Quelles différences existent entre les notions de niche écologique, de domaine vital et de territoire ? Comment expliquer les comportements territoriaux observés chez certains animaux ?
Si vous regardez un documentaire animalier sur les loups, si vous cherchez des informations sur le chant des oiseaux, si vous essayez de comprendre le comportement de votre chat ; dans tous ces cas, vous allez vous confronter au concept de « territoire ». En ouvrant une monographie sur l’écologie d’un vertébré, vous trouverez des informations sur la superficie moyenne du « domaine vital » des individus. Bien souvent par exemple, on vous dira que le domaine vital (ou le territoire) d’un mâle couvre une large surface chevauchant le domaine vital de plusieurs femelles.

Un guépard marche un peu plus vite qu’un humain (disons 10 km/h), mais peut atteindre les 100 km/h lorsqu’il pourchasse une proie [1] . Il est capable de marcher activement environ 2 h par jour. Au Botswana dans des zones cultivées, le domaine vital d’un mâle (calculé sur plus de 40 jours) est globalement circulaire et s’étend sur environ 400 km² (soit un cercle d’un rayon de 11 km). Plus de 50 % de son temps est passé dans une zone de 80 km² (cercle d’un rayon de 5 km) [2]. Pourquoi se cantonne-t-il à un territoire restreint au lieu d’aller plus loin ? Photographie d’une femelle au Kruger National Park, Afrique du Sud
Ces données sont si souvent mises en avant qu’on ne s’étonne même plus qu’un animal mobile, capable de se déplacer sur de grandes distances, reste pourtant plus ou moins sédentaire au lieu de chercher des ressources partout dans l’environnement. Pourquoi ce comportement ? Quels sont les mécanismes identifiés menant à cette structuration, quels avantages évolutifs cela confère-t-il aux individus ?
La question n’est pas neuve, elle est présente depuis les débuts de l’écologie animale moderne (début du XXe siècle) ; mais elle continue d’occuper la recherche actuelle et n’a commencé à être formalisée de façon théorique que très récemment. Nous commencerons par clarifier les terminologies et définitions, puis nous verrons les apports de la modélisation théorique sur ces questions. Des exemples empiriques seront utilisés tout le long pour illustrer les concepts et pointer leurs limites.
Niche écologique, territoire, et domaine vital : quelles différences ?
La première distinction à faire concerne le concept de niche écologique. Il s’agit pour faire simple de l’ensemble des conditions biotiques (ex : la présence de proies spécifiques) et abiotiques (ex : la température) compatibles avec la survie et la reproduction d’un organisme. Historiquement, le concept de niche s’est surtout appliqué à l’échelle d’espèces, ou du moins de populations, c’est-à-dire d’ensembles d’individus comparables. À l’inverse, on parle de territoire à l’échelle d’individus (ou de groupes dans le cas d’espèces sociales). Néanmoins cette distinction par l’échelle d’étude n’est pas suffisante et en particulier l’essor actuel des études sur la spécialisation individuelle et la personnalité animale s’accompagne de tentatives pour définir des niches écologiques à l’échelle individuelle [3].
La distinction vient du fait que la niche est un « hyper-volume » de n-dimensions non uniquement spatiales (température, pression, densité et comportement des proies, des prédateurs, etc.), tandis qu’un territoire est un espace bi- ou tri-dimensionnel (coordonnées de latitude, longitude et éventuellement d’altitude ou profondeur). Évidemment les conditions répondant à la niche écologique d’un organisme ne se rencontrent que dans certaines zones spatiales, qui limitent de fait les zones « compatibles ». Néanmoins, un individu territorial peut encore restreindre ses mouvements à seulement une petite partie de cette zone compatible, même s’il a les capacités locomotrices pour aller plus loin [4].
