

Adaptations des animaux face aux variations de température. Étude des variations ultradiennes de températures chez des canetons de Barbarie soumis à différents stress.
Le physiologiste Claude Bernard remarquait, il y a plus d’un siècle, la différence entre le milieu extérieur, par définition très variable, et un milieu interne maintenu plus constant. L’homéostasie correspond à l’ensemble des réponses dynamiques au sein des organismes qui permettent cet équilibre interne malgré les variations de l’environnement. Au cours du XXe siècle, de nombreux mécanismes de régulation homéostatique, en particulier face aux variations thermiques de l’environnement, ont été décrits.
En effet, parmi toutes les variables environnementales, l’une des plus importantes est la température car elle affecte toutes les échelles du vivant, de la vitesse de réaction biochimique entre molécules à la distribution des espèces dans le milieu. Les variations de température constituent donc une contrainte forte pour tous les êtres vivants. Des mécanismes de compensation face à ces variations thermiques, permettant ainsi le maintien de l’homéostasie, ont été sélectionnés au cours de l’évolution. On peut citer par exemple les cas d’enzymes capables de garder une affinité équivalente avec leur substrat pour une large gamme de températures ambiantes, telles que la lactate déshydrogénase (LDH) ou la malate déshydrogénase (MDH), enzymes clés dans le métabolisme énergétique (Somero 2004).
La thermorégulation
Tous les animaux vivants dans des environnements soumis à des changements de température (augmentation ou diminution) doivent maintenir leur température interne dans un intervalle compatible avec la vie. Se pose alors la question de la température corporelle optimale qui détermine de nombreux paramètres physiologiques (comme la consommation de dioxygène, les capacités de digestion, la puissance musculaire, etc.) et écologiques (vitesse de croissance, capacité de reproduction, etc.).

On remarque bien sur ce cliché la différence de température entre les deux animaux.
Ectothermes vs endothermes
Deux stratégies sont ainsi principalement rencontrées chez les animaux : l’ectothermie et l’endothermie, en fonction de la capacité à produire ou non de la chaleur (Figure 1).
Les ectothermes (anciennement appelés « poïkilothermes ») ont leur température interne qui est le reflet de la température ambiante (Figure 2, courbe verte). Ce groupe, composé de l’immense majorité des animaux (Arthropodes, Annélides, Mollusques, « poissons », « reptiles », Amphibiens…), utilise presque exclusivement des adaptations comportementales pour augmenter ou diminuer sa température interne (qui peut être maintenu à des valeurs très élevées). Le principal avantage d’une telle stratégie est une économie d’énergie mais les organismes sont strictement liés aux variations de température externe.
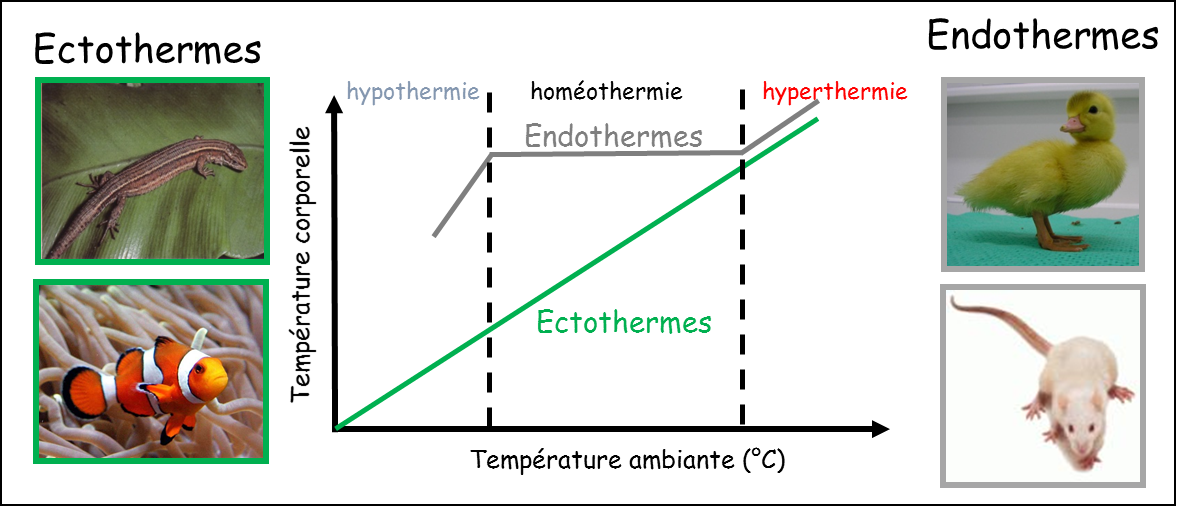
On remarque sur cette représentation graphique, que la température corporelle des ectothermes peut être parfois élevée. Du coup, l’appellation « animaux à sang froid » est un abus de langage…
En effet, l’autre groupe, composé uniquement des Mammifères et des Oiseaux est caractérisé par le maintien d’une température interne à un niveau élevé et constant, dans une plage de température ambiante donnée (Figure 2, courbe grise). Cette capacité de thermorégulation est due à un ensemble de mécanismes que l’on appelle thermogenèse (lors d’une production de chaleur) ou thermolyse (lors d’une évacuation de chaleur). Ces productions et/ou pertes de chaleur sont dues à des mécanismes internes, d’où le nom d’endotherme (du grec ancien « endon » : dans, « thermos » : chaud).
