

La voie Wnt canonique est impliquée dans la polarisation de l'axe antéro-postérieure chez de nombreuses espèces animales. Mais de nouvelles données obtenues chez l'hémichordé Saccoglossus kowalevskii viennent remettre en cause le modèle de régionalisation couramment admis chez les Vertébrés.
La voie de signalisation Wnt
Au sein d’un organisme en formation ou au cours de la vie adulte, les cellules communiquent entre elles par l’intermédiaire de molécules qui peuvent voyager à plus ou moins longue distance. La réception de ces messagers conduit à des réponses cellulaires variées (changement d’identité, division, migration…). Chez les animaux, ces voies de signalisation sont en nombre réduit (voies FGF, Hedgehog, JAK/STAT…) et ont été étudiées en profondeur. En effet, elles ont un rôle majeur dans le contrôle du développement embryonnaire, mais aussi dans l’homéostasie chez l’adulte. Étant aussi impliquées dans de nombreuses maladies (malformations congénitales, cancer…), elles font l’objet d’un intérêt majeur dans le domaine biomédical. Le contrôle de leur activité est aussi essentiel à la mise en œuvre de thérapies utilisant les cellules souches.
La voie de signalisation Wnt illustre ce type de convergence de plusieurs pans de recherche. Des études chez la souris visant à identifier des proto-oncogènes (gènes dont une mutation peut causer un cancer) ont identifié le gène int1 dont l’équivalent chez la mouche était connu sous le nom de wingless pour son rôle dans l’embryogenèse. La fusion de ces deux noms a conduit au nom Wnt qui désigne le ligand qui est sécrété à l’extérieur de la cellule. Après fixation sur son récepteur Frizzled à la surface de la cellule réceptrice, une cascade de signalisation intracellulaire conduit à une réponse spécifique de la cellule.
On distingue 3 signalisations distinctes (Fig. 1) :
