
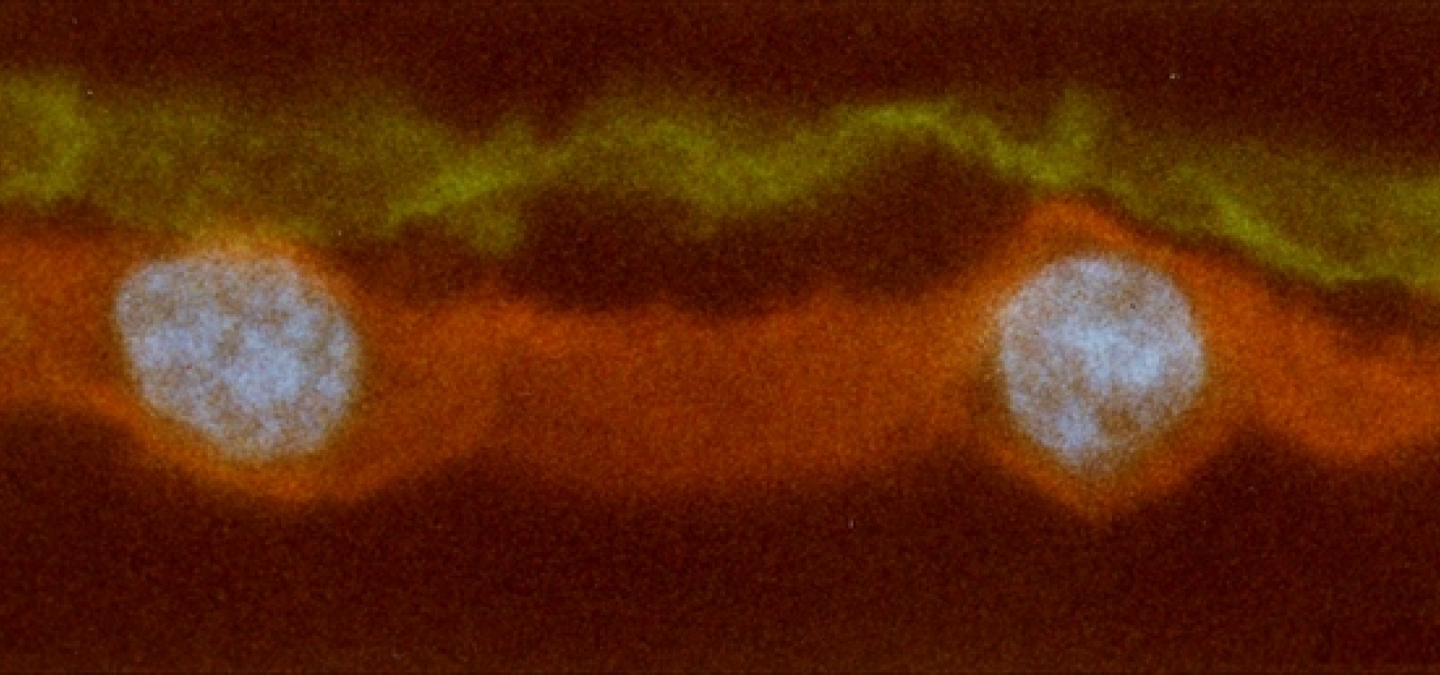
L’électrocyte du poisson torpille (Torpedo marmorata) est un syncytium (cellule comprenant plusieurs noyaux) de forme discoïdale pouvant atteindre plusieurs millimètres de diamètre. Cette cellule reçoit une intense innervation cholinergique (plusieurs centaines à plusieurs milliers de terminaisons par cellule) localisée sur sa face ventrale. Elle est la principale cellule de l'organe électrique du poisson torpille.
Cette image montre une coupe transversale d'un électrocyte. La face ventrale riche en terminaison nerveuse est repérée par l'utilisation d'une toxine de venin de serpent ayant une forte affinité pour le récepteur nicotinique de l'acétylcholine, l'α-bungarotoxine, couplée à un fluorochrome vert (le FITC). La face dorsale, non innervée, est repérée par l'utilisation d'un anticorps dirigé contre la pompe Na+K+, lequel anticorps est lui même révélé par un second anticorps dirigé contre le premier et couplé à un fluorochrome rouge (la rhodamine). Les noyaux sont repérés par l'utilisation de DAPI, une molécule fluorescente bleue ayant de l'affinité pour l'ADN.
On constate que la distribution des protéines membranaires est polarisée, avec le récepteur de l'acétylcholine présent exclusivement au niveau de la face ventrale, et la pompe Na+K+ au niveau de la face dorsale. Des accumulations locales similaires sont observées à plus petite échelle au niveau des jonctions neuromusculaires et des synapses centrales, mais aussi dans les cellules épithéliales. Parmi les mécanismes qui concourent à ces accumulations locales citons le tri des molécules néosynthétisées dans les voies de la biosynthèse, leur adressage spécifique à la membrane de résidence, la double stabilisation spatiale et métabolique de ces molécules une fois arrivées à destination, et une régulation différentielle de l'expression des gènes codant pour ces protéines.
Noter que la cellule étant un syncytium, elle est dotée de plusieurs noyaux, ici cernés par des replis de la membrane non-innervée.



