
Après avoir rappelé ce que signifie « classer les êtres vivants », quels en sont et quels en ont été, historiquement, les objectifs, cet article présente les principes de la classification phylogénétique moderne. Il présente également, pour le niveau scolaire primaire, une façon de classer n'utilisant pas de données évolutives, mais compatible avec elles. Il s'accompagne d'une bibliographie, d'un glossaire et de mises au point relatives à plusieurs idées fausses.
Introduction générale
On pourrait penser que classer les êtres vivants est une activité maîtrisée des chercheurs en systématique (science des classifications), des naturalistes amateurs, des enseignants et des utilisateurs de la nature (agriculteurs, sélectionneurs, agronomes, pharmaciens, vétérinaires, paysagistes, conservateurs de parcs et de musées, etc.). Il n’en est rien.
Même chez les scientifiques dont le métier utilise des classifications, il arrive encore trop souvent qu’on se méprenne sur ce qu’est que « classer ». Ceci est peut-être dû au fait qu’on a tous une intuition de ce en quoi consiste cette activité. Il peut paraître inutile alors de rappeler les fondements de l’activité classificatoire en général, et plus particulièrement en sciences naturelles. Faire des livres sur un aspect aussi fondamental des sciences semblerait aussi futile qu’enfoncer des portes ouvertes.
C’est mal connaître le contexte. Récemment, à l’occasion d’un changement de programme, beaucoup d’enseignants du secondaire découvrirent qu’ils n’avaient jamais classé, et que ce que les programmes et les livres scolaires présentaient comme de la « classification » n’en était pas...
Qu'est-ce que classer ?
Introduction
Dans les livres scolaires, on confondait encore il y a quelques années trier, assigner et classer. L'erreur la plus courante consistait à présenter une clé de détermination comme une classification.
Trier : la clé de détermination
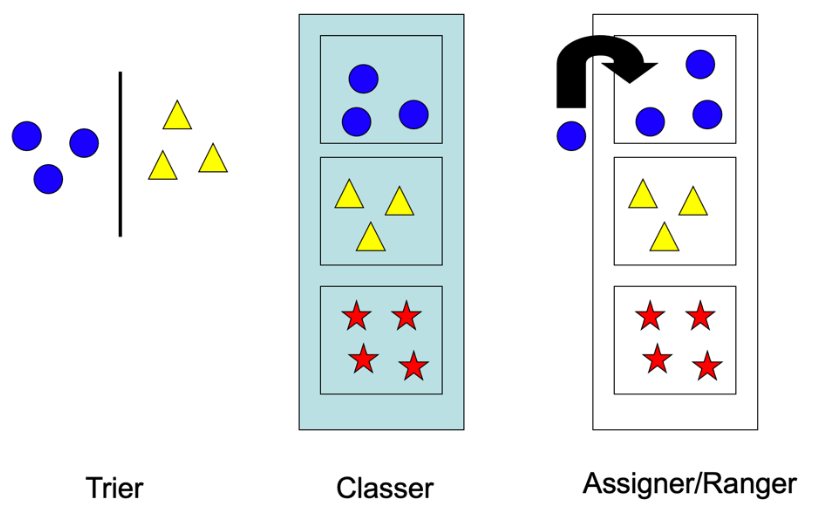
Trier, ce n’est pas tracer des ensembles, ce n’est pas créer des groupes. C’est simplement séparer un échantillon d’objets en fonction d’arguments arbitraires. Au sein de cet échantillon, on discrimine, on distingue, souvent avec une suite de critères de type présence/absence. La clé de détermination est un chemin décisionnel au cours duquel, à partir des attributs présentés par une espèce, l’utilisateur s’oriente vers le nom de l’espèce. C’est une sorte de jeu de piste le long d’une arborescence décisionnelle (Fig. 2). Pour fabriquer la clé de détermination, le concepteur a procédé par tris successifs, qu’il a organisés en une série précise de décisions à prendre. L’utilisateur, lui, ne trie pas vraiment mais s’oriente dans le produit de ce travail.
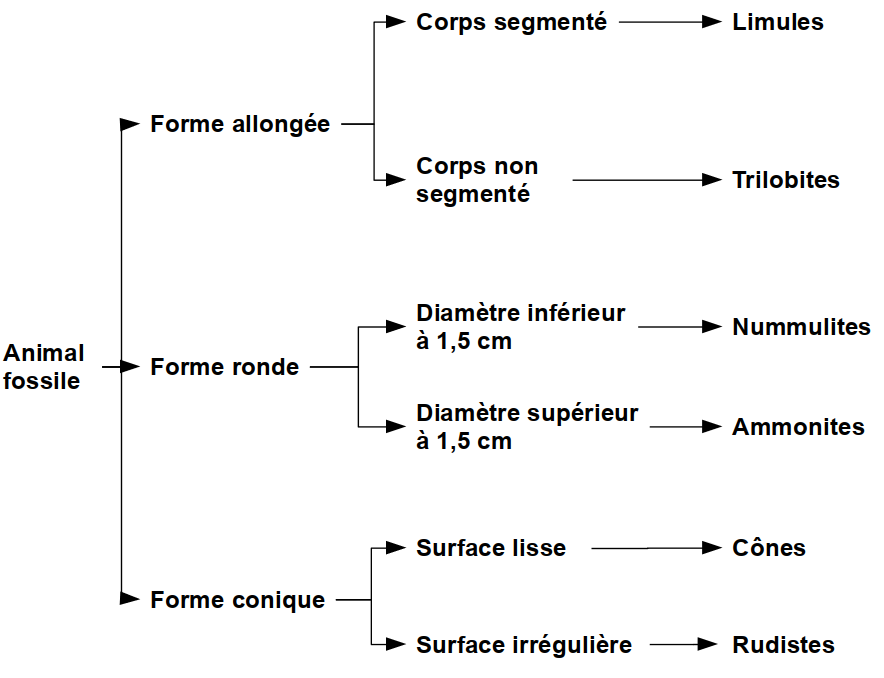
La clé de détermination a pour vocation d’être pratique. Une bonne clé doit faire aboutir au plus vite au bon nom d’espèce, sans erreur. Elle ne dit rien sur le monde, et donc ne créé ni ne justifie aucun groupe, aucun concept. C’est le contraire de la classification, qui, elle, n’a pas pour vocation d’être pratique et qui restitue certaines propriétés du monde (selon un « cahier des charges » fixé à l’avance) à travers les groupes qu’elle créée. Confondre la détermination et la classification n’est pas une faute biologique, c’est une faute cognitive.
Remarque : certaines clés de détermination se contentent d’aboutir à de grands groupes du vivant. D’autres aboutissent à des noms d’espèces mais signalent, le long du cheminement, des noms de groupes, fonctionnant comme des « balises ». Sur telle branche de l’arbre décisionnel, on indique qu’on entre dans les vertébrés, dans les rosacées, etc. Ces balises ont certainement contribué à entretenir la confusion entre clé et classification.
Classer : faire des ensembles argumentés et les nommer
Classer, c’est regrouper des objets dans un ensemble, faire un lien entre ces objets qui fait sens. Cet ensemble est un concept. Il est argumenté : on met ensemble des objets parce que certaines de leurs propriétés sont partagées par tous (Fig. 3). On ne fait pas un ensemble sur une absence d’attribut, puisque la liste de ce qu’un ou plusieurs objet(s) n’a pas est infinie. Si on s’autorise cela, on peut toujours faire des ensembles absurdes : je mets un train, une feuille de papier et un nuage dans un groupe, et je peux toujours le justifier en disant qu’ils n’ont pas de chaussettes. C’est ce qui fait la différence entre le tri et la classification : on peut séparer sur un critère de présence/absence, mais on ne peut pas regrouper sur la base d’une absence. Ensuite il faut nommer ce concept.
Prenons par exemple le concept de « fruits de mer ». Dans la nature il n’y a pas de fruits de mer. Il n’y a que des individus. « Fruits de mer » est un concept très sophistiqué. C’est un concept culinaire. On a mis dans un ensemble, sous ce nom, une cinquantaine d’espèces parce qu’elles constituent une palette gustative homogène. On ne choisit d’ailleurs pas n’importe quel vin avec un plateau de fruits de mer.
