez les Vertébrés. Ce tube nerveux est à l’origine du système nerveux central. Au niveau céphalique, et plus précisément au niveau du diencéphale (voir le document sur les vésicules embryonnaires du système nerveux) on voit apparaître deux expansions latérales : ce sont les vésicules optiques. L’épiderme situé à leur contact s’épaissit, se différenciant en une placode cristallinienne, qui s’invagine en vésicule cristallinienne. Dans le même temps, la vésicule optique se replie en une cupule optique qui se referme progressivement en une sphère.
Après différenciation, la vésicule cristallinienne donne le cristallin et la cupule optique la rétine et le nerf optique. L’ectoderme qui avait recouvert la vésicule cristallinienne après son internalisation se différencie en donnant la cornée.
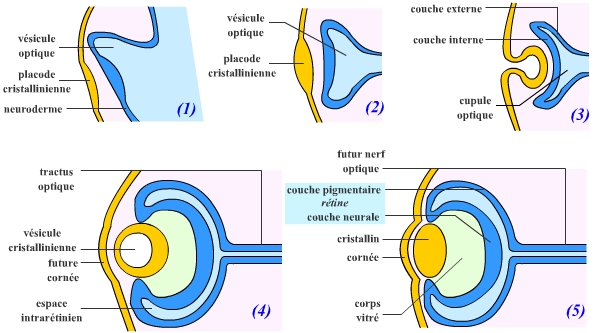
(1) Bourgeonnement de la vésicule optique à partir du diencéphale.
(2) et (3) Mise en place de la cupule optique et de la placode cristallinienne.
(4) et (5) Mise en place de la rétine et du cristallin
Documents à télécharger
Animation Flash (format swf)
Observations in utero
Une coupe transversale, réalisée au niveau céphalique d’un embryon de souris de 13 jours permet d’observer les vésicules optiques s’évaginant à partir du diencéphale. On peut noter qu’à ce stade la placode cristallinienne n’est pas encore discernable.
La coupe transversale est en fait réalisée ici sur un utérus de souris gravide. La coupe complète (à gauche sur la figure) permet d’observer l’embryon en situation dans l’utérus, et en particulier l’importante structure que forme le placenta. La deuxième structure observable dans la cavité amniotique (au-dessus de la section agrandie) correspond à une coupe transversale du même embryon, au niveau troncal : l’embryon est en effet replié sur lui-même.
De plus, le cerveau étant replié, le mésencéphale, vésicule postérieure au diencéphale apparaît aussi sur cette coupe.
Cette coupe d’un autre embryon de souris de 13 jours, permet de visualiser la structure sphérique de la vésicule optique. Elle reste toutefois toujours reliée au diencéphale.
En même temps que la structure de l’œil se met en place, les connexions nerveuses s’établissent entre la rétine et le cerveau, au cours de phénomènes faisant intervenir dans le même temps spécificité et plasticité (voir le document Spécificité des connexions rétino-tectales chez la grenouille). Pour une vue d’ensemble des voies visuelles ainsi mises en place, voir le document Voies visuelles primaires des Vertébrés.
Origine embryonnaire de l’œil : inductions
Expériences historiques : mise en évidence de phénomènes d’induction
Spemann (1901) et Lewis (1904) ont réalisé des expériences chez les Amphibiens montrant que le champ morphogénétique de la vésicule optique (c’est-à-dire le neuroderme donnant par la suite du développement la vésicule optique) est capable d’induire la formation du cristallin.

L’ablation du champ morphogénétique de la vésicule optique (donc de la rétine) conduit à deux constatations : le territoire correspondant (la rétine) ne se développe pas, mais dans le même temps les autres structures de l’œil (et en particulier le cristallin, issu de l’ectoderme – donc de tissus encore présents chez l’embryon opéré) ne se développent pas non plus. Cette expérience permet de conclure qu’un signal était émis par la vésicule optique, induisant la formation du cristallin en particulier à partir de l’ectoderme. Si cela n’était pas le cas, la rétine aurait dû se développer. Tous les embryons sont observés par leur face dorsale, pôle antérieur en haut et pôle postérieur en bas.
En effet, Spemann a réalisé l’ablation du champ morphogénétique d’une vésicule optique chez une neurula. En conséquence de cela, il observe l’absence de formation d’une vésicule optique à ce niveau, mais aussi l’absence de cristallin. Ceci montre que l’ectoderme a besoin de la présence d’une vésicule optique pour se développer en cristallin.
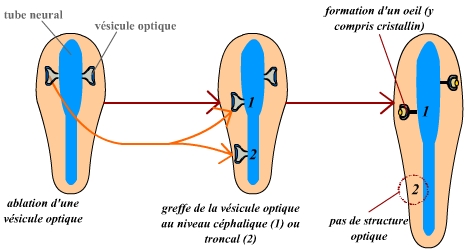
De manière similaire à ce que Spemann observe, la greffe d’une vésicule optique induit la formation des structures ectodermiques de l’œil, et ainsi aboutit à un œil complet. Ceci confirme l’induction du cristallin par la vésicule optique. Toutefois, une observation importante est réalisée : cette induction ne se produit que dans le cas d’une greffe dans la région céphalique de l’embryon receveur (par souci de simplicité, les structures optiques situées du même côté que le greffon ont ici été omises). Une greffe en région troncale n’aboutit pas à une induction. Ceci permet de conclure que l’ectoderme doit être compétent pour recevoir ce message inducteur : l’ectoderme céphalique a acquis cette compétence, et peut donc être induit, alors que l’ectoderme troncal n’est pas compétent, et ne peut donc pas être induit. Les embryons sont orientés de manière similaire à la figure précédente.