- la voie canonique ou voie Wnt/β-caténine qui fait intervenir la β-caténine comme médiateur prépondérant. En absence de ligand, cette protéine est soumise à une dégradation par ubiquitination. En présence de ligand, cette dégradation est bloquée, les niveaux de β-caténine augmentent et celle-ci est transloquée dans le noyau où elle agit, avec des protéines de la famille TCF/LEF, comme facteur de transcription. La réponse de la cellule est donc l’activation de l’expression d’une palette de gènes.
- la voie calcique ne fait pas intervenir la β-caténine. La fixation du ligand à son récepteur conduit à une libération de Ca2+ depuis le réticulum endoplasmique ce qui, au final, régule l’adhérence et la migration cellulaires.
- la voie de polarité planaire (PCP, planar cell polarity) ne fait pas non plus intervenir la β-caténine, mais régule le cytosquelette d’actine.
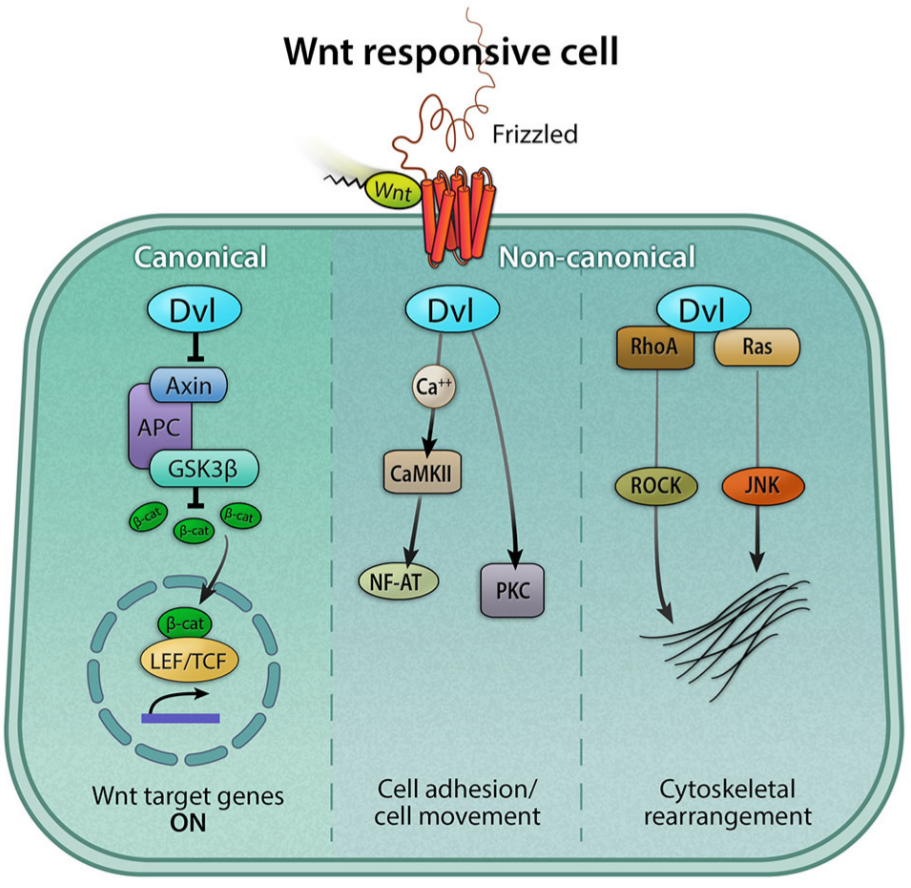
Voie canonique : activation des gènes cibles. Voie calcique : contrôle de l’adhérence et de la motilité des cellules. Voie PCP : réorganisation du cytosquelette.
Un rôle ancien de la voie Wnt dans la formation de l’axe primaire
La voie Wnt a une origine ancienne et sa présence est avérée dans toutes les branches du règne animal où elle a été recherchée. Elle est impliquée dans de très nombreux processus développementaux tels que l’acquisition de l’identité cellulaire, la différenciation, la polarité cellulaire ou la prolifération. La voie canonique participe en particulier, chez de nombreux animaux, à l’établissement de l’axe primaire de l’embryon (l’axe antéro-postérieur chez les bilatériens et l’axe oral-aboral chez les cnidaires) (Fig. 2).
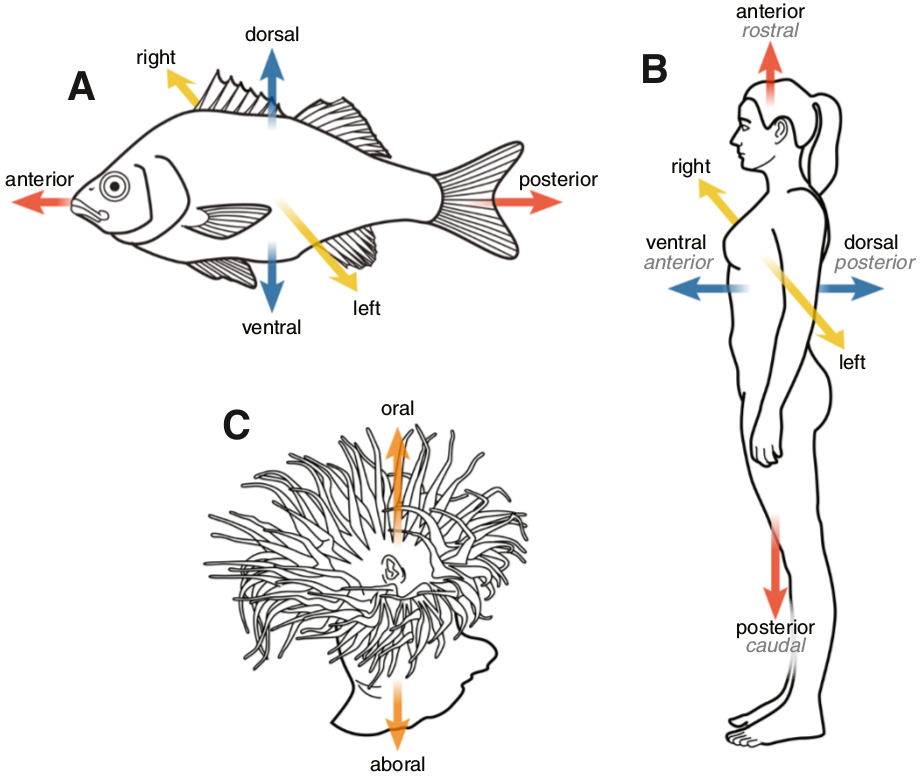
Chez les « poissons » (A) et chez les humains (B), l’axe antéro-postérieur est représenté en rouge, l’axe dorso-ventral en bleu et l’axe gauche-droite en jaune. Un cnidaire (une anémone de mer) possède un axe oral-aboral représenté en organe.