Dans la nature il n’y a pas d’oiseaux. Il n’y a que des individus. « Oiseaux » est un concept très sophistiqué. C’est un concept phylogénétique. On a mis dans un ensemble plus de 10 000 espèces parce qu’elles portaient toutes des plumes, un bréchet, une fourchette, un bec, un hallux retourné, un pygostyle. Tous ces traits ont été acquis par voie d’ascendance commune, ce qui fait que le concept « oiseau » est phylogénétique.
La classification construit des ensembles - des concepts - emboîtés les uns dans les autres. Au sein des oiseaux, il y a les psittaciformes, les falconiformes, etc.
Encore une fois, la classification n’a pas pour vocation d’être pratique. Elle nous parle du monde qui existe, selon un « cahier des charges » qui a été fixé, et qui édicte la nature des concepts qui sont créés. C’est tout le contraire de la clé de détermination, qui, elle, a pour vocation d’être pratique et ne dit rien sur le monde. Il existe des naturalistes qui préfèrent les « anciennes classifications » parce qu’ils les trouvent « pratiques ». En fait, il ne s’agit pas de classifications aujourd’hui valides, mais d’un double mélange (Fig. 4), d’une part entre échelle des êtres et phylogénie, d’autre part d’un mélange de multiples cahiers des charges, linnéen typologistes (qui relève de concepts d’un autre temps), écologico-adaptatifs (qui relève de concepts différents) et phylogénétiques (qui relèvent de concepts modernes que les classificateurs ont décidé de créer). Ces figures émanaient d’une procédure où les classes linnéennes étaient prises comme des existants et l’on se mettait à chercher alors les « affinités évolutives » entre ces classes. Dans la procédure moderne, on cherche d’abord les affinités phylogénétiques entre les espèces et on les classe seulement ensuite.
On reproche parfois aux scientifiques de compliquer la classification, de faire des classifications « pas pratiques ». On exige parfois qu’une classification donnée soit pratique parce qu’on n’a pas compris que c’est elle qui créé les concepts. On n’entend personne dire que le nombre d’objets astronomiques ne rend pas « pratique » la compréhension de l’univers, ni qu’il n’est pas « pratique » d’avoir plus de 100 éléments dans la table de Mendeleïev. Le monde est ce qu’il est ; la science doit en rendre compte rationnellement et ne peut dans son exercice subir les contraintes de l’utilisation des connaissances. En revanche, dans un but pédagogique, on peut toujours simplifier une classification scientifique.
En outre, la figure mélange arbre phylogénétique et échelle des êtres
Toute classification est arbitraire. Ce qui compte, c’est qu’elle réussisse à remplir le cahier des charges qui lui a été fixé. De quelle réalité, de quelles propriétés des objets la classification va-t-elle rendre compte ? C’est ce que nous verrons dans la section 3.
Assigner : ranger
Supposons que nous ayons devant les yeux une série d’ensembles emboîtés, avec pour chacun des arguments justifiant leur regroupement. Supposons que nous ayons un objet dans les mains. Assigner consiste à replacer cet objet dans le bon ensemble, en observant si ses propriétés correspondent aux justifications données pour chaque ensemble (Figure 5).
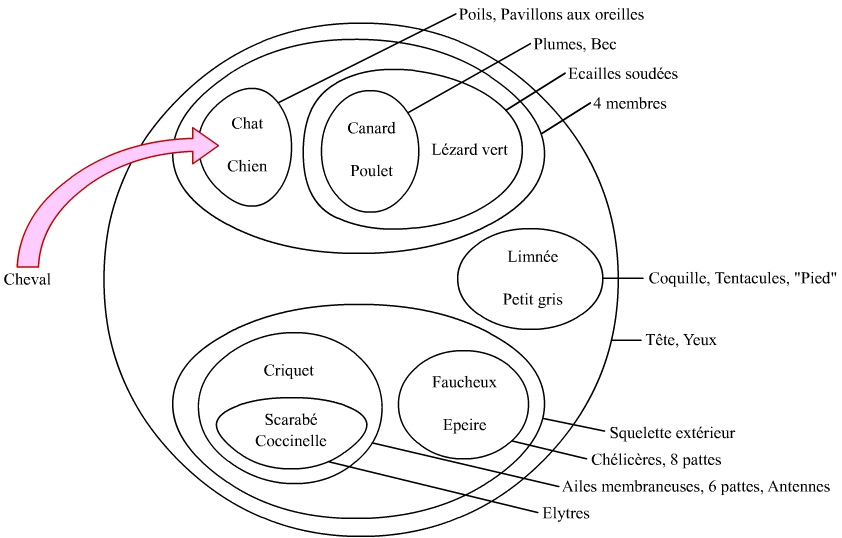
Les mathématiciens spécialisés dans la classification nomment cette activité « ranger ». Certains enseignants parlent de « placer », ce qui est juste également. Notons toutefois que pour certains enseignants, si trier était synonyme de discriminer des objets selon un critère discontinu, ranger consistait à discriminer selon un critère continu. Dans ce dernier cas, « ranger » deviendrait en quelque sorte synonyme de « ordonner ».
Quel cahier des charges ?
Classer pour parler des origines de ce qui existe
Nous avons vu que toute classification est arbitraire, et qu’une bonne classification est celle qui réussit à remplir le cahier des charges qui lui a été fixé. Dans une cuisine, nous regroupons des objets dans des ensembles conceptuels destinés à rendre compte de la palette gustative : fruits de mer, gibiers, volaille, poissons, fruits, légumes, etc. On ne choisit pas n’importe quel vin avec les aliments de chacun d’entre eux.
Même à l’intérieur des sciences biologiques, il existe des concepts classificatoires relevant de cahiers des charges différents. En écologie : phytoplancton, zooplancton, algues, super-prédateurs, fouisseurs, détritivores et consommateurs primaires sont des ensembles qui rendent compte des relations fonctionnelles qu’entretiennent leurs membres avec la biosphère. En systématique : diatomées, chaetognathes, rhodobiontes, chlorobiontes, falconiformes, phocidés, talpidés, lumbricidés, nématodes, ruminants, périssodactyles, mammifères et oiseaux sont des concepts phylogénétiques. Chaque ensemble contient des membres plus apparentés entre eux qu’ils ne le sont avec quoi que ce soit d’autre.
Certains assignent à la classification phylogénétique une sorte d’hégémonie, où le concept de « poissons » n’étant plus valide, il ne faudrait plus l’employer chez le poissonnier. Ainsi, certains pensent que toute entreprise de modernisation et de perfectionnement des savoirs en matière de classification est vain puisqu’on ne dira jamais « actinoptérygien » ou « téléostéens » chez le poissonnier. C’est se méprendre sur les intentions et les buts pédagogiques et conceptuels recherchés. Il ne s’agit pas de proscrire certains mots du vocabulaire. Il s’agit de savoir quels concepts employer dans quels registres, et donc de les employer à bon escient. Ainsi, on continuera à dire « poissons », « fruits de mer » dans une cuisine ou chez le poissonnier, et on dira « actinoptérygiens », « deutérostomiens » dans un laboratoire de zoologie ou de systématique.
Utiliser un concept dans le registre erroné conduira à des erreurs. Ainsi, « nuisibles», « bétail », « poissons », « invertébrés », « pachydermes », « palmipèdes » ne sont plus utiles en systématique puisqu’ils ne se conforment pas au cahier des charges de la systématique contemporaine, qui forge des ensembles pour parler des origines de ce qui existe, et donc fonde ces ensembles sur les relations phylogénétiques.
Remarque : on pourra parfois être surpris de trouver dans un musée d’anciens noms comme « poissons », « reptiles », « invertébrés », etc. Cela tient à de multiples raisons sociales et historiques -relevant de l’histoire des Hommes comme celle des sciences- qui seraient longues à exposer ici. Plusieurs musées en France ont déjà entamé une profonde restructuration de ce qu’ils présentent de cette discipline, la systématique.
