

Cet article présente l’algue Volvox, être vivant formé d’une colonie de cellules chlorophylliennes, situé à la frontière entre unicellulaire et pluricellulaire. Il explique ensuite les deux mécanismes de reproduction du genre Volvox et précise en quoi le mode de formation des colonies filles (par une sorte de gastrulation) présente un intérêt biologique majeur.
Volvox, une algue verte coloniale
Une algue d’eau douce composée de plusieurs cellules
Le genre Volvox regroupe des algues vertes appartenant aux Chlorobiontes, ensemble qui comprend, en plus des Algues vertes, la plupart des plantes terrestres (Embryophytes).
Algues d’eau douce (voir ci-dessous), elles forment des colonies sphériques de plusieurs centaines à plusieurs milliers de cellules, qui sont individuellement analogues à Chlamydomonas sp.
On trouve cependant deux types cellulaires haploïdes différents au sein de ces colonies : des cellules somatiques et des cellules reproductrices, un peu plus grosses, appelées gonidies.
Le thalle qu’elles forment est un archéthalle, la catégorie de thalle dont la structure est la plus simple. Chez Volvox, cet archéthalle est un cénobe (ou cœnobe), ce qui signifie littéralement « vivre ensemble ».

Toutes les photos de cet article ont été réalisées à partir de prélèvements effectués dans des mares similaires à celle présentée sur la photo ci-dessus. Ces prélèvements ont été réalisés en Loire-Atlantique au mois d’août.
Toutes les cellules sont situées à la périphérie de la sphère, le centre étant constitué d’un gel mucilagineux produit par les cellules elles-mêmes. Les gonidies, bien que situées en périphérie comme les cellules somatiques, sont légèrement en retrait, décalées vers l’intérieur de la sphère. Les cellules résultant des divisions de ces gonidies s’accumulent à l’intérieur de la sphère (voir figure ci-dessous).
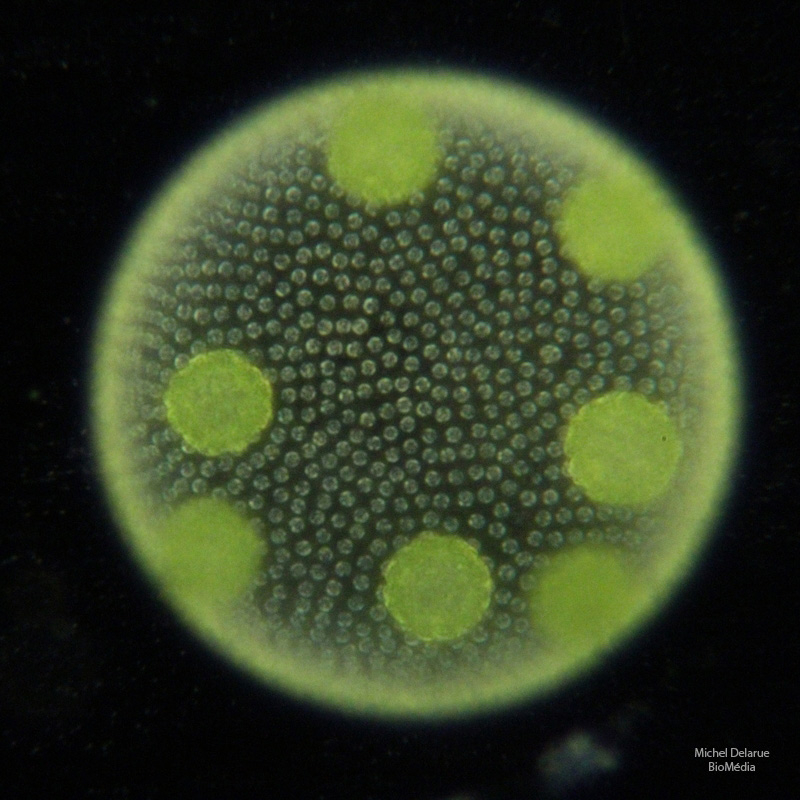
Les cellules somatiques apparaissent non jointives, d’une couleur vert pâle. Elles forment un unique feuillet situé à la périphérie de la colonie. Elles reposent sur un « gel » qu’elles secrètent et qui remplit l’intérieur de la sphère. On peut observer, sur cette photo, sept massifs de cellules d’un vert plus soutenu : il s’agit de jeunes colonies filles, fruit d’une reproduction asexuée (voir plus loin).