Bien qu’ils permettent de se mouvoir quelle que soit la température externe, ces processus de thermorégulation active présentent un inconvénient majeur pour les organismes endothermes : le coût énergétique important qui nécessite une consommation de dioxygène et un apport alimentaire très supérieur à ceux des ectothermes.
Hétérothermie : différentes échelles temporelles
Afin de limiter ce coût énergétique, certains endothermes ont développé des stratégies légèrement différentes, imprimant des variations de température interne en fonctions de cycles plus ou moins longs. Ce sont alors des organismes appelés « hétérothermes ». Les exemples les plus connus sont sans doute parmi les animaux hibernant, tels que les marmottes, les loirs, les hérissons ou encore certains écureuils. La température interne de ces animaux va significativement diminuer de plusieurs dizaines de degrés lors de la saison hivernale, évitant ainsi un surcoût énergétique lié au maintien de leur température interne à un niveau élevé. C’est finalement le même principe que la position « hors gel » des radiateurs d’une maison secondaire que l’on n’habite qu’une partie de l’année.
D’autres organismes hétérothermes vont montrer des variations de température interne en fonction du cycle circadien. Ce phénomène, appelé torpeur, est caractérisé par une diminution de quelques degrés de la température interne au cours de la nuit, puis un retour à un niveau plus élevé durant la journée.
Outre les cycles saisonniers ou infradien (hibernation, estivation) et les cycles journaliers ou circadiens (torpeur), il existe un cycle à une échelle de temps plus fine, le cycle ultradien, dont dépendent de nombreux phénomènes physiologiques (Figure 3).
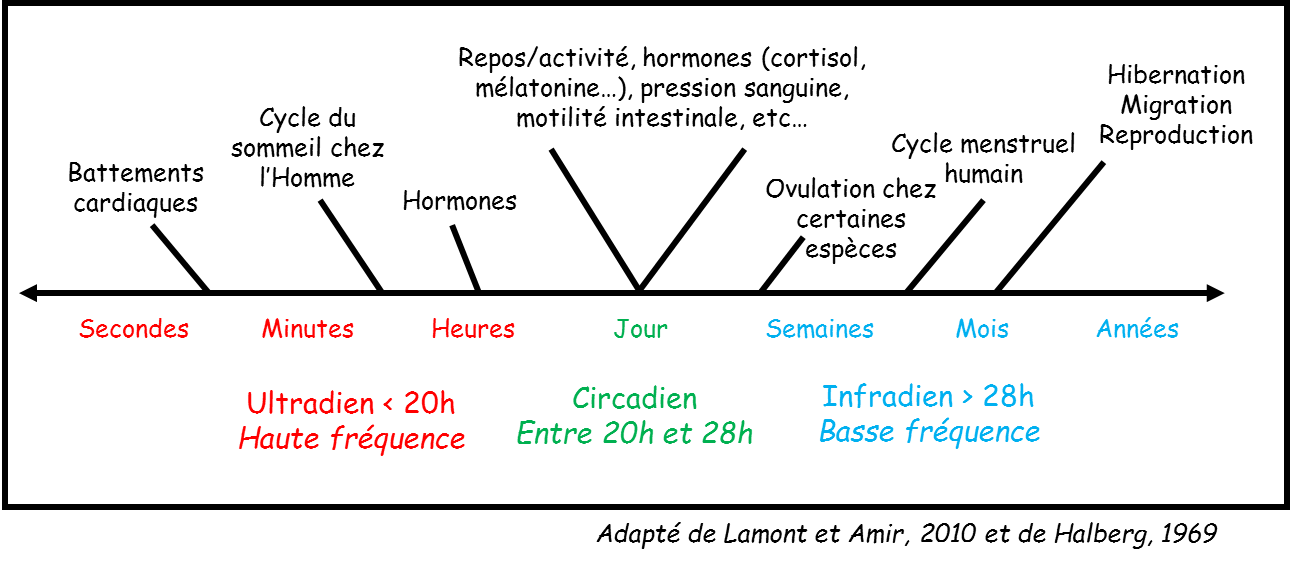
Régulation ultradienne au niveau physiologique