Chez de nombreux métazoaires, les ligands Wnt sont globalement et schématiquement exprimés dans la partie postérieure/orale et les antagonistes (molécules sécrétées inhibant la voie) dans la partie antérieure/aborale. Cette répartition spatiale binaire agonistes/antagonistes et les effets de la modulation de la voie ont conduit à proposer le modèle suivant : l’activation de la voie Wnt à un pôle de l’embryon définit la partie postérieure/orale et son inhibition au pôle opposé définit la partie antérieure/aborale [1], [2]. Bien que simple et élégant, ce modèle semble, à mon avis, confondre ou mélanger plusieurs processus. L’étude de l’embryon d’hémichordé nous a permis, avec mes collègues aux États-Unis, de raffiner ce modèle et je souhaiterais présenter ici une version plus nuancée sur le rôle de la voie canonique dans la formation de l’axe antéro-postérieur en distinguant trois fonctions majeures.
Les hémichordés et l’histoire évolutive des deutérostomiens
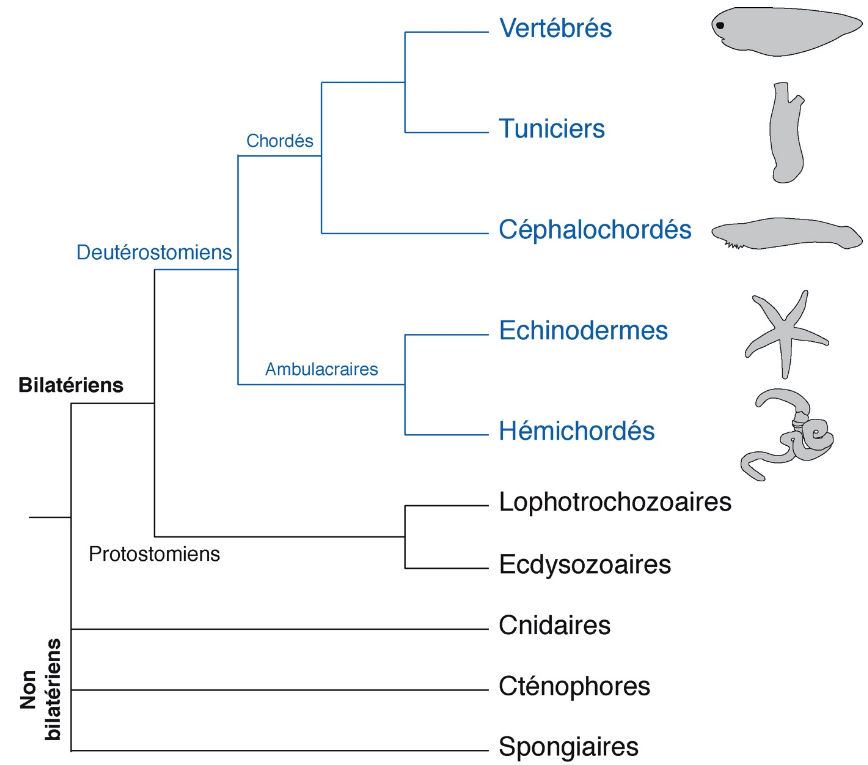
Une manière de comprendre l’évolution morphologique des animaux est d’effectuer des études de développement comparé d’animaux plus ou moins proches dans l’arbre phylogénétique. Plus spécifiquement pour comprendre l’origine évolutive des vertébrés, groupe animal auquel nous appartenons, il faut s’intéresser aux deutérostomiens (Fig. 3).