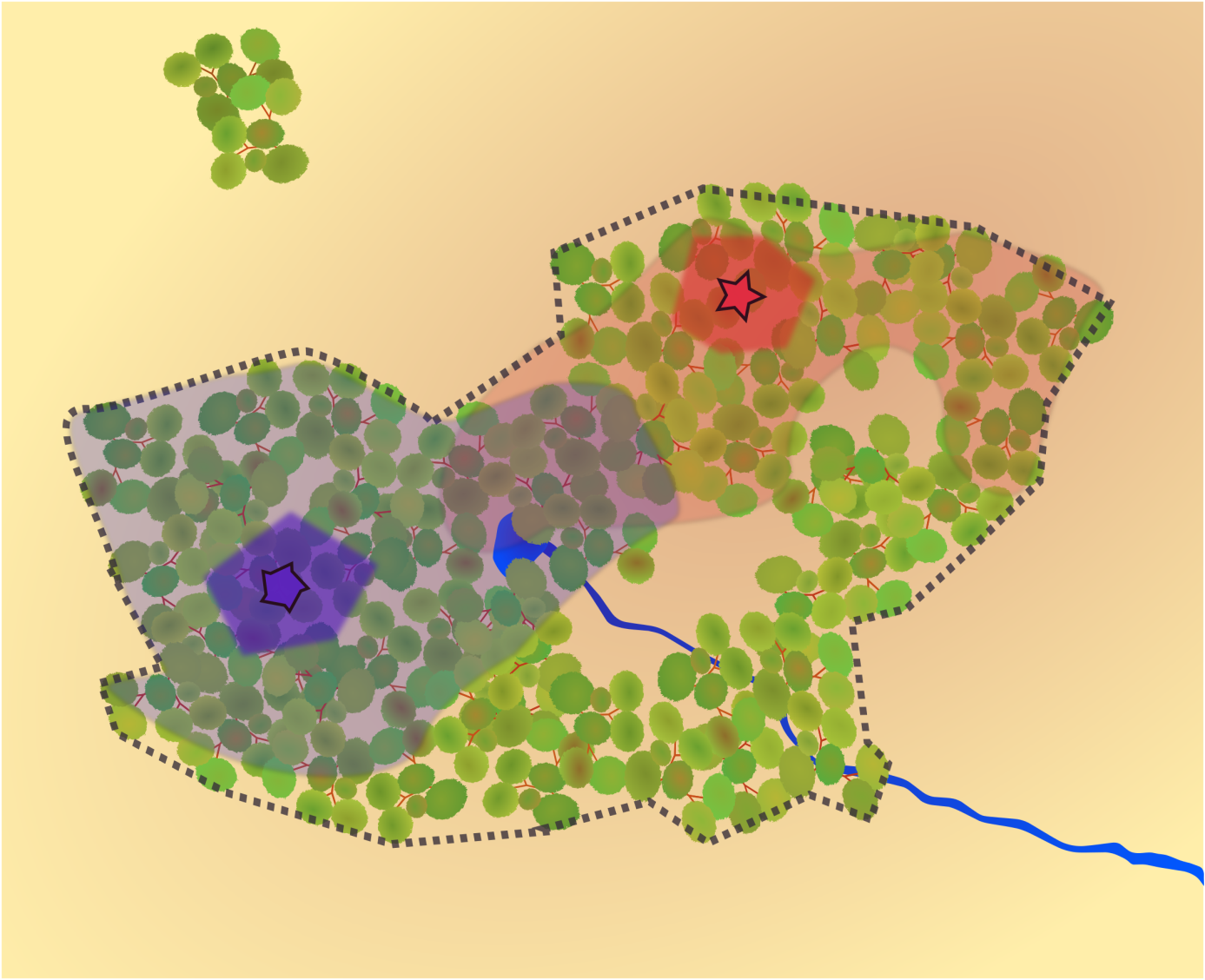
Pour une espèce forestière, on ne s’attend pas à la trouver en dehors des zones boisées de bonnes tailles. La niche écologique coïncide ainsi spatialement avec les frontières de la forêt (en pointillés). Néanmoins, toute la forêt n’est pas forcément occupée. Ici deux individus (étoile bleue et étoile rouge) occupent des domaines vitaux distincts, et une partie de la forêt est inoccupée, bien que favorable. Leurs domaines vitaux se chevauchent partiellement, en particulier autour d’un point d’eau. Ils pourront s’y rencontrer paisiblement. En revanche, au sein de chacun des domaines vitaux, une zone est âprement défendue contre tout intrus (bleu et rouge plus foncés), à proximité de la tanière (étoiles) : ce sont des territoires, au sens strict du terme. La proportion du domaine vital proprement territoriale est très variable selon les espèces et les contextes.
