

À Londres, les populations de moustiques aériennes et celles du métro semblent ne pas se reproduire. Peut-on pour autant dire que l'on a affaire à deux espèces différentes ?
Introduction
Depuis sa publication en 1999, les résultats de Byrne et Nichols sur les populations de moustiques de Londres ont été abondamment relayés. Pourtant, certains résultats de cette étude ont été mal compris, et de nouvelles découvertes ont été réalisées depuis. Nous nous proposons d’apporter quelques éclaircissements sur ce cas d’école.
Katharine Byrne et Richard A. Nichols ont publié en 1999 une étude sur les différences existant entre les populations de Culex pipiens vivant dans les tunnels et stations du métro de Londres d’une part, et celles de surface d’autre part. Ces dernières (forme pipiens) se reproduisent exclusivement en milieu ouvert, réalisent une diapause hivernale (phase de vie ralentie), piquent essentiellement les oiseaux et les femelles pondent uniquement après un repas de sang ; les moustiques du métro (forme molestus) présentent des caractéristiques opposées.
Les formes molestus et pipiens ne sont pas spécifiques à la ville de Londres mais se retrouvent en Europe, en Afrique du Nord, au Moyen-Orient, aux États-Unis et plus généralement dans toutes les zones tempérées du globe.

Les formes pipiens et molestus de Culex pipiens sont morphologiquement indistinguables.
|
|
Forme molestus |
Forme pipiens |
|
Lieu d’accouplement |
Souterrain |
En surface |
|
Accouplement |
Dans des espaces confinés |
En dehors d’espace confinés |
|
Principale source de nourriture |
Mammifères |
Oiseaux |
|
Production d’œufs |
Pas de nécessité d’un repas de sang avant la première ponte |
Nécessité d’un repas de sang avant la première ponte |
|
Cycle de vie |
Pas de diapause hivernale |
Diapause hivernale |
Malgré des caractéristiques morphologiques identiques, les formes pipiens et molestus présentent donc d’importantes différences physiologiques et écologiques. Faut-il alors considérer que ces deux formes font partie de la même espèce ou bien qu’elles constituent deux espèces différentes ?
Les moustiques de Londres
Pour le savoir, Byrne et Nichols ont collecté, à différents endroits de Londres, des populations de moustiques, en milieu aérien ou souterrain. Ils ont testé chaque population pour savoir s’il s’agissait de formes molestus ou de formes pipiens. Les populations pouvant se reproduire dans l’espace confiné du laboratoire ont été identifiées comme des molestus ; les autres comme des pipiens.
Les croisements entre des populations de molestus issues de différentes stations de métro ont tous donnés une génération F1 et une génération F2, montrant ainsi que les populations souterraines de moustiques font bien partie de la même espèce.
Les croisements entre des mâles pipiens et des femelles molestus n’ont pas donné de descendance, sauf dans un seul cas, dont la F1 n’était pas fertile. Ce résultat ne signifie pas à lui seul que les formes pipiens et molestus de Londres font partie de deux espèces différentes, mais seulement que les mâles pipiens ne peuvent se reproduire en milieu confiné. Comme c’est précisément ce critère qui avait été utilisé pour les identifier comme pipiens, ce résultat est cohérent.
Après leurs études sur la reproduction en laboratoire, Byrne et Nichols ont cherché à quantifier la diversité génétique des différentes populations de moustiques en s’intéressant à 20 allozymes (ou isoenzymes : enzymes issues d’allèles différents d’un même gène). Ils ont ainsi pu montrer que le polymorphisme enzymatique des populations de surface est bien plus important que celui des populations souterraines, qui ne possèdent qu’une fraction des allèles des formes pipiens. L’hétérozygotie globale des populations pour les locus étudiés a été déterminée : Hsurface = 0,27 et Hmétro = 0,07.