Les deutérostomiens comprennent 2 grands groupes : les chordés (vertébrés, tuniciers et céphalochordés) et les ambulacraires (échinodermes et hémichordés). Alors que les échinodermes ont acquis secondairement une symétrie pentaradiale, les hémichordés présentent toujours la symétrie bilatérale ancestrale. Comprendre comment s’établit cette symétrie et donc comment se mettent en place les axes de polarité au cours du développement embryonnaire permettrait de reconstituer la mise en place de ces axes chez l’ancêtre commun des deutérostomiens [3], [4]. Nous avons étudié le rôle de la voie Wnt canonique au cours du développement de l’embryon de Saccoglossus kowalevskii [5], [6]. Cet hémichordé de la catégorie des entéropneustes est un ver marin qui vit dans les sédiments. Il a un développement direct ; après 3-4 jours de développement éclot un petit ver qui présente déjà l’organisation du corps de l’adulte (Fig. 4). Cela permet d’analyser directement l’effet de perturbations moléculaires au cours du développement embryonnaire sur la mise en place du plan d’organisation caractéristique des hémichordés. De manière importante, son ectoderme est régionalisé le long de l’axe antéro-postérieur par la même combinatoire de facteurs de transcriptions que le système nerveux central des vertébrés, en particulier par les produits des gènes Hox pour le cerveau postérieur et la moelle épinière [7], [8].
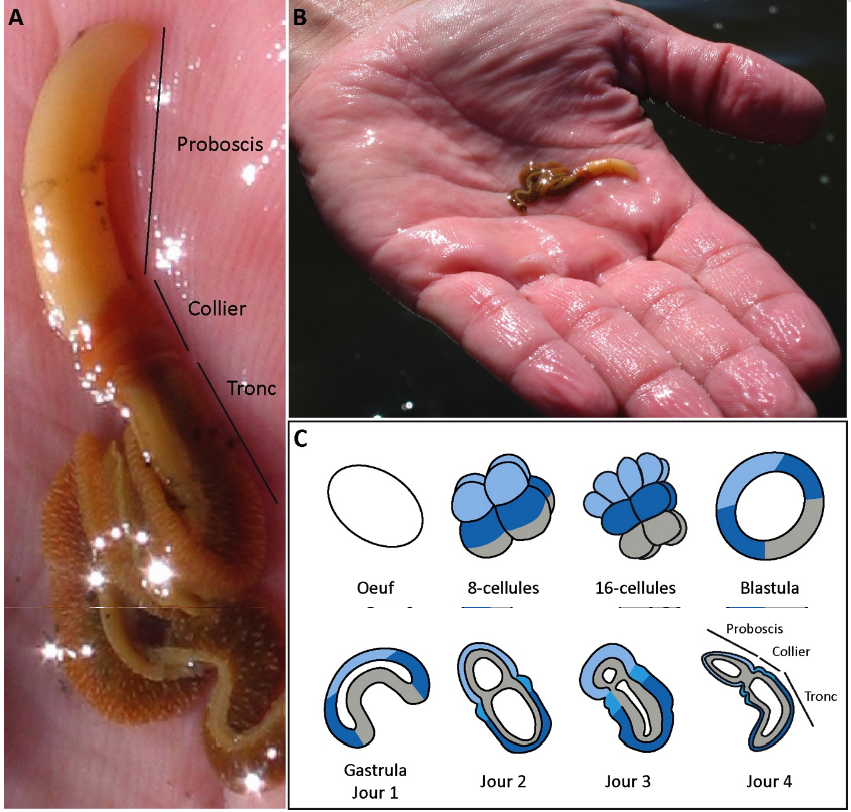
On distingue chez l’adulte (en gros plan en A et au moment de la collecte en B) l’organisation tri-partite de l’antérieur vers le postérieur : le proboscis blanc, le collier orange et le tronc marron. La même organisation est déjà visible à l’issue de l’embryogenèse chez le juvénile (C, jour 4). C) Illustration schématique des principales phases du développement embryonnaire. L’endomésoderme qui est internalisé est en gris, l’ectoderme est en bleu (bleu clair : antérieur ; bleu moyen : intermédiaire et bleu foncé : postérieur.