Les oscillations ultradiennes sont caractérisées par des cycles de durée inférieure à 20 h (Halberg, 1969). Elles ont été très étudiées au niveau hormonal, car il est relativement connu que les taux d’hormones varient de façon pulsatile au cours de la journée. Dans une revue bibliographique de 1991 (Lloyd and Stupfel, 1991), les auteurs font état des nombreux mécanismes physiologiques cadencés par les cycles ultradiens. C’est le cas par exemple du sommeil paradoxal chez l’homme qui est en moyenne déclenché toutes les 90 minutes, ou toutes les 180 minutes pour les phases de sommeil chez les nouveau-nés. Cependant, très peu d’éléments sont connus à ce jour sur le contrôle ultradien de la température corporelle chez les endothermes.
Thermorégulation corporelle pendant les phases de contraintes environnementales
Chez un endotherme, la température du corps est déterminée par l’équilibre entre la production de chaleur et les mécanismes de dissipation de chaleur. Une réponse métabolique bien connue face aux contraintes environnementales est la réduction de la production de chaleur, accompagnée par une réduction de température corporelle. Les fluctuations de la température corporelle peuvent résulter de changements dans le point de consigne, des changements d’activité, ou des modifications de la conductance thermique. Néanmoins, aucune étude n’avait à ce jour testé si la variabilité dans la température corporelle pouvait en elle-même être une source d’économies d’énergie en conditions stressantes. Ce concept a été formellement testé dans une étude menée par notre équipe et publiée en 2016 dans Proceedings of the Royal Society (part B).
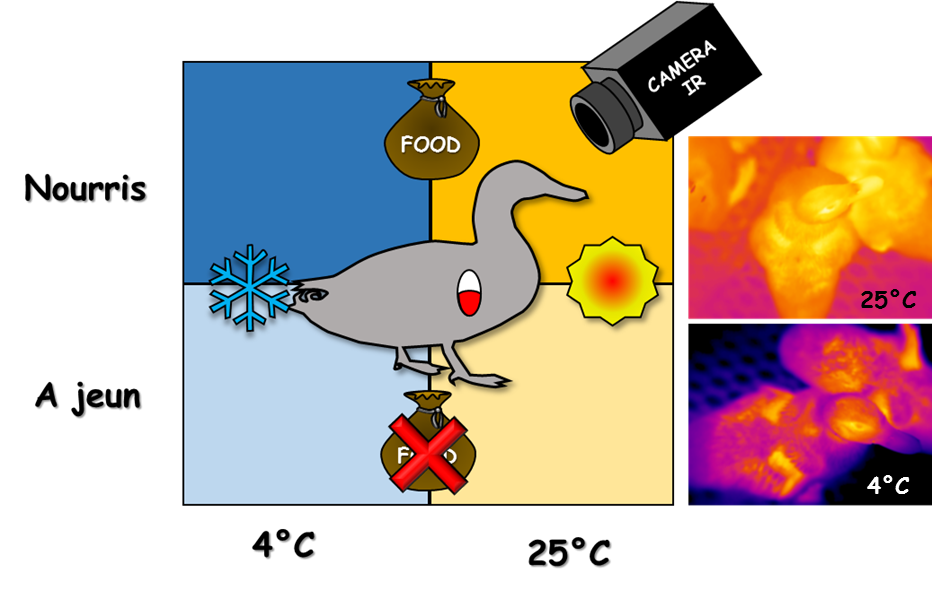
Deux conditions ont été croisées : la température (4 °C, bleu ou 25 °C, orange) et la disponibilité en nourriture. La température corporelle a été mesurée à l’aide de capteurs présents dans le gésier et d’une caméra infrarouge. À droite, deux images thermiques de canetons prises à 25 °C et à 4 °C.