On n’a pas toujours pensé la même chose sur les origines de ce qui existe
Le cahier des charges modernes des classifications en sciences naturelles est de rendre compte des origines de ce qui existe. On va mettre dans un ensemble des êtres vivants non pas par ce qu’ils font, non pas par la façon dont nous les utilisons, mais par leur communauté d’origine. Notons que ce cahier des charges est déjà très ancien. Les classifications « naturelles » des seizième, dix-septième et dix-huitième siècles entendaient restituer, dans une pensée créationniste fixiste, un ordre intrinsèque à la Nature qui n’était autre que celui voulu par le créateur à l’origine du monde. Karl von Linné (1707-1778) écrivit lui-même que la systématique était une « science divine » et qu’elle avait pour but, à travers la classification, de rendre intelligible le plan du créateur. Ce cahier des charges –parler des origines- n’a pas changé. Seulement, nos idées sur ce qui est à l’origine des espèces, elles, ont changé. En 1859, Charles Darwin (1809-1882) fixe explicitement le nouveau cahier des charges des classifications en sciences naturelles : comme les espèces ont évolué dans le passé, elles sont plus ou moins apparentées entre elles ; et c’est donc l’ascendance commune qui justifie les groupes fabriqués par la systématique, et non pas « quelque plan inconnu de création ». C’est par les attributs possédés en commun par des espèces qui ne se croisent plus aujourd’hui qu’on a les meilleures chances de détecter l’apparentement. Ce point d’histoire des sciences est, pour les enseignants, important à noter à deux égards.
Premièrement, il faut noter que les mots ont davantage d’inertie que les concepts, et que par conséquent nous utilisons encore des mots qui nous trompent. Par exemple, « invertébrés » (ceux qui n’ont pas les vertèbres), « agnathes » (ceux qui n’ont pas les mâchoires), et même « poissons » (ceux qui ne sont pas « sortis » de l’eau : ceux qui n’ont pas les pattes) ne nous parlent aucunement d’apparentement. Ce sont des groupes qui ont été forgés à une époque où l’Homme était au centre du monde. Ils ne prennent sens que parce que leurs membres ne possèdent pas quelque chose que l’homme a. Or, on ne parle pas de l’escargot ou du ver de terre quand on dit qu’ils n’ont pas de vertèbres. Ces groupes ne parlent pas d’apparentement car seuls les attributs présents peuvent témoigner de l’ascendance commune. On n’hérite que de ce qu’on a. La liste de ce que l’on n’a pas est infinie, et ne peut donc servir à circonscrire quelque ensemble que ce soit. Ces groupes centrés sur l’homme ou, plus généralement, tout groupe prenant référence à l’extérieur de lui-même, sur un autre groupe (procaryotes, protozoaires, invertébrés, agnathes, poissons, reptiles, prosimiens, aptérygotes, etc.), ne nous parle pas d’apparentement. Certains poissons (coelacanthe, dipneustes) sont plus apparentés aux tétrapodes qu’ils ne le sont des autres poissons. Certains invertébrés (échinodermes, urochordés…) sont plus apparentés aux vertébrés qu’à leurs confrères au sein des invertébrés. Certains reptiles sont plus apparentés aux oiseaux (les crocodiles) ou aux mammifères (les pélycosaures) qu’ils ne le sont des autres reptiles. Les enseignants doivent non seulement faire face à des mots qui viennent de registres non scientifiques (« fruits de mer », « nuisibles », « bétail »), mais aussi à l’héritage culturel reflétant le passé des sciences mais pas la science actuelle (« poissons », « invertébrés », etc.).
Deuxièmement, il a fallu attendre un siècle après Darwin pour que les scientifiques trouvent les moyens de réaliser son cahier des charges. Entre temps, on a multiplié le cahier des charges de la classification « naturelle ». La systématique qui s’est appelée elle-même « éclectique » voulut parler non seulement des liens d’apparentement, mais aussi d’écologie, d’adaptations « majeures », des « grandes transitions », de « complexité croissante ». Ces différentes propriétés se contredisant parfois entre elles quant à leurs résultats classificatoires, et par manque de cahier des charges univoque, ces méthodes ne réussirent qu’à pérenniser une bonne part des vieux cadres linnéens en les rhabillant d’évolution tout au long des deux premiers tiers du vingtième siècle. Ceci explique le retard pris par l’enseignement, et au premier chef ceux des premier et second cycles universitaires.
Le cahier des charges moderne : construire une phylogénie avant de classer
Introduction
Pour les enseignants, la façon d’aborder la classification pourra être différente selon si l’évolution est connue comme fait, mieux comme théorie, ou non. Pour les professionnels de la classification, aujourd’hui, une bonne classification repose sur une phylogénie préalable. Dans cette section on va donc voir comment ils construisent une phylogénie. Mais pour aborder la classification à l’école primaire, et même dans le secondaire, on peut classer (c’est-à-dire tracer des ensembles) sans passer par la phylogénie, en travaillant sur des collections d’espèces contrôlées. La raison en est qu'en cycle 2 à l'école élémentaire, l'évolution n'est pas au programme alors que l'élève aborde les bribes de la classification. Le but est non pas d’être capable de classer tout ce qui vient, mais de comprendre que la mise en ensemble se fait sur la base d’arguments, et de comprendre le rapport entre les mots et les choses. C’est dans le chapitre 5 que ces aspects seront abordés.
Comment construire une phylogénie (pour les professionnels) : l’homologie
L’homologie par comparaison
En systématique, on pratique la biologie comparée qui consiste à détecter la ressemblance entre les structures présentées par un échantillon d’organismes, en les comparant. On utilise une grille de lecture théorique commune à tous les organismes de notre échantillon, qui va permettre de détecter les ressemblances et les différences entre organismes. Selon une définition vieille de 150 ans, sont homologues des structures qui, prises chez des organismes différents, entretiennent avec les structures voisines les mêmes connexions et les mêmes relations topologiques, et ceci quelles que soient leur forme et leur fonction.
Prenons l’exemple classique du membre antérieur des tétrapodes. Le squelette du membre antérieur du dauphin, de la chauve-souris et de l’homme ne sont pas identiques, ne fonctionnent pas de la même manière, mais il est pourtant possible de détecter un agencement commun de leurs différentes pièces constituantes, symbolisé selon le plan d’organisation, grille de lecture théorique. Le radius du dauphin est homologue du radius de la chauve-souris, en dépit de leurs formes et tailles respectives très dissemblables : dans les deux cas ils sont connectés à un segment proximal unique lui-même connecté à la ceinture scapulaire (l’humérus). Ils sont aussi connectés à un second os parallèle (ulna), puis à des pièces carpiennes plus distales. Si, de plus, de par la forme du radius, nous pouvons mettre en évidence que le radius de la chauve-souris ressemble plus à celui de l’homme qu’à celui du dauphin, on peut émettre une hypothèse d’homologie (ou homologie primaire), c’est-à-dire qu’on fait le pari qu’à cette ressemblance anatomique correspond une communauté d’ascendance. C’est l’arbre phylogénétique qui, une fois reconstruit, donnera la réponse au pari. Il dira si les structures supposées homologues sont bel et bien héritées d’un ancêtre commun hypothétique (homologie secondaire, ou homologie confirmée), ou bien si elles sont apparues plusieurs fois indépendamment. L’homologie primaire est une hypothèse d’homologie à partir des connexions. L’homologie secondaire, c’est l’homologie par ascendance commune, confirmée par l’arbre.
L’homologie par l’embryologie
L’ontogenèse nous révèle également l’origine des organes. Il existe un parallèle entre le développement embryonnaire et le déroulement phylogénétique, remarqué depuis le XIXe siècle. Si l’ontogenèse récapitule la phylogenèse, à une communauté d’origine embryologique doit correspondre, en première approximation, une communauté d’origine phylogénétique. L’embryologie peut donc également conduire à formuler des hypothèses d’homologie primaire. Par exemple, une mandibule de lézard est constituée de plusieurs os, notamment le dentaire, l’articulaire, l’angulaire ; cette mandibule s’articulant au crâne au niveau de l’os carré. Or, chez un mammifère, la mandibule est formée du seul os dentaire, lequel s’articule au crâne au niveau d’un autre os, le squamosal. L’articulaire, l’angulaire et l’os carré sont passés dans l’oreille. En effet, deux lignes d’argumentation se corroborent très fortement pour établir des homologies entre des os de la mandibule reptilienne et des os de l’oreille mammalienne. L’une est paléontologique, l’autre est embryologique. Toute une série de fossiles du Permien et du Trias montrent une position de plus en plus postérieure de l’articulaire et de l’angulaire, jusqu’à leur intégration à la sphère auditive tandis que l’articulation de la mandibule au crâne devient dento-squamosale. L’embryologie des marsupiaux montre le détachement de pièces cartilagineuses de la mandibule vers la sphère auditive. Ces données établissent que l’angulaire de la mâchoire d’un lézard est l’homologue de l’os tympanique des mammifères, que l’articulaire du lézard est l’homologue au marteau, l’un des trois osselets de l’oreille moyenne mammalienne conduisant les vibrations du tympan vers la fenêtre ovale, et que l’os carré du lézard est l’homologue de l’enclume, le second des trois osselets de l’oreille moyenne des mammifères. Sans l’embryologie ni les fossiles, il eût été fort difficile d’établir une hypothèse d’homologie, c’est-à-dire de communauté d’origine, entre des os mandibulaires et des os de l’oreille moyenne.