Un rôle précoce et ancestral pour la voie Wnt/β-caténine : la formation des feuillets germinaux
La fonction la plus précoce de la voie Wnt/β-caténine a été initialement établie chez l’embryon d’oursin. Lors du clivage (série de divisions cellulaires ayant lieu après la fécondation et produisant un embryon constitué de nombreuses cellules), la protéine β-caténine est détectée dans le noyau des cellules de l’hémisphère végétatif (signe de l’activation de la voie canonique). La voie Wnt canonique est nécessaire et suffisante à la spécification de l’endomésoderme (tissu embryonnaire produisant à la fois de l’endoderme et du mésoderme). La même fonction a ensuite été retrouvée chez d’autres animaux à symétrie bilatérale (autres échinodermes, ascidies, hémichordés, némertes), mais aussi chez les cnidaires. Cette dernière observation suggère donc une fonction de spécification de l’endomésoderme très ancienne qui a d’ailleurs été perdue ou modifiée chez les vertébrés.
La définition des feuillets germinaux par la voie Wnt canonique le long de l’axe animal-végétatif a des conséquences indirectes sur l’axe antéro-postérieur. En effet, les cellules de l’endomésoderme ont très souvent un rôle précoce pour organiser l’embryon. Chez les chordés, ces cellules sont essentielles par exemple pour spécifier et régionaliser le système nerveux lors de l’induction neurale. Chez l’hémichordé Saccoglossus kowalevskii, nous avons montré que l’endomésoderme défini par la β-caténine agit comme un organisateur en instruisant l’ectoderme à adopter une identité postérieure [5] (Fig. 5).
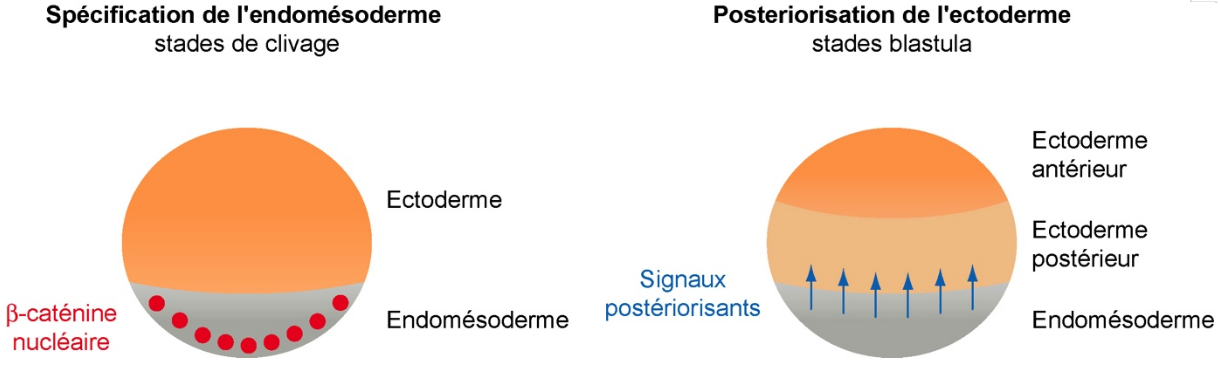
La voie est active aux stades de clivage (localisation nucléaire de la β-caténine) et spécifie le destin endomésoderme. Ce tissu ainsi spécifié agit comme organisateur en envoyant des signaux postériorisants à l’ectoderme sus-jacent.