Les cellules somatiques
Les cellules somatiques sont situées à la périphérie de la sphère de la colonie. Espacées les unes des autres, elles sont reliées par des ponts cytoplasmiques visibles sur la figure ci-dessous.
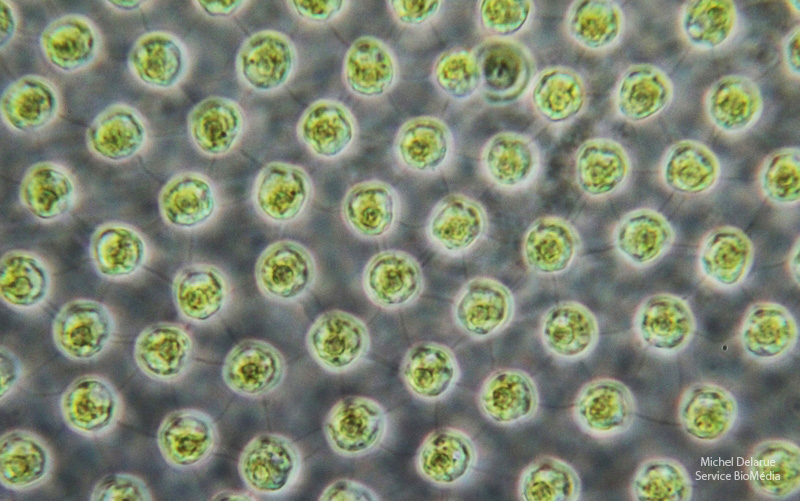
On constate que les cellules somatiques de Volvox sont chlorophylliennes (comme en atteste leur couleur verte) et qu’elles sont toutes reliées les unes aux autres par des ponts cytoplasmiques. Ceux-ci apparaissent comme des filaments noirs allant d’une cellule à une autre.
Les Volvox sont considérés comme un exemple très intéressant d’être vivant situé à la frontière entre l’unicellulaire et le pluricellulaire. En effet, si les cellules somatiques sont rassemblées, elles ne sont pas différenciées. D’ailleurs, chaque cellule de Volvox est très proche de Chlamidomonas sp. (les deux appartenant à l’ordre des Volvocales). C’est pourquoi on parle de « colonie » et non « d’organisme ».
Cela dit, il est important de noter que la forme coloniale des Volvox n’est pas une survivance d’une forme primitive de pluricellularité, qui serait à l’origine des algues et des végétaux pluricellulaires actuels.
En effet, la transition entre l’ancêtre unicellulaire de Volvox et la forme coloniale, telle que nous la connaissons aujourd’hui, a commencé il y a environ 200 millions d’années, et a duré approximativement 35 millions d’années. Cela correspond à la fin du Trias et à une partie du Jurassique, c’est-à-dire à une époque où les végétaux pluricellulaires étaient largement répandus, et ce depuis longtemps.
Pour mémoire, au Jurassique, la flore se caractérise par le développement de jungles, dominées par les conifères, grâce à un climat chaud et humide. Et plus de 100 millions d’années auparavant, le Carbonifère (− 360 à − 300 millions d’années) était déjà caractérisé par la présence de forêts comportant des végétaux de taille exceptionnelle (en particulier des Cryptogames, comme les Fougères, et des Gymnospermes, comme les Conifères).
Les cellules reproductrices
Les cellules responsables de la multiplication végétative des Volvox sont les mêmes qui, sous certaines conditions, donneront les zoïdes de la reproduction sexuée. On appelle indifféremment gonidies ce groupe de cellules responsables de la multiplication végétative ou de la reproduction sexuée.
Situées au départ à la périphérie de la colonie, légèrement en retrait des cellules somatiques, elles vont, en fonction des conditions environnementales, soit se diviser activement pour former des colonies filles (reproduction asexuée), soit être à l’origine de la formation de gamètes mâles ou femelles (reproduction sexuée).