Les auteurs ont également montré que les populations de surface, même géographiquement éloignées, étaient génétiquement assez homogènes ; idem pour les populations souterraines. Par contre, les moustiques de surface et ceux du métro montrent des différences génétiques importantes, même lorsqu’il s’agit de populations prélevées dans une même zone géographique et distantes de quelques centaines de mètres seulement. Ces résultats montrent que le flux de gènes entre les populations de surface et les populations souterraines est très limité, voire absent. Pour autant, et contrairement à ce que laissent penser des synthèses publiées par la suite (Chanet et al.; Lecointre et al., 2009), Byrne et Nichols ne vont pas jusqu’à distinguer les pipiens et les molestus comme des espèces différentes :
« In this study no assumptions have been made about the taxonomic status of the two types of Culex. »
« Dans cette étude, nous n’avons fait aucune supposition sur le statut taxonomique des deux types de Culex. »
Byrne et Nichols insistent seulement sur le fait qu’il existe un isolement reproducteur fort entre les deux populations de moustiques. Celui-ci viendrait des pressions de sélection différentes s’exerçant en surface et dans le métro, et d’une dérive génétique importante dans les petites populations de molestus, menant à une différenciation génétique des deux formes de moustiques. Le flux de gènes est également limité par les différences physiologiques existant entre les deux formes : les moustiques de surface qui passeraient dans le métro ne pourraient se reproduire dans un espace confiné ; les moustiques du métro qui arriveraient en surface ne passeraient pas l’hiver (car ils ne peuvent entrer en diapause) limitant ainsi la période durant laquelle ils seraient capables de se reproduire.
Comme les populations du métro sont génétiquement homogènes, même celles situées à de longues distances, cela suggère qu’il n’y a eu qu’un seul (ou tout du moins un nombre limité) événement de colonisation du métro depuis des populations de surface. Dans leur étude, Byrne et Nichols, en se basant sur des données génétiques, soutiennent l’hypothèse d’une colonisation du métro par une population de C. pipiens de Londres, plutôt qu’une colonisation par des C. molestus présents plus au sud de l’Europe.
D’où viennent les molestus du métro de Londres ?