Régionalisation de l’axe antéro-postérieur par la voie Wnt canonique
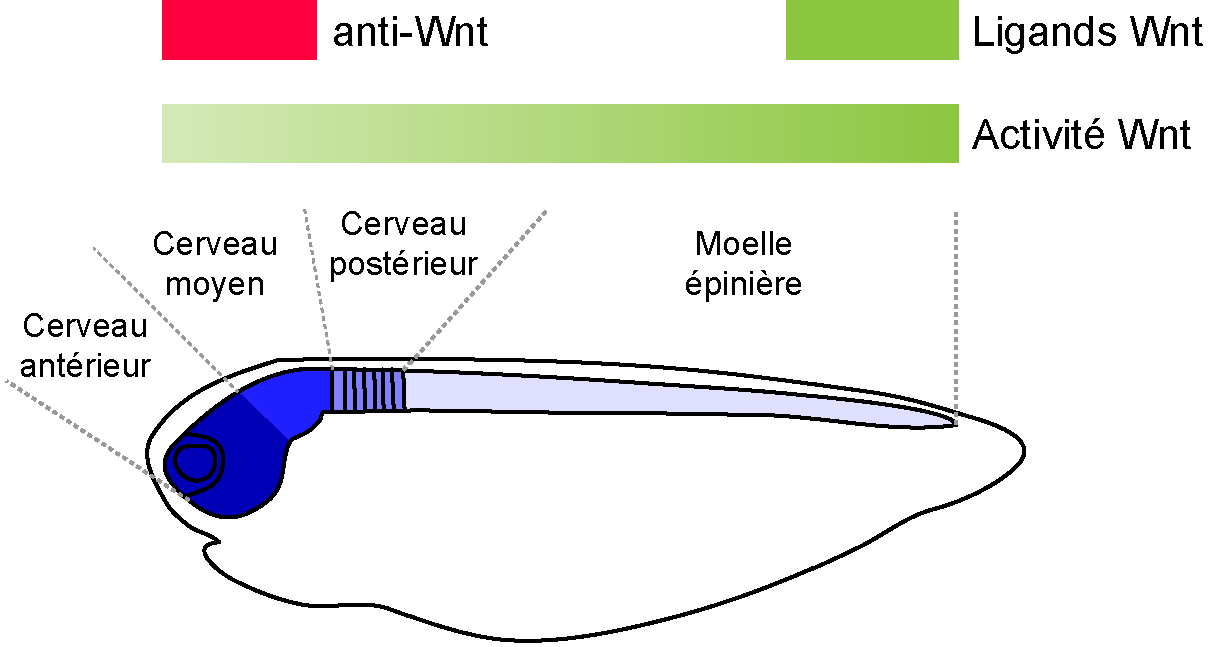
Selon ce modèle l’axe antéro-postérieur est régionalisé par un gradient d’activité de la voie Wnt canonique décroissant du postérieur vers l’antérieur.
La voie Wnt régionalise le système nerveux central des vertébrés. Il a été montré que l’expression de ligands postérieurement et d’antagonistes antérieurement permettait d’établir un gradient d’activité Wnt canonique décroissante le long de l’axe postéro-antérieur. De manière importante, ces niveaux d’activité instruisent les cellules quant à leur position au sein de la plaque neurale et déterminent leur identité en activant une palette d’acteurs moléculaires. Le modèle couramment admis est donc qu’un gradient Wnt régionalise l’intégralité du système nerveux des vertébrés (Fig. 6) [1], [2].
Une des implications de ces observations est que la partie la plus antérieure de l’embryon des vertébrés se forme par inhibition active de la voie Wnt et que, par conséquent, l’activation de la voie abolit l’antérieur. Ceci est observé chez d’autres deutérostomiens (céphalochordés et échinodermes), mais aussi chez plusieurs protostomiens. En revanche, en dehors des vertébrés, le rôle plus global de la voie Wnt dans la régionalisation de l’axe complet, et non du seul pôle antérieur, semble absent chez les autres deutérostomiens où il a été étudié (céphalochordés et échinodermes). Cela suggère que l’utilisation d’un gradient Wnt pour régionaliser l’intégralité de l’axe antéro-postérieur est une innovation des vertébrés au sein des deutérostomiens.
