
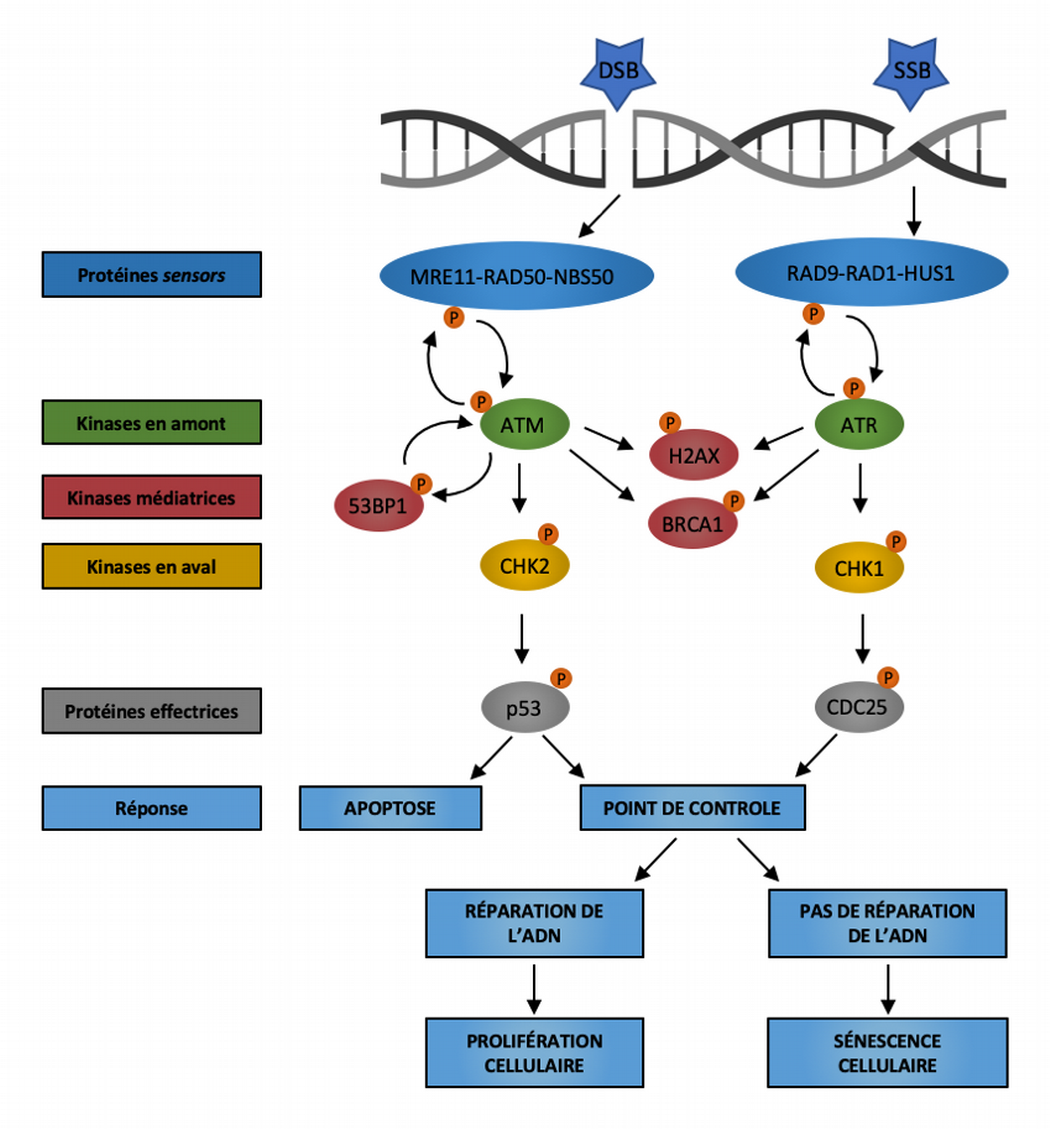
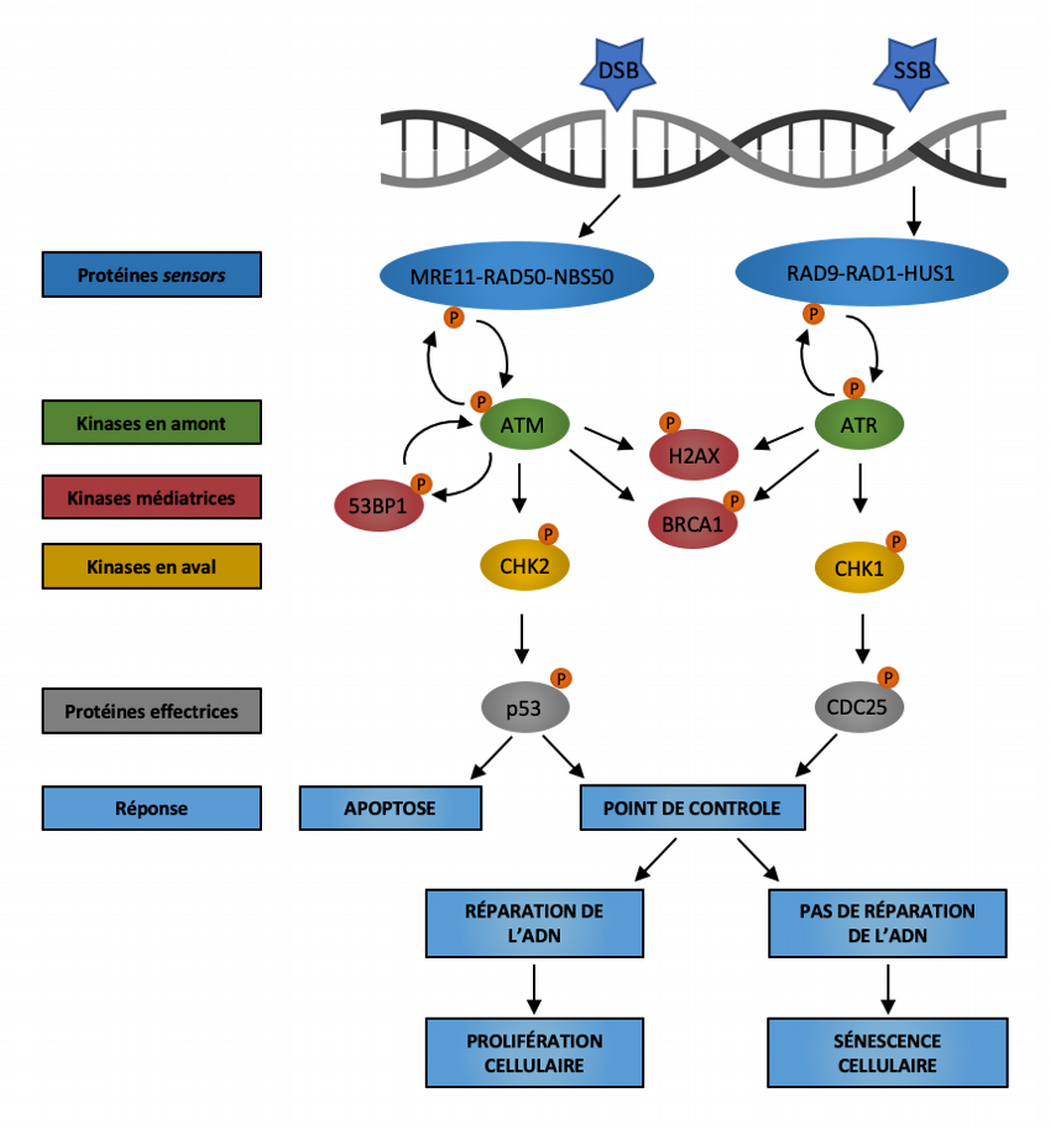
La voie de réponse aux dommages à l’ADN (DDR) repose sur deux principaux complexes protéiques senseurs détectant les dommages à l’ADN : le complexe MRE11-RAD50-NBS1 qui détecte les cassures double brin (DSB) et le complexe RAD9-RAD1-HUS1 qui détecte les régions exposées d’ADN monocaténaire (SSB). Ces senseurs recrutent alors les kinases ATM et ATR qui, à leur tour, phosphorylent le variant d’histone H2AX dans la région proximale de la lésion de l’ADN. Alors qu’ATM est principalement activée par les cassures double brin, ATR est essentiellement induite par les cassures simple brin causées par la réplication de l’ADN et l’hyperactivation d’oncogènes. La protéine BRCA1 est recrutée sur les sites d’altération de l’ADN après phosphorylation par ATM et ATR. La protéine 53BP1 participe également au maintien de la signalisation de réponse aux dommages de l’ADN en favorisant l’activation d’ATM. La suite de la signalisation DDR se déroule à distance du locus endommagé via le recrutement des kinases diffusibles CHK2 et CHK1, et converge vers des effecteurs en aval tels que la phosphatase CDC25 et le facteur de transcription p53. Les conséquences de l’activation de la réponse aux dommages de l’ADN peuvent être la mort cellulaire par apoptose, l’arrêt transitoire du cycle cellulaire suivi de la réparation des dommages à l’ADN et de la reprise de la prolifération, ou la sénescence cellulaire causée par la persistance des dommages non réparés de l’ADN. Modifié d’après [6].

{kind=link}