Les progrès du XXe siècle
On ne classe jamais qu’un échantillon
De nos jours, la classification naturelle unique que réalisent les chercheurs est la classification phylogénétique du vivant. Celle-ci a été rendue possible à partir de la Systématique phylogénétique de Willi Hennig, publiée en allemand en 1950, mieux diffusée à partir de sa traduction anglaise en 1966. Pour comprendre Hennig, il faut commencer par réaliser qu’on se propose de classer un échantillon du vivant, et jamais tout le vivant d’un coup. Cette remarque semble anodine mais elle est capitale pour garantir transparence et reproductibilité des procédures, ainsi qu’un véritable contrôle du principe d’économie d’hypothèses. Cet échantillon, on le veut « représentatif » d’un problème posé. La valeur et la signification des attributs des organismes (les caractères) peuvent ne pas être les mêmes d’un échantillon à l’autre ; et cela a une grande importance pédagogique. Par exemple, le fait d’avoir quatre pattes ne sert à aucun regroupement au sein des mammifères, tandis qu’au sein des vertébrés, la présence des quatre pattes est une innovation évolutive qui signe l’apparentement des tétrapodes.
Les innovations évolutives de l’échantillon sont les marqueurs de l’apparentement exclusif
On se propose donc toujours de retrouver les relations de parenté au sein d’un nombre limité d’espèces, un échantillon, par exemple un lézard, un serpent, un crocodile, un poulet, un canard et une pie. Cet échantillon possède toujours, dans l’arbre phylogénétique de la vie, un ancêtre commun à tous les représentants choisis. Ceci reste vrai même si on ne connaît pas l’ordre de leurs branches : il suffit de redescendre jusqu’à ce qu’on parvienne à la première branche commune. Appelons cet ancêtre abstrait X. Il s’agit d’enregistrer des caractères, ou attributs de ces organismes, pour lesquels on peut distinguer au moins deux états au sein de l’échantillon. Ces états sont, en première approximation, supposés homologues (voir paragraphe précédent). Par exemple, le bourgeon épidermique donne de l’écaille chez certains de ces organismes, et de la plume chez d’autres. Ecaille et plume sont homologues a priori en tant que dérivés de l’épiderme dont les bourgeons se ressemblent.
Une classification « classique » conduirait, pour ce seul caractère intitulé « bourgeon épidermique », à créer un groupe lézard + serpent + crocodile d’un côté, avec comme argument le partage des écailles, et de l’autre un groupe poulet + canard + pie fondé sur le partage de la plume. Pour Hennig, ce n’est pas la bonne manière de procéder. Si nous émettons une hypothèse d’homologie entre écaille et plume, et si nous assumons pleinement l’évolutionnisme dans la manière de construire une classification, alors l’un des deux états est dérivé de l’autre. Pour l’échantillon d’espèces, cela signifie que l’un des deux états a été acquis depuis X (l’état dérivé), et l’autre a été acquis dans l’arbre avant X (l’état primitif). Celui des deux états acquis avant X ne peut pas servir à faire des regroupements au sein de notre échantillon, puisqu’il est déjà présent en dehors, sur d’autres branches de l’arbre de la vie. Si nous faisions un groupe sur le partage d’un tel état, notre groupe serait incomplet, oubliant tous ceux qui sont à l’extérieur. Par contre, il faut faire des regroupements sur la présence commune d’états de caractères dérivés. Ils constituent des innovations évolutives dans l’échantillon, et elles seules peuvent fournir des apparentements exclusifs et donc des groupes complets.
Polariser les caractères par l’ontogénie
Déterminer l’état primitif, cela se nomme polariser le(s) caractère(s). Comment sait-on lequel des deux états est dérivé de l’autre ? Il existe deux critères utilisés actuellement. Le critère ontogénétique et le critère de l’extra-groupe.
Le critère ontogénénétique utilise le fait que l’ontogenèse récapitule la phylogenèse. En première approximation, le développement embryonnaire évoluant dans le temps phylogénétique par additions successives de séquences nouvelles de développement, les états de caractères d’apparition précoce dans le développement sont distribués plus généralement dans le vivant que ne le sont les états de caractères d’apparition plus tardive. Par exemple, dans le déroulement phylogénétique, nous avons été vertébrés (il y a 500 m.a.) avant d’être tétrapodes (il y a 380 m.a.), avant d’être des primates hominoïdes (il y a 25 m.a.). Dans le temps embryologique, la colonne vertébrale apparaît avant les membres, qui apparaissent eux-mêmes avant la régression de la queue qui caractérise les primates hominoïdes. Celui des deux états qui donne naissance à l’autre est donc l’état primitif, celui qui en découle est l’état dérivé. Le bourgeon épidermique préfigure déjà l’écaille mais se transforme en plume. On peut même, expérimentalement, transformer les écailles des tarses des poulets en plumes en ayant injecté de l’acide rétinoïque dans l’embryon à un certain stade de son développement. Tout semble donc indiquer que la plume est une écaille dérivée. Dans ce cas, le groupe que l’on va constituer est celui des espèces de l’échantillon possédant l’état dérivé, à savoir la plume, donc le groupe poulet + canard + pie.
Polariser les caractères par comparaison avec l’extra-groupe
Le critère extra-groupe consiste à prendre un référentiel, à aller voir ce qui se passe à l’extérieur de l’échantillon que l’on se propose de classer. Aucune construction scientifique ne se fait sans postulats. Une classification n’échappe pas à cette règle, mais il faut que les postulats soient explicites. On énonce alors clairement le postulat suivant : la tortue est extérieure à l’échantillon lézard + serpent + crocodile + poulet + canard + pie. Ce postulat fait de la tortue un extra-groupe. Observons à présent l’épiderme de la tortue. On y trouve de l’écaille. C’est donc que l’écaille était, dans l’arbre de la vie, déjà présente avant X. L’écaille est l’état primitif, la plume est l’état dérivé. Par conséquent, nous allons créer un groupe au sein de l’échantillon, qui signera l’apparentement exclusif poulet + canard + pie, « exclusif » signifiant à l’exclusion du lézard, du serpent et du crocodile.
Assumer ses postulats
Il convient de faire trois remarques au sujet de l’extra-groupe. La première, c’est qu’il est généralement recommandé d’utiliser plusieurs extra-groupes de manière à bien argumenter la polarisation des caractères. La seconde, c’est que si nous nous étions trompés d’extra-groupe en utilisant, par exemple, une autruche, nous aurions posé la plume comme état primitif. Le groupe créé aurait été lézard + serpent + crocodile. Cela signifie que la procédure en elle-même ne gère pas la pertinence des postulats. Comme partout en sciences, la pertinence des postulats ne relève que de la fiabilité des connaissances extérieures à l’expérience, lesquelles doivent être assumées par le chercheur. Si l’un des postulats est faux, le résultat a peu de chances d’être vrai, mais alors ce n’est pas la procédure qui est en cause. Enfin, il est clair qu’on ne fonde pas une classification sur un seul caractère, mais qu’on en utilise plusieurs simultanément. Cela nous conduit à l’exercice suivant.