Il peut arriver que les limites du territoire coïncident avec les limites de niche, particulièrement si l’on tente de définir des « niches réalisées » à l’échelle d’un individu. Il devient alors difficile opérationnellement de distinguer ce qui relève d’un comportement contraint par les coordonnées spatiales d’un comportement contraint par les propriétés de niche. En d’autres termes, si un individu ne sort pas de son petit bois, est-ce parce qu’il ne survit qu’en milieu forestier, ou parce qu’il est attaché à cette zone, bois ou pas (par exemple parce qu’il ne connaît qu’un seul point d’eau et ne s’en éloigne pas) ? Toutefois la difficulté n’est qu’opérationnelle et non conceptuelle, et nous reviendrons plus en détail sur ce point au moment d’aborder les mécanismes comportementaux menant à l’émergence de territoires.
En ce qui concerne la différence entre domaine vital et territoire, elle a été posée de façon claire en 1943 [5] : le territoire est un cas particulier du concept plus général de domaine vital. Le domaine vital est la zone spatiale restreinte régulièrement utilisée par un individu dans ses activités « normales » (repos, chasse ou nourrissage, reproduction, etc.). Ainsi si vous observez un individu assez longtemps (souvent grâce à des radio-émetteurs ou des petits GPS embarqués), vous pourrez obtenir sur une carte la zone où il passe la majorité de son temps, et qui désigne donc son domaine vital. Le Dr Powell, spécialiste de l’étude du domaine vital en particulier chez les ours, a illustré ça en enregistrant ses déplacements quotidiens pendant un mois à l’aide d’un petit GPS [6] : domicile, lieu de travail, supermarché, etc. Il en a fait une analyse ici (en anglais, home range signifie domaine vital) pour bien cerner le concept, et comment scientifiquement le mesurer.

Les fous de Bassan sont très agressifs quand ils sont dans leur nid, qui correspond à leur territoire. Ils ne le sont plus du tout lorsqu’ils vont se nourrir en mer, dans une zone qui constitue leur domaine vital. Ces variations de comportement sont communes chez les animaux coloniaux. Photographie (et suivantes) prise en Écosse dans le cadre d’une étude scientifique.
Dans un territoire, l’individu défend activement ses frontières contre les intrusions, par un comportement agressif envers les étrangers, en particulier s’ils sont de la même espèce, et/ou des marquages préventifs le long des frontières (chant d’oiseau, crottes et urine, frottement contre les arbres, etc.). Notez qu’un même individu peut posséder un territoire s’inscrivant dans un domaine vital plus large, c’est-à-dire qu’il peut utiliser régulièrement des zones hors des frontières qu’il défend activement [5]. C’est le cas par exemple des oiseaux marins nichant dans les colonies : ils sont très territoriaux et agressifs à leur nid, et punissent de coups de becs les voisins trop proches. En revanche ils ne présentent plus aucun comportement territorial lors de leurs déplacements quotidiens en mer pour aller chercher de la nourriture, et partagent la zone avec de nombreux individus [7]. Cette distinction nous amène aux modélisations théoriques des mécanismes d’émergence des domaines vitaux.
Modèles théoriques pour l’émergence des domaines vitaux
De manière générale, il existe deux grandes façons de modéliser un déplacement dans l’espace : les « marches aléatoires » qui définissent des règles de vitesse et d’orientation pour chacun des « pas » successifs d’un individu ; et les modèles de diffusion qui adoptent un point de vue d’ensemble en caractérisant la probabilité au cours du temps qu’une particule s’éloigne de son point de départ.
Dans certains cas particuliers il est possible de passer d’un type de modèles à l’autre de façon analytique, c’est-à-dire par des équations mathématiques. En particulier, si les angles entre pas successifs d’une marche aléatoire sont tirées dans une loi uniforme, et que les vitesses sont tirées dans des distributions mathématiques classiques (à variance finie : loi normale, loi uniforme, loi exponentielle, etc.), alors le théorème mathématique de la limite centrale implique que cette marche aléatoire converge vers un mouvement dit brownien. Le mouvement brownien est celui utilisé classiquement par les physiciens pour décrire le mouvement erratique des particules dans un fluide. Mais ce modèle peut-il s’appliquer aux déplacements des animaux ?
Pour un mouvement brownien, on s’attend à ce que la distance au point de départ augmente comme la racine carrée du temps. Ainsi plus on attend longtemps, plus on a de chances d’observer un « animal brownien » loin de son point de départ [8] (même si l’éloignement ralentit progressivement). Cela s’oppose diamétralement au concept de domaine vital où l’on finit nécessairement par observer un plateau de distance au point de départ avec le temps : l’animal réside dans une zone particulière [4]. Les animaux territoriaux ne se déplacent donc pas suivant un mouvement brownien.