L’étude a mobilisé des canetons de Barbarie âgés de 5 semaines, répartis en deux groupes égaux en fonction de la température environnante : les premiers ont été placés dans des conditions de froid intense (5 °C), alors que les seconds sont restés à neutralité thermique (25 °C). Les canetons ont été équipés de capteurs de température interne se présentant sous la forme de petites pilules stockées dans leur gésier tout au long de l’expérience. Puis chaque groupe a connu, sur trois périodes égales de 4 jours, une phase d’alimentation à volonté, puis un jeûne alimentaire et enfin une réalimentation. En plus du froid, les animaux ont donc été soumis à un stress supplémentaire : la disponibilité en nourriture. Les variations de la température corporelle ont alors pu être analysées très finement, les capteurs émettant des mesures toutes les 15 minutes. Parallèlement, une caméra thermique infrarouge a permis de mesurer les fluctuations de la température au niveau du bec des canetons (Figure 4).
Le premier résultat concerne tout d’abord la capacité des canetons à réguler leur température corporelle, à 4 °C comme à 25 °C lorsqu’ils sont nourris à volonté : on observe en effet des variations circadiennes de la température corporelle identiques chez les canetons à thermoneutralité et chez ceux soumis à un froid intense (Figures 5 A et B).
Lors de la phase de jeûne, la température corporelle a globalement baissé dans les deux cas. Bien qu’intuitif pour des canetons soumis à un froid intense, ce résultat est relativement étonnant car il touche également les canetons à neutralité thermique. La température interne a effectivement diminué, durant les phases de jour mais également durant les phases de nuit et ce, de manière un peu plus marquée, augmentant ainsi l’amplitude des variations de température corporelle.
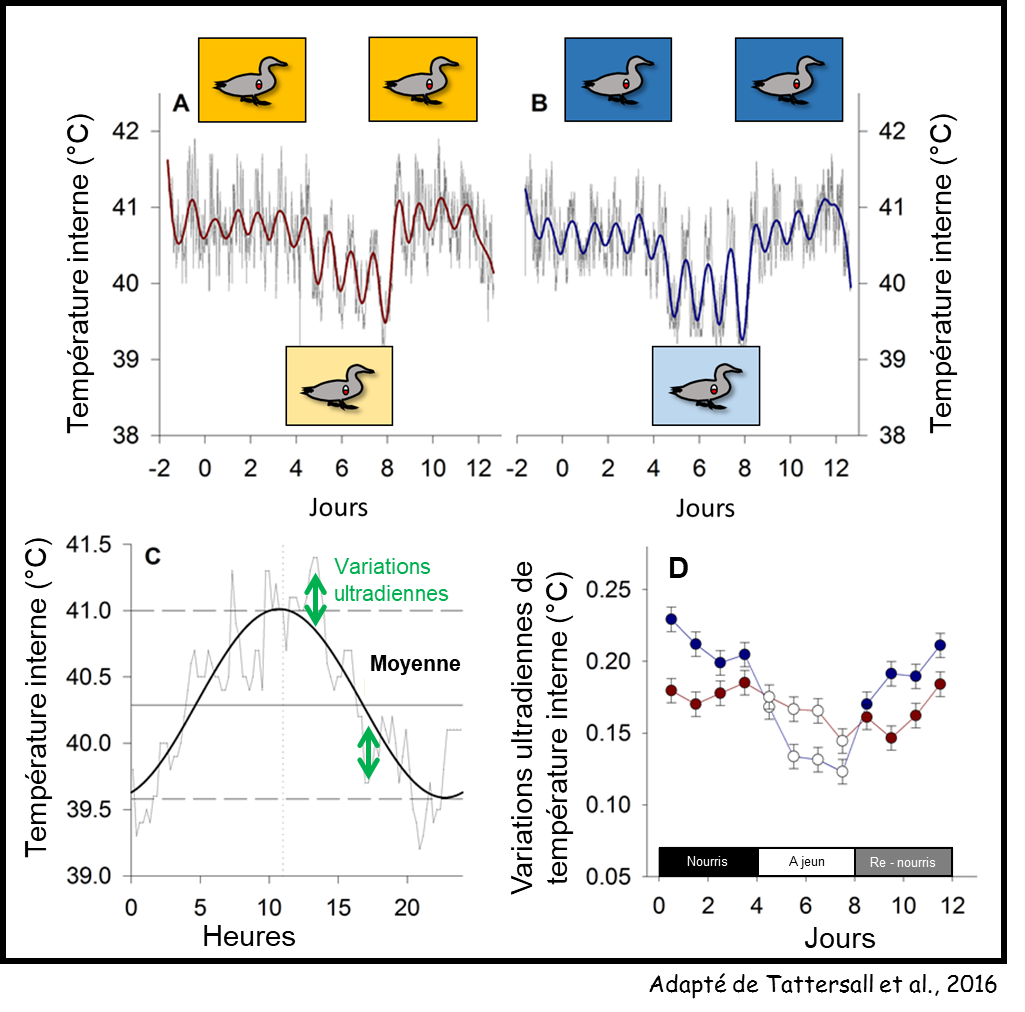
Suivi de température interne sur 14 jours chez des canetons à thermoneutralité (jaune, A) ou à 4 °C (bleu, B). Représentation de la variation de température interne d’un caneton élevé à thermoneutralité sur 24 h (C) et évolution de l’amplitude des variations ultradiennes de température interne en fonction de l’accessibilité à la nourriture (D).
Outre ces variations de température interne, d’autres indices d’une diminution de la thermorégulation ont été observés. Ainsi, la température du bec et d’autres extrémités comme les ailes a diminué par une vasoconstriction entraînant une diminution de l’afflux sanguin. Cela permet d’éviter de dissiper trop d’énergie et de conserver la chaleur au niveau des parties centrales comme le cerveau.