La procédure standard
Poser une question, envisager toutes les réponses possibles
Après l’exposé des principes, il convient à présent d’exposer la procédure élémentaire qui permet aujourd’hui de construire une classification. Dans un souci de simplicité et de place, nous proposons de classer un nombre minimal d’espèces. Rappelons-nous que la question fondamentale est « qui est plus proche de qui ? ». A deux espèces, cela n’a pas de sens. On peut répondre à partir de trois espèces. Avec trois espèces, deux d’entre les trois peuvent être plus proches entre elles que chacune ne l’est de la troisième. Enfin, si nous tenons compte du paragraphe précédent, il faut un référent extérieur pour polariser les caractères : l’extra-groupe. Le minimum d’espèces à considérer est donc de quatre. Prenons par exemple une chauve-souris, un homme, un pigeon. L’ancêtre commun à tous dans l’arbre de la vie (l’ancêtre X ci-dessus) est l’ancêtre commun hypothétique à tous les amniotes. L’extra-groupe sera donc pris en dehors des amniotes, par exemple une truite. La question est donc : parmi la chauve-souris, l’homme et le pigeon, quels sont les deux des trois les plus apparentés ? En laissant toujours la truite à l’extérieur, ce qui fait partie des postulats, il y a trois réponses possibles, exprimables ci-dessous sous forme de parenthèses, mais aussi sous forme d’arbres (Figure 6) :
((chauve-souris, homme) pigeon) : b
((chauve-souris, pigeon) homme) : c
((pigeon, homme), chauve-souris) : d
La procédure moderne de construction d’une classification s’emploie donc à explorer un champ d’hypothèses possibles.
Coder des caractères
Il faut à présent explorer des caractères pour évaluer l’argumentation de chacune d’elles. Les observations faites sur les différents états des caractères vont être codés à l’aide de chiffres et inscrites dans un tableau appelé matrice de caractères (fig. 6A). Par convention d’écriture, l’état trouvé dans l’extra-groupe est noté « 0 ». Attention, le zéro ne signifie pas une absence de caractères, mais simplement l’état dans lequel il est dans l’extra-groupe. Nous pouvons proposer :
1. Sacs aériens : sous forme de vessie natatoire (0) ; sous forme de poumons alvéolés fonctionnels (1).
2. Appendices pairs : sous forme de nageoires rayonnées (0) ; sous forme de membres marcheurs (1)
3. Constitution de la mandibule : plusieurs os (0) ; un seul os, le dentaire (1)
4. Glandes mammaires : absence (0) ; présence (1)
5. Ailes : absence (0) ; présence (1)
6. Dents : présence (0) ; absence (1)
7. Colonne vertébrale : présence (0) ; absence (1).
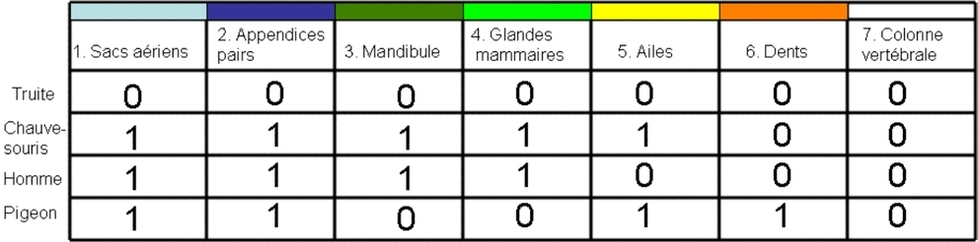
Compte tenu de la question posée ci-dessus, on constate d’emblée que certains caractères ne servent à rien. Le fait de posséder une colonne vertébrale (caractère 7) ne discrimine nullement deux d’entre les trois. C’est la même chose pour les caractères 1 et 2. La chauve-souris, l’homme et le pigeon possèdent tous les trois les caractères 1 , 2 et 7 sous le même état. On dit que ces caractères ne sont pas informatifs au regard de la question posée. Ceci valait d’être souligné, parce que c’est de cette manière que les chercheurs sélectionnent leurs caractères. Dans le gisement de caractères que constitue une collection d’organismes, ils ne prélèvent que les caractères potentiellement informatifs. Cependant, parmi les caractères 3, 4, 5 et 6, il en reste un qui ne pourra servir à établir un regroupement. C’est le caractère 6. En effet, il semble discriminant puisqu’il conduit à une ressemblance commune à l’homme et à la chauve-souris qui tous deux possèdent des dents. Mais sachant que la présence de dents est déjà observée dans l’extra-groupe, on en déduit qu’il s’agit ici un état primitif partagé et l’absence de dents un état dérivé. On se souvient de la règle de Hennig qui consiste à ne pas établir de groupes sur la base de caractères primitifs partagés mais seulement sur la base d’un état dérivé partagé. L’état dérivé étant l’absence de dents, le caractère 6 ne fait donc que regrouper le pigeon avec lui-même. En fait, du point de vue algorithmique, un caractère informatif est celui qui présente au moins deux états (ce qui n’est pas le cas pour le 7), chacun étant représenté au moins deux fois (ce qui n’est pas le cas des caractères 1, 2, 6). Les seuls caractères informatifs sont donc les caractères 3, 4 et 5.
Placer les transformations des caractères sur les arbres
On constate que les caractères se contredisent entre eux. En effet, les caractères 3 et 4 montrent les états dérivés communs à l’homme et à la chauve-souris, et le caractère 5 montre un état dérivé commun la chauve-souris et le pigeon. Comment s’en sortir ? En observant deux règles : la première, en plaçant dans chacun des arbres possibles les transformations des caractères, (Fig. 6B, bcd), la seconde , en respectant le principe d’économie d’hypothèses. Par exemple, (Fig. 6B, arbre b) on va tenter de placer l’endroit dans l’arbre où le caractère 3 change d’état, c’est-à-dire passe d’une mandibule à plusieurs os (état « 0 » trouvé chez la truite) à une mandibule constituée du seul os dentaire (état « 1 »). Ce passage deviendra une hypothèse de transformation (symbolisé par un petit cercle de couleur Figure 6B, bcd). Cette action doit respecter le principe d’économie d’hypothèses, selon lequel on ne va pas multiplier les transformations au-delà du simple nécessaire. Par exemple, dans l’arbre b, on pourrait vouloir faire apparaître la mandibule constituée du seul os dentaire une fois sur la branche menant à l’homme, et une autre fois sur la branche propre à la chauve-souris. Cela respecterait les observations. Mais on peut faire « moins cher » : en plaçant plus simplement cet événement au segment de branche commun à l’homme et à la chauve-souris, réalisant ainsi une seule transformation au lieu de deux. En réalisant cet exercice sur tous les caractères, on observera que pour certains caractères il n’est pas possible de faire moins de deux transformations. Dans l’arbre b, nous sommes forcés de faire apparaître les ailes deux fois, car il n’y a pas de branche menant exclusivement au couple chauve-souris + pigeon : une fois sur la branche propre à la chauve-souris, et une fois sur la branche propre au pigeon. Il faut répéter cet exercice sur chacun des trois arbres possibles, de telle manière qu’à la fin de cet exercice, tous les caractères ont été interprétés en termes de transformations évolutives dans chacun des arbres possibles.
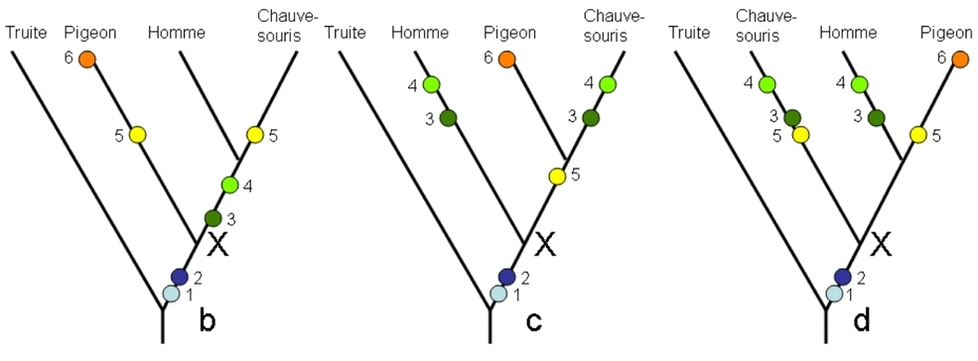
Appliquer le principe de parcimonie
Le principe d’économie d’hypothèses, utilisé dans toutes les sciences, va conduire à retenir l’arbre b, parce qu’il ne « coûte » globalement que 7 hypothèses de transformations, tandis que les deux autres arbres possibles sont moins parcimonieux (8 hypothèses pour c et 9 hypothèses pour d). Bien entendu, les chercheurs travaillent sur beaucoup plus d’espèces et plus de caractères. Ils utilisent alors des algorithmes qui réalisent la procédure exposée ici, à l’aide d’ordinateurs. Ceux-ci sont en fait indispensables, car le nombre d’arbres possibles à explorer augmente très rapidement avec le nombre d’espèces à classer. Remarquons que le principe de parcimonie est un principe de cohérence. L’arbre b montre la contigüité (la proximité) des états de caractères dérivés pour les caractères 1, 2, 3, 4, tandis que l’arbre c ne le fait que pour les caractères 1, 2, 5 et l’arbre d seulement pour les caractères 1, 2. L’arbre b est donc le plus cohérent. En général, en science, nous sommes parcimonieux parce que nous cherchons la cohérence interne de nos systèmes d’explication.