Cependant, deux cas dans lesquels la voie Wnt joue un rôle de régionalisation globale de l’axe suggèrent un rôle plus ancien : la régionalisation de l’axe oral-aboral au cours du développement de certains cnidaires et de l’axe antéro-postérieur au cours de la régénération des planaires. La comparaison de ces phénomènes avec la régionalisation du système nerveux central des vertébrés est néanmoins difficile. Tout d’abord, l’homologie entre axe antéro-postérieur des bilatériens et axe oral-aboral des cnidaires est fortement débattue. Enfin, il est loin d’être établi que les mécanismes qui maintiennent les polarités de l’organisme au cours de la régénération soient les mêmes que ceux impliqués lors de leur mise en place pendant l’embryogenèse.
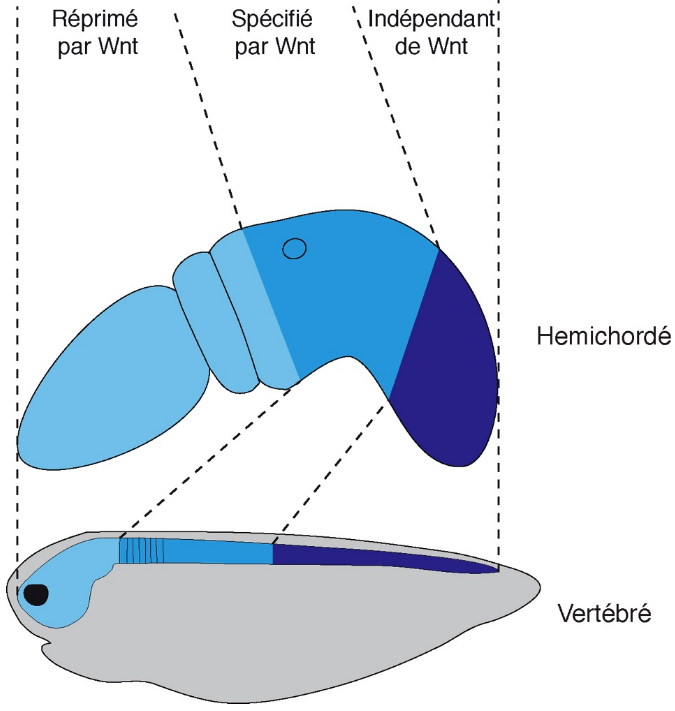
Chez les hémichordés et les vertébrés, la partie la plus antérieure ne peut se mettre en place qu’en l’absence de Wnt. La partie intermédiaire nécessite la voie Wnt tandis que la mise en place de la région postérieure est indépendante de Wnt.
L’histoire évolutive de la fonction de la voie Wnt au cours de la diversification des métazoaires est donc incertaine, principalement à cause d’un échantillonage faible des différents phyla, en particulier au sein des protostomiens. Afin de clarifier le rôle de la Wnt dans la régionalisation antéro-postérieure au sein des deutérostomiens, nous avons étudié spécifiquement son implication dans l’établissement de l’identité des cellules le long de l’axe antéro-postérieur chez l’hémichordé Saccoglossus kowalevskii [6]. Nous avons montré que, comme chez de nombreuses espèces, les ligands Wnt sont globalement exprimés postérieurement et que plusieurs antagonistes sont exprimés antérieurement. En utilisant des approches d’embryologie classique et en modulant l’activité Wnt canonique (par des approches pharmacologiques ou moléculaires), nous avons pu montrer , que l’ectoderme de S. kowalevskii est subdivisé en 3 régions : une partie antérieure spécifiée par l’inhibition active de Wnt, une partie intermédiaire activement définie par Wnt et une partie postérieure insensible à Wnt. Ces 3 régions correspondent respectivement aux cerveaux antérieur et moyen, au cerveau postérieur, et à la moelle épinière des vertébrés. Ces résultats nous ont amené à suggérer un nouveau modèle pour le rôle de la voie Wnt dans l’établissement de la polarité antéro-postérieure des deutérostomiens. Alors que l’on considérait jusqu’ici que la partie la plus postérieure de l’embryon était spécifiée par une forte activité Wnt (Fig. 6), nous proposons que celle-ci est au contraire mise en place indépendamment de la voie Wnt (Fig. 7).