Lewis, quant à lui, a réalisé des greffes de vésicule optique. De manière intéressante, il remarque alors que la présence d’une vésicule optique induit la formation d’un cristallin au niveau de l’ectoderme sus-jacent. Ceci montre donc que la vésicule optique émet un signal vers l’ectoderme, changeant le devenir de celui-ci qui se différencie alors en cristallin.
Il est important de noter que cette expérience ne donne un tel résultat que si la greffe est réalisée au niveau céphalique. Si la vésicule optique est greffée au niveau troncal, aucune formation de cristallin n’est observée : ceci montre que le tissu ectodermique induit en cristallin doit être compétent, c’est-à-dire capable de recevoir ce message inductif.
On retrouve bien ici les différents éléments correspondant à la notion d’induction. Une induction est l’action par laquelle une cellule (ou ensemble de cellules) émet un message vers une autre cellule (ou ensemble de cellules) capable de le recevoir, ce message ayant pour conséquence de changer le devenir de la cellule (ou ensemble de cellules) réceptrice.
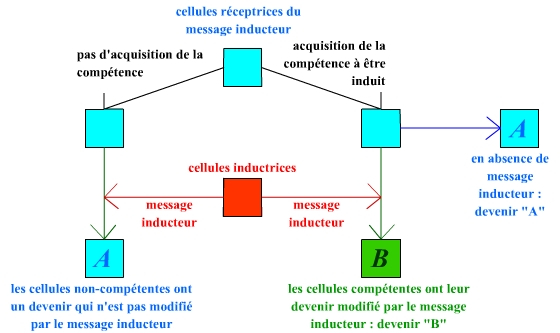

L’induction de l’ectoderme céphalique compétent par la vésicule permet sa différenciation en cristallin.
Une cascade d’inductions
Divers travaux, en particulier par Grainger dans les années 1990, ont permis de décrypter la cascade d’inductions qui a lieu lors de la formation de l’œil.
- Au stade gastrula moyenne, l’ectoderme présomptif du futur cristallin devient compétent à être induit.
- Au stade neurula et par la suite, les cellules de la plaque neurale (et des cellules de l’endoderme antérieur) puis les vésicules optiques en formation induisent l’ectoderme sus-jacent en placode cristallinienne.
- Ces placodes induisent alors la formation des cupules optiques à partir des vésicules optiques, tout en se différenciant elles-mêmes en vésicules cristalliniennes.
- Ensuite, les vésicules cristalliniennes induisent à la fois la différenciation des cupules optiques en rétines, et la formation de cornée à partir de l’ectoderme.
- Enfin, les rétines induisent la différenciation des vésicules cristalliniennes en cristallins.
On observe donc en fait un « dialogue » entre la vésicule optique et les précurseurs du cristallin, qui permettent un développement coordonné de ces deux structures et ainsi l’obtention d’une structure complexe comme celle de l’œil.
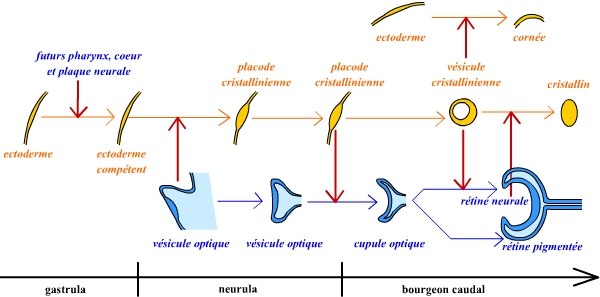
Les phénomènes d’induction sont représentés par les flèches rouges. L’ensemble de cette représentation est volontairement très schématique.
Documents à télécharger
Animation Flash (format swf)
On peut signaler que la capacité de l’ectoderme à répondre aux signaux en provenance des vésicules optiques est lié à l’expression du gène Pax6. De manière intéressante, ce gène intervient de manière importante dans la mise en place de l’œil chez les Vertébrés, mais aussi chez des organismes très différents comme les drosophiles (mouche du vinaigre).
Mise en évidence expérimentale des propriétés de l’œil
CultureSciences-Physique, le site de ressources Sciences Physiques ENS-DESCO, propose quelques expériences simples sur les propriétés de l’œil.
Au sommaire :
- Voir à l’intérieur de l’humeur vitrée
- Montrer que le cerveau inverse les images qui se forment sur la rétine
- Expérience sur la profondeur de champ de l’œil
- Largeur de la partie nette du champ de vision
- Estimation de la distance entre deux photorécepteurs
- Inhomogénéités de la rétine
- Cônes et bâtonnets
Logiciel
Un excellent complément à cette présentation et à ces expériences : le logiciel L’œil et la vision, réalisé par Pierre Perez, du lycée de Bagatelle à Saint-Gaudens. Ce logiciel (PC Windows) présente la structure de l’œil en trois dimensions, les principes de fonctionnement de l’œil et la transmission de l’information jusqu’au cerveau.
Téléchargez l'ensemble des logiciels de Pierre Perez sur le site de l'académie de Toulouse.
Remerciements
Un grand merci à l’ensemble du laboratoire Biologie et Multimédia pour leur relecture attentive. Merci tout particulièrement à Michel Delarue pour sa relecture attentive, et les photographies d’embryons présentées ici. N’hésitez pas à consulter le dossier de Biologie et Multimédia sur le développement du xénope.