L’arbre final, clé de la classification, clé des homologies
Deux groupes-frères ont le même rang
L’arbre retenu est celui qui est le plus parcimonieux. Il fournit une classification, et en même temps les réponses aux paris sur les homologies. Dans l’arbre b, on dit que l’homme et la chauve-souris sont groupes-frères. Deux groupes-frères ont le même rang. Ainsi, dans l’arbre phylogénétique des vertébrés, les chondrichthyens (vertébrés cartilagineux: requins, raies) et les ostéichthyens (vertébrés osseux ) sont groupes-frères : alors chondrichthyens et ostéichthyens sont de même rang (par convention, on décidera qu’il s’agira de deux classes).
Nommer les clades
Clade est synonyme de « groupe monophylétique ». Homme et chauve-souris sont tous deux sont inclus dans un clade. On peut donner un nom à chaque clade. Dans l’exemple simplifié qui est le nôtre, ce nom pourrait être « Mammifères ». Pigeon, homme et chauve-souris sont inclus dans un clade plus inclusif, comprenant le premier, et donc de rang supérieur. S’il fallait le nommer, il s’agirait ici des Amniotes. En fait, dans tout arbre phylogénétique, chaque segment de branche interne correspond à un groupe nommé qui englobe tout ce qui est en aval de cette branche (voir figure 1, mais surtout Lecointre et Le Guyader, 2017). Si nous sommes capables de construire des phylogénies et de suivre cette règle, tous les groupes ainsi créés seront monophylétiques, et la classification qui en découlera sera réellement phylogénétique, comme l’avait souhaité Darwin. Dans le cahier des charges des classifications actuelles, l’arbre phylogénétique guide donc, voire fournit la classification.
L’arbre retenu nous enseigne quels attributs sont hérités d’un ancêtre commun exclusif
Il y a trois types de ressemblance.
La première est celle due à un caractère primitif partagé, trop général pour pouvoir opérer une classification au sein de l’échantillon présent. Par exemple, le fait d’avoir des vertèbres font se ressembler nos trois espèces, mais cela ne nous dit rien sur « qui est plus proche de qui ? ».
La seconde est due à des caractères dérivés partagés. Ces ressemblances sont celles qui permettent de créer des classifications phylogénétiques. Nous avions fait le pari (« homologie primaire ») que la mandibule faite du seul os dentaire et les mamelles avaient été acquises d’un ancêtre commun. L’arbre le plus parcimonieux nous enseigne que nous avions eu raison. Ces ressemblances-là sont des homologies confirmées, ou « homologies secondaires ».
La troisième est la ressemblance qui correspond à des caractères dérivés non obtenus par ascendance commune. En effet, au cours du déroulement évolutif, des caractères qui se ressemblent beaucoup peuvent être acquis plusieurs fois indépendamment dans des lignées différentes. Par exemple, les mammifères « ont fait de la taupe » deux fois : une fois en Australie (la taupe marsupiale) et une fois en Eurasie (la taupe placentaire), sans que les ancêtres respectifs de ces animaux n’aient eu une forme de taupe. Ces convergences évolutives sont dues au fait qu’une bonne partie des contraintes physiques que subissent les espèces sont les mêmes partout. Ainsi, les ailes de notre échantillon ne sont pas homologues en tant qu’ailes : on a appris à voler une fois chez les chauves-souris et une autre fois chez les oiseaux. Ces phénomènes expliquent pourquoi, dans une matrice de caractères, les caractères se contredisent les uns les autres : nous avons fait des paris sur l’homologie des caractères, mais pour certains d’entre eux nous nous sommes trompés, sans pouvoir détecter notre erreur a priori. Cela se produit toujours : il n’existe pas de matrice de caractères qui ne contienne pas de la contradiction interne. Cette contradiction entre caractères, on l’appelle homoplasie, ou incohérence de la matrice. Un arbre donné possède une certaine quantité d’homoplasies, ou ressemblances non héritées d’un ancêtre commun. Ici, l’arbre b présente une homoplasie, l’aile.
Ces trois types de ressemblance, directement tirés de la pensée de Willi Hennig, ont une importance pédagogique considérable pour la préparation de séances de classes. En particulier, l’enseignant pourra gérer l’homoplasie en choisissant l’échantillon d’espèces à classer de telle manière à contrôler l’introduction de ressemblances trompeuses.
L’analyse cladistique, ou encore la construction d’une classification phylogénétique (au sens de Hennig, 1950 et 1966), permet de prendre conscience qu’une classification n’est plus un art qui enferme son praticien dans l’autorité de son expertise, mais une démarche scientifique qui requiert des postulats, des observations formalisées, transparentes à tout autre classificateur ou utilisateur, des règles du jeu parmi lesquelles l’économie d’hypothèses, des décisions explicites, et surtout, l’exploration d’un champ des possibles (les différents arbres possibles). En somme, la classification phylogénétique, par l’explicitation qu’elle force, est capable de produire de la connaissance objective, qui est le propre des connaissances scientifiques.
Les bouleversements de nos classifications
Les classifications biologiques ont plus changé en 30 ans qu’elles n’ont changé auparavant. Les moteurs de ces changements sont au nombre de deux.
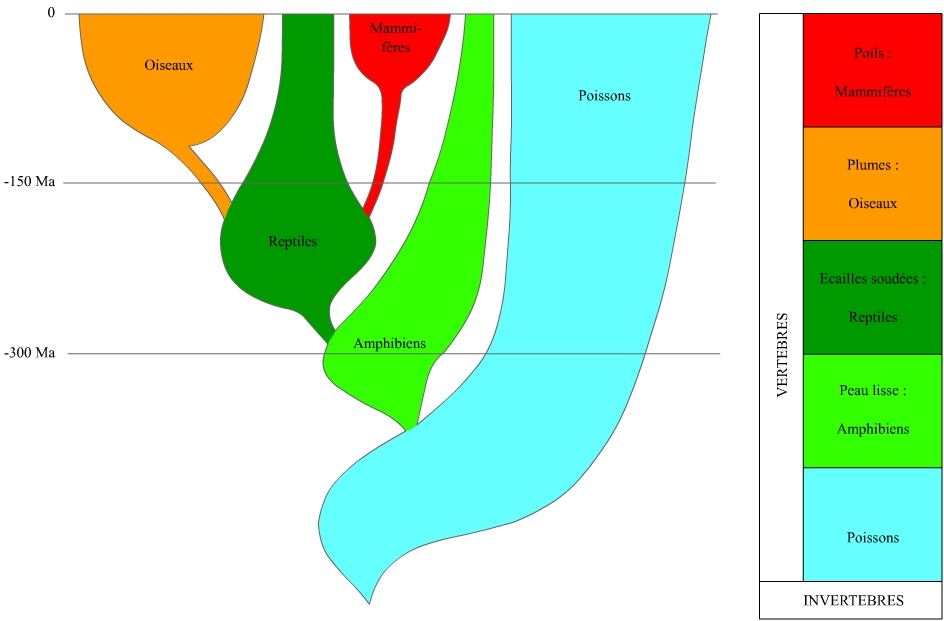
D’abord, il y a la mise en pratique généralisée et informatisée des idées de Hennig. C’est là le moteur principal. Des groupes ne sont plus valides dans les laboratoires, comme procaryotes, protistes, algues, ptéridophytes, gymnospermes, invertébrés, diploblastiques, acoelomates, pseudocoelomates, agnathes, poissons, reptiles, tandis que beaucoup d’autres ont été confirmés (eucaryotes, angiospermes, protostomiens, deutérostomiens, chondrichthyens, actinoptérygiens, oiseaux, mammifères…). Il faut y voir un progrès inestimable dans la cohérence et la précision de la perception que nous avons du monde vivant.