La croissance postérieure
Chez de nombreux animaux, le corps s’allonge au cours de la vie embryonnaire par addition d’éléments dans la partie postérieure. C’est ce qu’on appelle la croissance ou élongation postérieure. Chez les vertébrés, par exemple, plusieurs structures axiales s’allongent postérieurement (la notochorde, la moelle épinière…) grâce à une structure appelée le bourgeon caudal. Cette phase d’allongement est plus tardive au cours de la vie embryonnaire et a lieu après les deux phases mentionnées plus haut, la détermination des feuillets germinaux et la régionalisation antéro-postérieure. Là encore la voie Wnt est impliquée et promeut la formation de structures additionnelles, donc postérieures. De manière frappante, l’élongation postérieure existe chez les vertébrés et chez les insectes et fait intervenir les mêmes acteurs moléculaires, dont la voie Wnt [2], [9].
Chez l’hémichordé Saccoglossus kowalevskii, la définition initiale du territoire le plus postérieur aux stades blastula/gastrula ne fait pas intervenir la voie Wnt. Néanmoins, plus tard au cours du développement embryonnaire (après la gastrulation), le maintien de cette identité est là régulé par Wnt [6]. Ceci est peut-être le reflet d’un rôle de la voie Wnt dans cette phase tardive de croissance postérieure qui existe aussi chez l’hémichordé mais qui n’a pas encore été étudiée.
Conclusion
La voie Wnt a une origine ancienne chez les métazoaires et des rôles variés au cours de l’embryogenèse. En particulier, la voie canonique régule trois processus développementaux qui ont chacun un impact sur la formation de l’axe antéro-postérieur (ou oral/aboral chez les cnidaires) :
- la détermination des feuillets germinaux
- la régionalisation antéro-postérieure
- la croissance postérieure
L’atout de l’étude de l’embryon d’hémichordé, outre sa position phylogénétique, est que ces 3 processus sont séparés temporellement au cours de l’embryogenèse et donc découplables expérimentalement.
Bibliographie
- C. Niehrs, « On growth and form : a Cartesian coordinate system of Wnt and BMP signaling specifies bilaterian body axes », Development, vol. 137, p. 845‑57, mars 2010.
- C. P. Petersen et P. W. Reddien, « Wnt signaling and the polarity of the primary body axis », Cell, vol. 139, p. 1056‑68, déc. 2009.
- J. Gerhart, « The deuterostome ancestor », J. Cell. Physiol., vol. 209, no 3, p. 677‑685, déc. 2006.
- C. J. Lowe, D. N. Clarke, D. M. Medeiros, D. S. Rokhsar, et J. Gerhart, « The deuterostome context of chordate origins », Nature, vol. 520, no 7548, p. 456‑465, avr. 2015.
- S. Darras, J. Gerhart, M. Terasaki, M. Kirschner, et C. J. Lowe, « {beta}-Catenin specifies the endomesoderm and defines the posterior organizer of the hemichordate Saccoglossus kowalevskii », Development, vol. 138, p. 959‑70, mars 2011.
- S. Darras et al., « Anteroposterior axis patterning by early canonical Wnt signaling during hemichordate development », PLOS Biol., vol. 16, no 1, p. e2003698, janv. 2018.
- C. J. Lowe et al., « Anteroposterior patterning in hemichordates and the origins of the chordate nervous system », Cell, vol. 113, p. 853‑65, juin 2003.
- J. Aronowicz et C. J. Lowe, « Hox gene expression in the hemichordate Saccoglossus kowalevskii and the evolution of deuterostome nervous systems », Integr. Comp. Biol., vol. 46, p. 890‑901, déc. 2006.
- B. L. Martin et D. Kimelman, « Wnt signaling and the evolution of embryonic posterior development », Curr Biol, vol. 19, p. R215-9, mars 2009.