Cependant, une autre étude publiée cinq ans plus tard soutient plutôt la seconde hypothèse. Dans celle-ci, Dina M. Fonseca et ses collaborateurs ont analysé huit séquences microsatellites de 33 populations de moustiques venant de différents pays. Leurs résultats montrent que les moustiques souterrains de différentes régions du monde sont plus proches entre eux que des populations de surface et vice-versa. Par ailleurs les moustiques souterrains (vivant dans des tunnels de métro, des fosses septiques, des égouts…) du nord de l’Europe ont plus d’allèles en commun avec les populations du nord de l’Afrique et du Moyen-Orient, qu’avec les populations aériennes nord-européennes. Ceci laisse donc penser que les moustiques du métro londonien proviendraient de populations de molestus venant d’Europe méditerranéenne et non de pipiens londoniens.
Les différences génétiques entre les formes molestus et les formes pipiens sont considérées par les auteurs comme suffisantes pour distinguer deux espèces : Culex pipiens et Culex molestus qui auraient divergé il y a environ 10 000 ans.
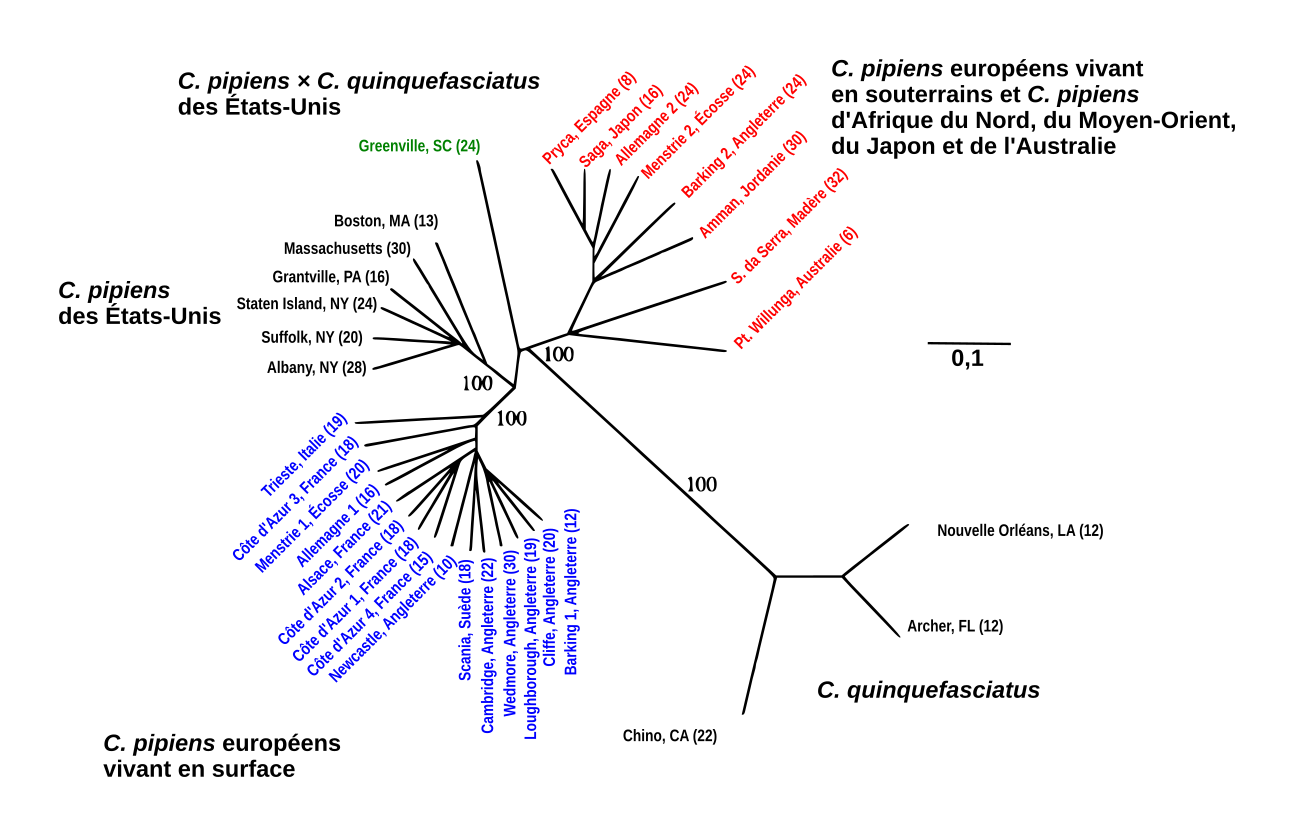
Arbre de distance non raciné, basé sur la proportion d’allèles partagés et représentant les relations entre les 33 populations étudiées par Fonseca et al. Les nombres entre parenthèses correspondent aux nombres d’individus échantillonnées et ceux sur les branches aux valeurs de bootstrap. Les individus en rouge sont ceux ne nécessitant pas de repas de sang pour pondre et qui seraient donc des formes molestus. En Allemagne et en Angleterre, ce sont des populations souterraines mais elles sont aériennes dans le reste du monde. Les individus en bleu forment des populations aériennes et nécessitent un repas de sang pour pondre ; il s’agirait donc des formes pipiens. Culex quinquefasciatus est un autre représentant du genre Culex, mais pas nécessairement une espèce à part puisqu’il existe des hybrides avec C. pipiens.
Par ailleurs, l’étude de Fonseca et al. montre que plus de 40 % des moustiques présents aux États-Unis seraient des hybrides de pipiens et de molestus venant d’Europe. Ces hybrides américains posent un problème de santé publique car ils piquent à la fois les oiseaux et les humains, et jouent donc un rôle important dans la propagation d’épidémies, comme celle du virus du Nil occidental.
Les auteurs terminent leur étude en se demandant pourquoi les hybrides sont si courants aux États-Unis alors qu’ils semblent absents dans le nord de l’Europe et peu fréquents dans le sud.