Mécanismes de reproduction chez Volvox
Reproduction asexuée
La reproduction asexuée assure la multiplication des colonies. Tout commence lorsque les gonidies d’une colonie deviennent matures (ce qui caractérise le passage du stade « colonie juvénile » à « colonie adulte »). Ces cellules entrent alors dans une phase de multiplication rapide par mitose (11 à 12 cycles au total). Les 5 premières divisions sont équilibrées, donnant naissance à 32 cellules identiques. Lors de la 6e division, des mitoses déséquilibrées donnent naissance à 16 cellules plus grosses que les autres : les futures gonidies. Les divisions continuent alors pour les petites cellules, donnant naissance à une jeune colonie de 2000 à 4000 cellules somatiques et 16 gonidies.
La taille des colonies filles, ainsi que de la colonie mère, va alors augmenter sans changement du nombre de cellules, par production et dépôt de mucilage par les cellules somatiques. Enfin, les colonies filles vont sortir par éclatement de la colonie mère, dont les cellules somatiques vont entrer en dégénérescence. Les cellules somatiques sont donc des cellules mortelles, alors que les gonidies sont potentiellement immortelles.
La phase de multiplication dure environ 8 heures, et la durée totale d’un cycle est d’environ 2 jours.

On voit la formation de colonies filles par division active des cellules reproductrices. Notons que toutes les cellules sont haploïdes, depuis les gonidies de départ jusqu’aux cellules des colonies filles, en passant pas les cellules somatiques de la colonie mère.
On peut noter sur la photo de droite que la colonie fille en cours de formation est une sphère creuse.
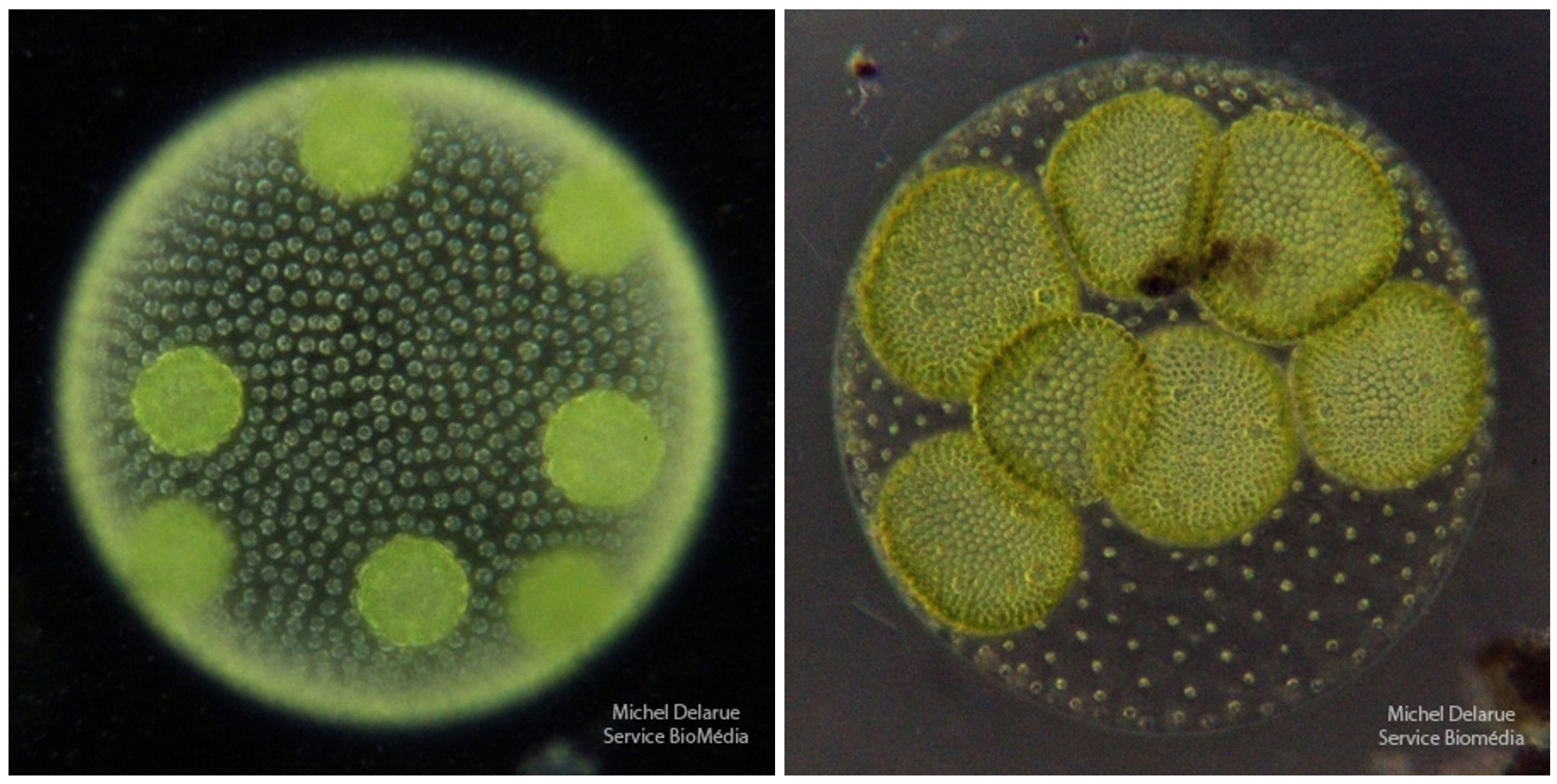
On constate que les cellules somatiques de la colonie mère sont beaucoup plus espacées sur la photo de droite, comparé à la situation observée sur la photo de gauche. Cela correspond à la phase de croissance de la taille de la colonie sans multiplication supplémentaire, phase qui précède l’éclatement de la colonie mère et la libération des colonies filles.