Le second moteur est l’accès à des classes de caractères auxquelles nous n’avions pas du tout accès avant les années soixante, et, pour des raisons techniques, très difficilement accès avant le dernier quart du vingtième siècle. Ces caractères sont les séquences d’ADN et de protéines, qui permettent de comparer des êtres vivants préalablement incomparables par leur seule morphologie, et ainsi permettent de défricher des pans entiers de l’arbre de la vie. En comparant les séquences d’un gène à évolution lente comme ceux des histones, chez une pâquerette, une vache, un champignon et des bactéries, on peut en produire une classification phylogénétique, ce qui était très difficile avant l’accès aux données moléculaires. Ainsi, ces trente dernières années, un certain nombre d’organismes se sont avérés ne pas être ce que l’on croyait. La reconstruction phylogénétique fait de la systématique une science, réalise le rêve de Darwin et bouleverse la perception et la connaissance que nous avons du monde vivant
Comment faire de la classification sans passer par l'évolution ni par la phylogénie ?
Objectifs généraux
Si l’on n’enseigne jamais que la « mise en ensemble » des espèces doit être argumentée, et que le nom qui est ensuite posé sur l’ensemble est une convention de langage (nominalisme), rien n’empêche les enfants ou les adultes de continuer à croire que les noms sont co-substantiels aux choses, ou, dit autrement, que les choses sont des instanciations d’entités idéales, dont elles portent l’essence (essentialisme). Il n’est pas rare de relever des phrases du type : « il a six pattes parce que c’est un insecte ». Toute classification génère des concepts, ce sont des ensembles argumentés sur lesquels, à terme, les scientifiques posent un nom. Si l’on comprend cela, on aboutit à : « Ils sont ensemble parce qu’ils ont tous six pattes ; les scientifiques appellent cela : insectes ». Le nom d’insecte n’émane pas de l’individu donné à observer.
Les objectifs de la classification dans le primaire et dans le secondaire sont donc beaucoup plus cognitifs que biologiques :
- Distinguer la classification de la clé de détermination
- Passer, en matière de classification, de l’essentialisme au nominalisme :
- Observer une collection contrôlée d’espèces, voire les dessiner
- Nommer les attributs observables
- Faire des ensembles en les argumentant sur la base d’attributs partagés
- Faire des ensembles emboîtés (comprendre que c’est nous qui classons ; cela ne provient pas des organismes classés : « ce n'est pas écrit dessus »)
- Comprendre que les noms qui viennent ensuite dessus sont des conventions créées par des scientifiques
Les objectifs ne sont pas (en tout cas pas dans un premier temps) :
- Faire de la phylogénie
- Pouvoir classer n’importe quel échantillon de terrain
- Connaître la liste exhaustive des groupes de la classification
Des activités en primaire qui peuvent êtres réinvesties dans le secondaire
En donnant une collection d’espèces à classer par les élèves, on peut atteindre un certain nombre d’objectifs conceptuels, dont le premier est de classer sur la base de ce que les organismes ont, et non sur ce qu’ils n’ont pas, sur ce qu’ils font, là où ils vivent, ce à quoi ils servent ou encore sur des a priori concernant ce qu’ils sont. On évacuera ainsi d’une classification qui se veut scientifique les classifications utilitaire, anthropocentrique, environnementale. On évitera aussi de confondre classer et trier, c’est-à-dire classification et clé de détermination, confusion sous-jacente à la distinction vertébrés-invertébrés. Seule une classification fondée sur ce que les organismes ont a des chances d’être phylogénétique.
Au niveau de l’école primaire, pour garantir un résultat le plus proche possible de réelles classifications phylogénétiques, l’échantillon d’espèces à classer et les critères (les arguments) devront être contrôlés. En effet, pour un échantillon d’espèces il existe trois types de ressemblances.
Il existe des ressemblances qui constituent des innovations évolutives au sein de l’échantillon donné. Celles-là signent un apparentement. Au sein d’un échantillon de vertébrés, avoir des poils va signer l’apparentement des mammifères de l’échantillon
Il existe aussi des ressemblances obtenues par convergence évolutive, et dont les regroupements vont contredire les regroupements précédents. Toujours au sein du même échantillon, si celui-ci contient une chauve-souris, sur le partage des ailes on va pouvoir proposer un regroupement de l’oiseau et de la chauve-souris, lequel s’oppose au rattachement de celle-ci avec le chat sur la base des poils partagés. Il est clair que l’on va gérer les contradictions possibles par le choix des espèces à classer (en fait, dans la réalité de la recherche, ces contradictions étaient gérées par le principe de parcimonie).
Enfin, il existe un troisième type de ressemblance, les caractères primitifs communs à tout l’échantillon. Si, dans un échantillon de vertébrés on dispose d’un Saint-Pierre, une truite et une perche, on va pouvoir faire remarquer que ces trois-là ont des branchies. Or, les branchies existent en dehors des vertébrés et en aucun cas ne serviraient classiquement à regrouper trois téléostéens entre eux. Comme il est impossible d’expliquer cela à l’école primaire (à moins de fixer un extra-groupe et d’y attacher une « règle du jeu »), on tentera de limiter la possibilité de tels arguments de classification par un choix méticuleux des espèces opéré en amont.
Les objectifs sont, sur plusieurs séances :
- Décrire les espèces : établir le niveau de description et faire émerger le vocabulaire anatomique
- Distinguer ranger, trier et classer
- Faire émerger des critères de classification pour, finalement, classer sur ce qu’ils ont
- Provoquer un débat où les observations faites et les savoirs seront confrontés
- Faire émerger des ensembles emboîtés
Note : au collège, une bonne partie de ces étapes est incluse dans un tableau d’observations (Figure ci-dessous).
Éventuellement (en primaire):
- Faire émerger la causalité sous-jacente au partage de ce qu’ils ont
- Dessiner un arbre à partir des ensembles
- Assigner des espèces surnuméraires, dont des fossiles, ou des espèces problématiques, dans la classification déjà constituée.
Observer et décrire
La première étape est un travail d’observation/description. Pour les enfants connaissant peu les êtres vivants, une première étape de description d’animaux et de végétaux à partir de photos est nécessaire afin de faire émerger des énoncés de description, puis, à terme une abstraction. En effet, on peut regarder une photo d’animal pour ce qu’il est en tant qu’individu, ou bien en tant que représentant d’un taxon déjà assimilé. Par exemple, on peut voir ce rouge-gorge là comme quelque chose d’inconnu à gorge rouge ou bien comme oiseau. Dans la description de ce que les animaux ont, on peut voir émerger que ce chat-là est roux, ou que cet autre chat a une pelote de laine. Pour permettre l’abstraction, une fois des descriptions obtenues, on peut tenter de trier les attributs généralisables des attributs qui résultent de situations particulières en :
- limitant les effets de taille
- limitant les couleurs par l’utilisation de photos ou de dessins en noir et blanc
- distinguant de qui est propre à la structure physique des organismes de la situation contingente de la photo
- multiplier les photos d’individus d’une même espèce pour ensuite, à terme, permettre de percevoir ce qui est propre à une espèce puis ôter les doublons
- on peut tenter de fournir un animal nouveau et de le rattacher à d’autres qu’on aura préalablement décrit.
À terme, en évacuant les tailles, les couleurs, les situations, on aura ajusté le niveau de description des espèces et fait émerger du vocabulaire anatomique (tête, yeux, ailes, poils, plumes, membres, pattes, nageoires, antennes…). L’étape suivante consistera à reproduire le même exercice sur un échantillon de composition contrôlée à classer.
Remarque : cette séance sera radicalement différente si l’on part d’un échantillon réel d’espèces collectées dans la nature. Cependant les objectifs sont les mêmes.