Des hybrides en Europe
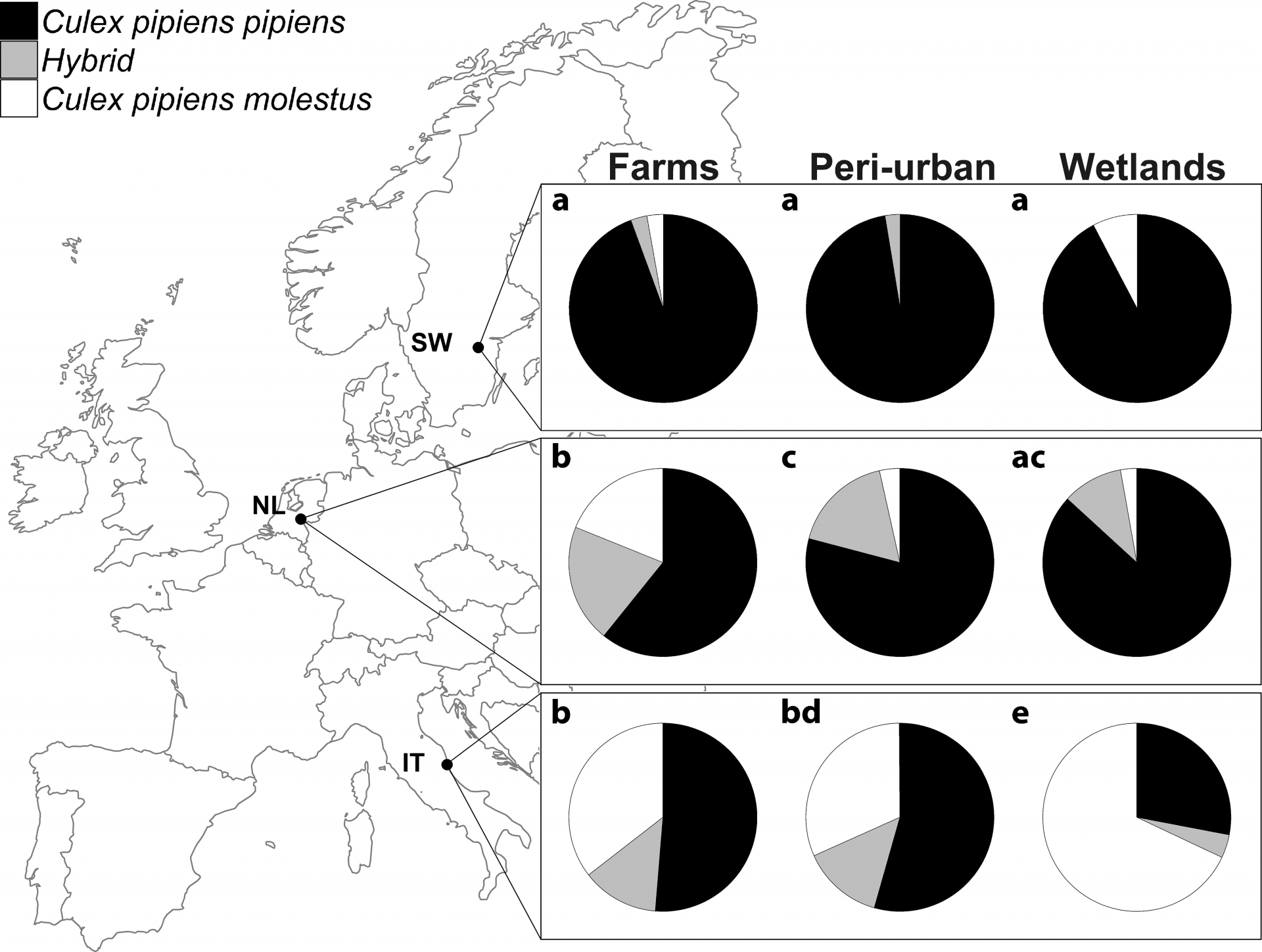
Répartitions des différents biotypes de Culex pipiens et de leurs hybrides en fonction de l’habitat dans un même pays (en lignes) et en fonction du pays pour un même habitat (en colonnes). La taille de l’échantillon de chaque graphique varie entre n = 26 et n = 115. Les lettres montrent les différences significatives entre les ratios présentés en lignes et en colonnes avec p < 0,05. SW : Suède, NL : Pays-bas, IT : Italie.
Deux études plus récentes ont cependant montré que les hybrides entre pipiens et molestus existent bel et bien dans toute l’Europe, même s’ils sont plus fréquents aux basses latitudes qu’aux hautes latitudes (Rudolf et al., 2013; Vogels et al., 2016).
Cependant, dans ces travaux, les auteurs ont identifié les populations de pipiens et de molestus en réalisant des PCR sur un seul locus (le microsatellite CQ11), quand Byrne et Fonseca utilisaient dans leurs études de nombreux locus pour discriminer les deux types de populations.
Les travaux de Rudolf et Vogels, qui amèneraient à considérer que pipiens et molestus sont deux sous-espèces d’une même espèce, sont donc à considérer avec précaution.
Conclusion
Que retenir de l’ensemble de ces études ? Tout d’abord, on peut remarquer que les contradictions apparentes entre les résultats de telle ou telle équipe résultent de deux phénomènes.
Premièrement, se pose le problème de l’échantillonnage. Lorsque Fonseca et al. concluent qu’il n’existe pas d’hybrides au nord de l’Europe c’est probablement parce que cette région est représentée dans leur étude par dix points de collecte « seulement », dont sept en Grande-Bretagne. Des études plus locales, se focalisant sur un pays en particulier, mais multipliant les points de collecte et considérant des échantillons de plus grande taille, mettent ainsi facilement en évidence les hybrides (Rudolf et al., 2013; Vogels et al., 2016).