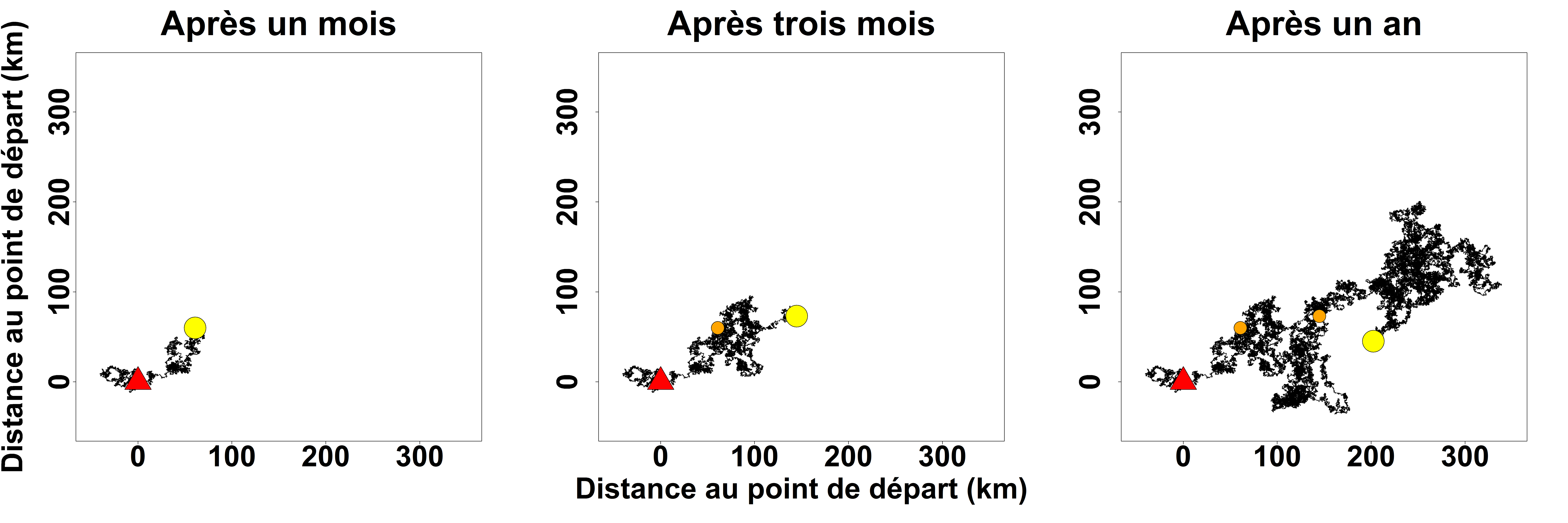
Imaginons un chien qui se déplacerait en moyenne à 5 km/h (soit à peu près la vitesse d’un humain qui marche), lâché depuis un point de départ quelconque (triangle rouge). Supposons qu’il se déplace en suivant un mouvement brownien c’est-à-dire avec peu de brusques accélérations ou ralentissements, et des directions complètement aléatoires, sans cesse changeantes. À gauche, vous avez une simulation de son trajet au bout d’un mois (le point jaune marque l’extrémité), il s’est alors éloigné d’une centaine de kilomètres de son point de départ. Même si vous laissez votre chien non attaché dans le jardin, c’est très peu probable que cela puisse arriver ! Au milieu, au bout de 3 mois, le chien (point jaune) est à 150 km (le point au bout d’un mois est toujours indiqué, en orange), et à droite après plus d’un an, il a atteint plus de 300 km, mais s’est ensuite par hasard rapproché à nouveau de son point de départ. Cependant il pourrait plus tard s’éloigner encore. On remarque que sa trajectoire est très curviligne et change constamment de direction, ce qui est là encore peu réaliste : on imagine qu’il y a en réalité plus de phases de déplacement rectiligne, qui le feraient s’éloigner encore plus vite de son point de départ si le chien était nomade et sans domaine vital !
Si maintenant on applique un modèle où les angles entre pas successifs ne sont pas aléatoires (le déplacement devient alors directif), ou si la vitesse de déplacement suit des distributions à variance infinie (mouvements dits « de Lévy »), alors la vitesse de diffusion augmente par rapport à une marche brownienne : les individus s’éloignent encore plus vite de leur point de départ [8]. Ainsi, réconcilier les observations de domaines vitaux avec des modèles de mouvement nécessite d’injecter dans ceux-ci des processus spécifiques qui permettent à l’individu de rester près de son point de départ. Ces processus peuvent se classer dans deux catégories : des points de rappels ou des frontières répulsives [4].