Le phénomène peut-être le plus marquant et le plus intéressant de cette séquence concerne un événement qui survient après la phase de multiplication.
Les cellules somatiques sont polarisées, avec leurs deux flagelles situés du côté apical, donc tournés vers l’extérieur de la sphère (d’où la capacité de la colonie à se déplacer), et les gonidies, pour leur part, sont légèrement décalées vers l’intérieur. Or, la multiplication de chaque gonidie aboutit à une sphère dans laquelle tous les flagelles des nouvelles cellules somatiques sont tournés vers l’intérieur, et dont les nouvelles gonidies dépassent légèrement à l’extérieur du feuillet des cellules somatiques.
Cette disposition est exactement à l’inverse de celle observée dans une colonie adulte. Le passage de l’une à l’autre est assuré par un mouvement du feuillet cellulaire des jeunes colonies, qui va se retourner jusqu’à inverser la configuration initiale (voir figure ci-dessous). Ce mouvement n’est pas sans rappeler ce que l’on observe au moment de la gastrulation chez les embryons des spongiaires, ce qui a contribué à faire des Volvox un objet d’étude particulièrement prisé.
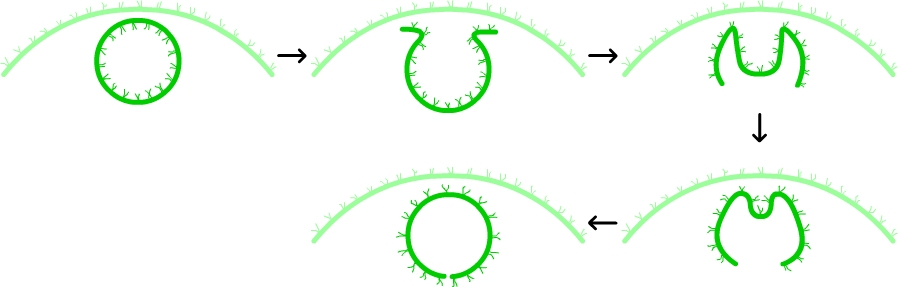
La multiplication active d’une gonidie a donné naissance à une jeune colonie sphérique dont l’orientation des cellules (pôle apical, caractérisé par la présence des flagelles, tourné vers l’intérieur de la colonie) est à l’inverse de ce qui est observé chez la colonie mère (pôle apical tourné vers l’extérieur). Lors de la formation d’une colonie fille, après la phase de multiplication, on assiste à un mouvement du feuillet cellulaire qui va se retourner et se refermer, pour reformer une sphère dans laquelle l’orientation des cellules a été inversée. Ce mouvement ressemble à la gastrulation observée dans le développement embryonnaire des Spongiaires.