Les effets du jeûne alimentaire combinés à un stress thermique sont principalement visibles au niveau de la masse corporelle des oiseaux : chez les canetons exposés au froid intense, le jeûne a entraîné une perte de masse 20 % plus rapide que chez les canetons à neutralité thermique.
Concernant la régulation de la température corporelle, elle s’est produite de manière beaucoup plus précise : les variations ultradiennes de la température corporelle ayant significativement diminué (Figures 5 C et D).
Pour résumer, les canetons, qu’ils soient à température ambiante ou soumis à un froid intense, régulent de manière très fine leur température interne tant qu’ils sont nourris correctement. Mais ce n’est plus le cas à jeun : la contrainte thermique devient alors trop importante et entraîne une hypothermie principalement nocturne. L’étude montre ainsi que le coût énergétique des changements de température à très court terme est abaissé au maximum chez les canetons soumis au froid et au jeûne : le gain énergétique permis par la thermorégulation ultradienne est compris entre 7 et 10,5 % du métabolisme de base, ce qui est loin d’être négligeable car ce dernier est incompressible. L’énergie ainsi économisée pourrait alors être stockée ou affectée à d’autres postes comme la croissance, la reproduction, etc. Cette étude, la première montrant la diminution des variations ultradiennes de température interne chez les oiseaux, soulève de nombreuses questions sur la gestion de ces rythmes dans l’optimisation des stratégies de conservation énergétique chez les animaux lors de fortes pressions environnementales.
Références
Lamont EW, Amir S. Circadian and Ultradian Clocks/Rhythms. In : George FK, Michel Le M, Richard FT, editors. Encyclopedia of Behavioral Neuroscience. Oxford : Academic Press ; 2010. p. 257-261
Lloyd D., Stupfel M. 1991. The occurrence and functions of ultradian rhythms. Biological Reviews, Volume 66, Issue 3, Pages 275–299.
Halberg, F. 1969. Chronobiology. Annual review of physiology, 31(1), 675-726.
Somero G.N. 2004. Adaptation of enzymes to temperature : searching for basic “strategies”. Comparative Biochemistry and Physiology Part B.
Tattersall G.J., Roussel D., Voituron Y., Teulier L. 2016 A novel energy savings to multiple stressors in birds : The ultradian regulation of body temperature. Proceeding of the Royal Society B. 283 : 2016.1551.