Avec les modèles à frontière répulsive, on retrouve le cas particulier des bordures de niche écologique, ou celui des territoires, où les marquages aux frontières et/ou l’agressivité des voisins dissuadent de trop s’aventurer chez eux. Ces modèles pourraient aussi couvrir des mécanismes de néophobie où les comportements exploratoires sont réprimés par les individus et les zones inconnues évitées, même en l’absence de voisin, mais c’est un processus très difficile à mettre en évidence empiriquement.

Ces mouvements de tête de bas en haut, les ailes mi-ouvertes, sont utilisés par les adultes au début de la période de nidification pour revendiquer leur zone de nid auprès des voisins. Ces rituels démonstratifs, comme les chants d’oiseaux ou le dépôt d’urine et de crottes chez les carnivores, servent à communiquer sur la présence et l’emplacement des frontières territoriales à ne pas dépasser. Cela évite le plus souvent les coûts liés aux affrontements directs.
Par opposition, le fait de rester dans un domaine restreint pourrait s’expliquer par le fait que cette zone contient des points d’attraction. Cela peut être le cas d’individus ne s’éloignant guère de leur nid (ou « place centrale »), mais on peut imaginer des cas plus complexes où les individus préfèrent exploiter des zones ou ressources avec lesquels ils sont familiers [9]. Il est raisonnable de penser que cela implique l’usage de mémoire spatiale, qui permet des retours à des points déjà visités par le passé, surtout si le domaine vital est bien plus vaste que les capacités de perception directe de l’individu. Insérer des règles de mémoire à court terme dans les modèles de marche aléatoire est alors suffisant pour voir émerger et se maintenir un domaine vital, la mémoire étant remise à niveau à chaque nouvelle visite récompensée de ressources [7,10].
Par ailleurs, un résultat remarquable des modélisations théoriques est que si l’on lâche plusieurs individus ensemble au centre d’un même environnement, et que ces individus n’ont aucune interaction directe entre eux quand ils se croisent (en particulier, pas de territorialité), il est possible de voir émerger des domaines vitaux qui se chevauchent très peu entre eux [11]. Cela fonctionne si les ressources sont agrégées de façon hétérogène dans l’environnement, et si les individus suivent deux règles : premièrement utiliser une mémoire à court terme des zones de ressources rencontrées et de la quantité de ressources qu’on peut y trouver, pour éventuellement y retourner plus tard, et deuxièmement oublier progressivement les zones de ressources non-revisitées.
Ainsi l’on voit que des domaines vitaux ségrégeant peuvent effectivement très bien émerger sans besoin de défense active des frontières ni même d’interactions directes entre voisins. La raison est qu’en mémorisant les emplacements des ressources locales, chaque individu augmente ses chances d’y retourner avant que les ressources soient dévorées par un voisin ; ainsi chacun devient familier avec son environnement local et donc plus compétitif que ses voisins en son sein. Il n’y a ainsi plus de raisons valables pour aller explorer les zones des voisins, du moins tant que l’environnement et les ressources ne changent pas. L’arrivée d’un nouveau voisin en particulier peut souvent redéfinir les domaines vitaux [11].

Les troupes de babouins adoptent des domaines vitaux souvent stables sur de longues périodes, et qui se chevauchent partiellement entre troupes voisines. Hormis dans de très rares cas de troupes vivant dans des environnements particuliers, ils ne sont pas territoriaux12,13. Leur fidélité spatiale est donc probablement le signe de leur mémoire. Photo prise dans le cadre du Tsaobis Baboon Project en Namibie (Zoological Society of London).
La territorialité permet néanmoins de bien mieux monopoliser et de sécuriser un ensemble de ressources, et ainsi de s’octroyer un important avantage adaptatif par rapport à des individus sans territoire [14] . Cependant c’est un comportement coûteux : il faut patrouiller ses frontières, laisser des marques lors de son passage (crottes, griffures, odeurs, etc.), et éventuellement risquer des blessures en cas de conflit direct. La territorialité s’exprime donc lorsqu’il est possible pour l’animal d’endurer ces coûts : une ressource plus facile à défendre (par exemple très localisée, comme une tanière), et/ou dont la perte, même momentanée, serait plus coûteuse que sa défense (comme la perte d’un abri pour sa progéniture). Ainsi on observe souvent que la territorialité est saisonnière, et maximale pendant la saison de reproduction (pensez au chant des oiseaux).