Reproduction sexuée
Le mode de reproduction sexué est très largement minoritaire. En fait, la reproduction sexuée n’intervient qu’en cas de survenue de conditions difficiles, car elle entraîne la production d’un œuf très résistant permettant d’attendre le retour de conditions meilleures.
La reproduction sexuée se fait par oogamie, c’est-à-dire par la rencontre entre un petit gamète mobile (produit en grand nombre) et un gros gamète immobile chargé de réserve (produit en petit nombre).
Sous l’influence de facteurs déclenchant, le développement des colonies juvéniles va être orienté vers une reproduction sexuée. On observe alors une différence de comportement entre les colonies mâles et les colonies femelles.
Dans les colonies femelles, les gonidies vont se diviser 6 fois pour donner naissance à 64 cellules identiques. À la 7e division, 32 gonidies vont être produites, les 96 autres cellules continuant à se diviser jusqu’à donner 2000 à 4000 cellules somatiques. Mais à l’inverse de ce que l’on observe dans la reproduction asexuée, les gonidies ne vont pas entrer dans une phase de multiplication. Elles vont donner naissance à des gamètes femelles immobiles.
Dans les colonies mâles, les gonidies vont se diviser 7 fois pour donner naissance à 128 cellules identiques. À la 8e division, 128 gonidies vont être produites, les 128 autres étant les cellules somatiques qui ne se diviseront plus, contrairement à ce qui se passe dans les autres cas de figure. En revanche, chaque gonidie va se diviser pour donner naissance à un massif compact de 64 à 128 gamètes mobiles, les gamètes mâles (voir figure ci-dessous).
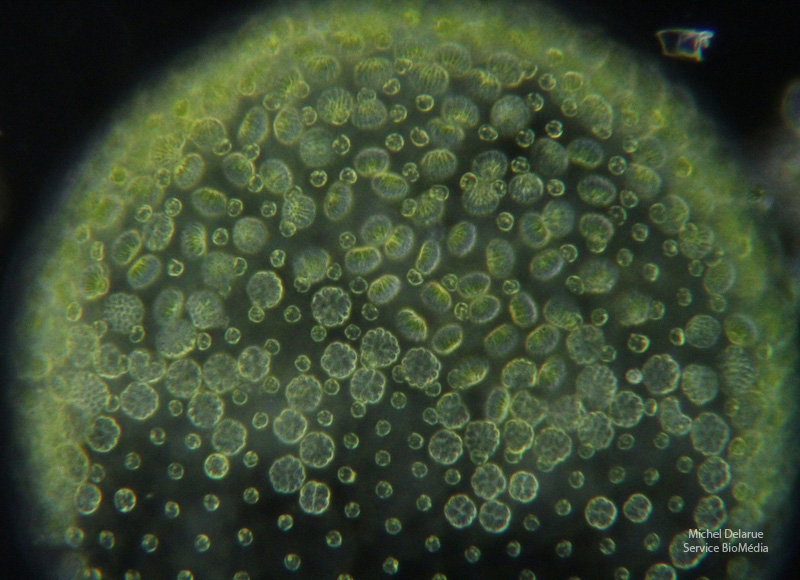
Sous l’action de conditions défavorables, le développement des futures gonidies des colonies juvéniles a été orienté vers une reproduction sexuée. Une fois ces colonies matures, chaque gonidie se divise pour donner naissance à un massif de cellules germinales mobiles (nombreux groupes cellulaires observés sur la photo). Ces gamètes seront libérés en présence de colonies femelles pour permettre une fécondation par oogamie.
En présence de colonies femelles, les gamètes mâles vont être libérés et pouvoir se déplacer. La rencontre avec un gamète femelle (qui est resté dans la colonie dont il est issu) va donner naissance à un œuf. Celui-ci va s’entourer d’une paroi qui lui permet d’être une forme de résistance. Lorsque les conditions le permettent, cet œuf va entrer en méiose, ce qui va aboutir à la formation d’une unique zoospore haploïde (cellule mobile). Cette dernière va pouvoir se diviser pour donner naissance à une nouvelle colonie.