Deuxièmement, les résultats peuvent également varier à cause de méthodes d’identification différentes des populations de Culex pipiens. Alors que Byrne et Fonseca distinguent pipiens et molestus grâce à l’étude de plusieurs locus, Rudolf et Vogels n’en observent qu’un.
Ensuite, il faut faire attention à la manière dont l’étude de Byrne et Nichols a pu être vulgarisée. D’une part, il faut bien comprendre que les formes molestus existaient, dans de nombreux pays, bien avant la construction du métro de Londres. Ce n’est donc pas la construction du métro qui a fait « apparaître » cette nouvelle forme.
Le corollaire est qu’il ne faut pas présenter le fait de se nourrir de sang humain ou de pouvoir se reproduire en milieu confiné comme des adaptations de molestus à la vie du métro. En effet molestus existait avant la construction du réseau souterrain et présentait déjà ces caractéristiques.
Enfin, il ne semble pas possible de dire que les formes molestus et pipiens correspondent à deux espèces différentes puisque des hybrides sont retrouvés, dans toute l’Europe et aux États-Unis. Il faut cependant bien distinguer ce qui se passe en Europe septentrionale, où pas ou peu d’hybrides sont identifiés, et ce que l’on observe plus au sud, où les populations de pipiens et de molestus se croisent régulièrement. Cela pourrait s’expliquer par des pressions de sélection différentes entre zones géographiques : au nord de l’Europe les conditions de vie des populations aériennes, soumises à des variations saisonnières de température de fortes amplitudes, contrastent avec celles des populations souterraines (où les variations de températures sont minimes). Ces différences de conditions de vie entre formes pipiens et molestus sont bien moins marquées dans le sud de l’Europe.
Pour en revenir aux populations de Londres, Byrne et Nichols ont montré que le flux génétique entre molestus et pipiens était très réduit, mais ne concluent pas quant à l’existence de deux espèces. Une preuve intéressante, mais a priori inexistante à ce jour, serait de voir si ces moustiques peuvent se reproduire dans des conditions naturelles. Dans l’affirmative, les molestus et pipiens de Londres correspondraient à des sous-espèces. Sinon, l’on pourrait bien parler de deux espèces différentes et donc de spéciation.
Plutôt que de chercher à tout prix à ranger molestus et pipiens dans des cases fixes et définitives, les spécialistes de Culex pipiens préfèrent utiliser pour ces populations la notion de complexe d’espèces. Celui-ci se définit comme un ensemble de taxons, issus d’un ancêtre commun récent, et qui sont en cours de différenciation. Les barrières reproductives séparant ces différents taxons sont ainsi plus ou moins perméables. Ceci illustre bien ce que l’on observe actuellement pour les formes molestus et pipiens de Culex pipiens, ainsi que pour d’autres populations apparentées (C. quinquefasciatus et C. pipiens pallens par exemple).
Références
- Abercrombie, M., Hickman, C.J., and Johnson, M.L. (1957). A Dictionary of Biology (Transaction Publishers).
- Chanet, B., Mouge, S., and Tostivint, H. Enseigner Classification Evolution (ECEV): Observer l’évolution aujourd’hui.
- Forêt, R. (2012). Dico de Bio: 11,000 définitins pour un panorama complet des sciences de la vie. (Bruxelles: De Boeck supérieur s.a.).
- Lecointre, G., Fortin, C., Bénéteau, A., Haessig, T., and Visset, D. (2009). Guide critique de l’évolution (Paris: Belin).
- Rudolf, M., Czajka, C., Börstler, J., Melaun, C., Jöst, H., Thien, H. von, Badusche, M., Becker, N., Schmidt-Chanasit, J., Krüger, A., et al. (2013). First Nationwide Surveillance of Culex pipiens Complex and Culex torrentium Mosquitoes Demonstrated the Presence of Culex pipiens Biotype pipiens/molestus Hybrids in Germany. PLOS ONE 8, e71832.
- Vogels, C.B.F., Möhlmann, T.W.R., Melsen, D., Favia, G., Wennergren, U., and Koenraadt, C.J.M. (2016). Latitudinal Diversity of Culex pipiens Biotypes and Hybrids in Farm, Peri-Urban, and Wetland Habitats in Europe. PLOS ONE 11, e0166959.