En guise de conclusion
Tout l’enjeu des études empiriques consiste alors à déterminer quels mécanismes prévalent selon les espèces ou les contextes écologiques. Cette question, pour pouvoir dépasser la valeur d’une pure accumulation de faits, peut s’aborder sous l’angle de la sélection naturelle et des avantages que l’adoption d’un domaine vital ou d’un territoire confère aux individus. De façon générale, domaines vitaux et territoires présentent pour les individus les adoptant l’avantage de considérablement réduire la compétition avec les voisins [15]. Tant que ces avantages sont supérieurs aux coûts du comportement (comme ceux de l’entretien du cerveau pour la mémoire, ou ceux liés aux rituels et agressions pour la territorialité), on s’attend à observer le comportement.
Néanmoins l’estimation exacte des coûts et bénéfices est complexe et peut dépendre d’une multitude de facteurs [12] : abondance et vitesse de renouvellement des ressources, dominance des individus et propension des ressources à être monopolisables, densité de la population, taille et besoins des individus en particulier selon les sexes ou les périodes du cycle de vie, espérance de vie et fréquence de reproduction de l’espèce en question, etc.
On n’a donc pas fini d’étudier les cas empiriques ! Cette façon d’aborder l’étude du comportement, en analyse coûts/bénéfices, s’inscrit dans la théorie de la sélection naturelle et s’inspire de modèles développées initialement par des économistes. Elle permet souvent de rapprocher des observations qui semblaient sans rapport entre elles. Par exemple, pour quitter les animaux mais rester en écologie, vous pouvez aller consulter l’article La géopolitique des champignons du Saule Causeur, qui aborde ces questions territoriales sous un angle un peu différent !
Quelques références
- Wilson, A. M. et al. Locomotion dynamics of hunting in wild cheetahs. Nature 498, 185–189 (2013).
- Houser, A., Somers, M. J. & Boast, L. K. Home range use of free-ranging cheetah on farm and conservation land in Botswana. South Afr. J. Wildl. Res. 39, 11–22 (2009).
- Ingram, T., Costa-Pereira, R. & Araújo, M. S. The dimensionality of individual niche variation. Ecology in press, (2017).
- Börger, L., Dalziel, B. D. & Fryxell, J. M. Are there general mechanisms of animal home range behaviour? A review and prospects for future research. Ecol. Lett. 11, 637–650 (2008).
- Burt, W. H. Territoriality and Home Range Concepts as Applied to Mammals. J. Mammal. 24, 346–352 (1943).
- Powell, R. A. & Mitchell, M. S. What is a home range? J. Mammal. 93, 948–958 (2012).
- Wakefield, E. D. et al. Space Partitioning Without Territoriality in Gannets. Science 341, 68–70 (2013).
- Benhamou, S. On the expected net displacement of animals’ random movements. Ecol. Model. 171, 207–208 (2004).
- Piper, W. H. Making habitat selection more “familiar”: a review. Behav. Ecol. Sociobiol. 65, 1329–1351 (2011).
- Van Moorter, B. et al. Memory keeps you at home: a mechanistic model for home range emergence. Oikos 118, 641–652 (2009).
- Riotte-Lambert, L., Benhamou, S. & Chamaillé-Jammes, S. How memory-based movement leads to nonterritorial spatial segregation. Am. Nat. 185, E103-16 (2015).
- Hamilton, W. J., Buskirk, R. E. & Buskirk, W. H. Defense of Space and Resources by Chacma (Papio Ursinus) Baboon Troops in an African Desert and Swamp. Ecology 57, 1264–1272 (1976).
- Markham, A. C., Guttal, V., Alberts, S. & Altmann, J. When good neighbors don’t need fences: temporal landscape partitioning among baboon social groups. Behav. Ecol. Sociobiol. 67, 875–884 (2013).
- Maher, C. R. & Lott, D. F. A review of ecological determinants of territoriality within vertebrate species. Am. Midl. Nat. 143, 1–29 (2000).
- Riotte-Lambert, L. Se souvenir et revenir: Approche théorique et méthodologique des stratégies de déplacement récursif et de leurs conséquences populationnelles. (Université de Montpellier, 2016).