Distinguer trier, ranger, classer
Une collection d’espèces (Figure ci-dessous : criquet, coccinelle, hanneton, scarabée, truite, escargot petit-gris, limnée, saint-pierre, homme, lapin, chat, pigeon, poulet, lézard vert) est fournie sous forme de dessins photocopiés sur des feuilles de couleur. On demande aux élèves par petits groupes de trier ou classer les espèces selon la couleur de la feuille : ceux qui ont les feuilles roses devront trier, ceux qui ont les feuilles blanches devront classer. Avec des ciseaux et de la colle, chaque groupe réorganise les dessins en fonction de ce qu’il a à faire.
 |
 |
| Criquet | Coccinelle |
 |
 |
| Hanneton | Scarabé |
 |
 |
| Truite | Escargot petit gris |
 |
 |
| Limnée | Saint-Pierre |
 |
 |
| Homme | Lapin |
 |
 |
| Chat | Pigeon |
 |
 |
| Poulet | Lézard vert |
| Crédits photographiques : Michel Delarue (Criquet, Coccinelle, Scarabé, Lapin), Pauline Vonarx (Chat) et Gilles Camus (Pigeon), tous trois du service BioMédia de l'Université Paris 6; Pierre Davoust (Hanneton, Truite, Escargot, Limnée, Lézard vert), Université de Bretagne-Sud; site Marseille-Sympa (Saint-Pierre); site Kulturica (Homme); site Bresse Info (Poulet). |
|
Immédiatement, la notion de critère émerge. On va trier sur « avoir des poils » ou pas, sur « vivent à la ferme » ou pas.
On collecte les représentations, on les affiche toutes. On constatera que les deux opérations sont mélangées : certains ont trié en croyant classer et vice-versa. On peut alors redistribuer les feuilles en deux catégories :
Ceux qui ont trié :
Le tri se fait en fonction de la présence/absence du ou des critère(s). Par exemple, il y a ceux qui ont des poils (chat, lapin, homme) et ceux qui ne les ont pas (les autres). Cette activité est utilisée dans les clés de détermination d’espèces mais ne constitue en aucun cas une classification.
Ceux qui ont classé :
On regroupe sur la base de ce que les espèces ont (et pas sur ce qu’elles n’ont pas) et ces attributs sont appelés les arguments de la classification. Les arguments sont multiples et s’emboîtent les uns dans les autres. Par exemple, au sein de ceux qui présentent quatre pattes, il y a le sous-groupe de ceux qui ont des poils. Des ensembles emboîtés peuvent voir le jour.
Dans le secondaire, une séance particulière est prévue pour utiliser des clés de détermination préexistantes. On utilisera des clés sur les feuilles d’arbre ou sur les plantes à fleurs, plus faciles à gérer en identification qu’en véritable classification.
La classification, ses critères et ses arguments
En ayant laissé derrière soi les activités de tri, on peut passer à une réelle séance de classification. Dans un premier temps, on va laisser émerger les critères de classification, pour finalement aboutir à la consigne de ne classer que sur la base de ce que les animaux ont.
En effet, si on laisse émerger les critères de classification, on va obtenir un mélange de critères hétérogènes. Des animaux vont être classés ensemble parce que :
- ils font la même chose (« ils volent », ou « ils mangent de la viande »)
- ils vivent au même endroit (« ils habitent à la ferme »)
- ils sont (a priori) ceci ou cela (« ce sont des insectes »)
- ils servent à la même chose (« ils se mangent »)
- ils n’ont pas ceci ou cela (« ils n’ont pas de vertèbres », ou « ils n’ont pas de pattes »)
- ils ont ceci ou cela (« ils ont six pattes »)
En primaire, on pourra laisser émerger ces critères et l’on pourra les distinguer collectivement.
Si l’objectif est d’obtenir une classification qui a rapport avec l’histoire évolutive des organismes, la consigne devra être, finalement, de ne classer que sur ce que les animaux ont (le dernier des six ci-dessus). On peut y parvenir de manière directive, mais aussi par le dessin. Il suffit de se mettre à dessiner ce que l’on voit et à légender les parties. Lorsque l’on dessine, on s’attache à observer ce qu’il y a, et on laisse –de fait- tout le reste.
Pour le secondaire, il faut aller plus vite. Toutes ces étapes sont remplacées par un tableau à double entrée, donné à remplir, et qui permet de gagner du temps (Figure 8). Il suffira de mettre une croix là ou l’attribut nommé aura été observé.

Pour le primaire, une liste collective des organes (ou attributs) observés sur toute la collection est dressée. Un ajustement de vocabulaire est nécessaire (souvent, les « oreilles » sont données pour les « pavillons de l’oreille » ; préférer « membres » à « pattes » chez les tétrapodes ; attention à « ailes » ou à « nageoires » qui font référence à des fonctions, il faudra alors paire préciser les structures avec « ailes membraneuses », « ailes plumeuses », « nageoires rayonnées » ; préférer « tentacules » à « cornes » ou à « antennes » chez les gastéropodes ; le pied de l’escargot n’est pas son ventre, etc.). Un complément de documentation est toujours possible, puisque la séance a été préparée. Par exemple, l’homme a une queue, mais elle est « rentrée » dans le bassin, c’est le coccyx. De même, les oiseaux ont une queue, mais celle-ci est « tassée », c’est le pygostyle. Attention, les oiseaux aussi on des écailles soudées, il suffit d’observer leurs tarses. On différenciera les « écailles soudées » des sauropsides des « écailles libres » des téléostéens. Cette étape est déjà contrôlée dans le secondaire puisque le tableau utilise d’emblée le bon vocabulaire.
Puis on fabrique des ensembles argumentés à l’aide des attributs listés ci-dessus.
Spontanément la tendance sera de ne faire que des ensembles disjoints, non inclusifs. Une seconde consigne pourra encourager les jeunes classificateurs à faire surtout des ensembles emboîtés les uns dans les autres. Cela revient à hiérarchiser les arguments : tous ceux qui ont des poils avaient déjà quatre membres, etc. Une manière de démarrer l’emboîtement est de demander aux classificateurs ce que les animaux ont tous. Par exemple, ils ont tous une tête. Sur l’argument de la tête, le groupe le plus inclusif va pouvoir émerger. Les autres groupes seront ainsi nécessairement emboîtés dans le premier.
Dans l’option du tableau à remplir, la colonne où il y a le plus de croix est l’ensemble le plus inclusif. On termine l’exercice en épuisant toutes les colonnes du tableau.
En primaire, sur le plan matériel, les mêmes feuilles sont à découper, et les enfants par petits groupes devront coller les animaux dans des ensembles argumentés. Dans chaque groupe, les enfants discutent entre eux sur ce que les animaux ont. Cette discussion les pousse à observer les images et à confronter leurs savoirs sur ces espèces. Chaque groupe d’enfants produit donc des ensembles avec des arguments. Par exemple, l’ensemble (lapin + chat + homme) va associer les arguments des poils et des mamelles. On demande à chaque groupe d’enfants de dessiner les ensembles (des bulles, au besoin de couleurs différentes) et d’écrire les arguments à côté de l’ensemble. On encourage l’argumentation d’un ensemble sur la base de multiples arguments.
Au besoin, cet exercice va amener les enfants à reformuler leurs arguments lorsqu’ils se trompent de critère, pour les ramener à l’anatomie (c’est-à-dire ce que les animaux ont). Par exemple :
On ne dit pas : Je les mets ensemble parce qu’ils nagent (je classe sur ce qu’ils font)
On dit : Je les mets ensemble parce qu’ils ont des nageoires rayonnées (sur l’anatomie)
On ne dit pas : Je les mets ensemble parce que ce sont des mammifères (je classe sur ce qu’ils sont a priori)
On dit : Je les mets ensemble parce qu’ils ont des mamelles (sur l’anatomie : sur ce qu’ils ont en partage, et qui est observable)
Dans un second temps, tous les arguments de tous les groupes de la classe vont être lus, commentés collectivement et affichés au tableau. Pour l’enseignant, des agrandissements des figures d’animaux peuvent avoir été préalablement découpés sur du papier cartonné et affichés au tableau à l’aide de gomme à fixer. Les ensembles sont reconstitués à l’aide de tous les arguments produits et validés par la classe (Figure 9). On atteint la fin de l’exercice en épuisant la liste des attributs-arguments qui avait été dressée.
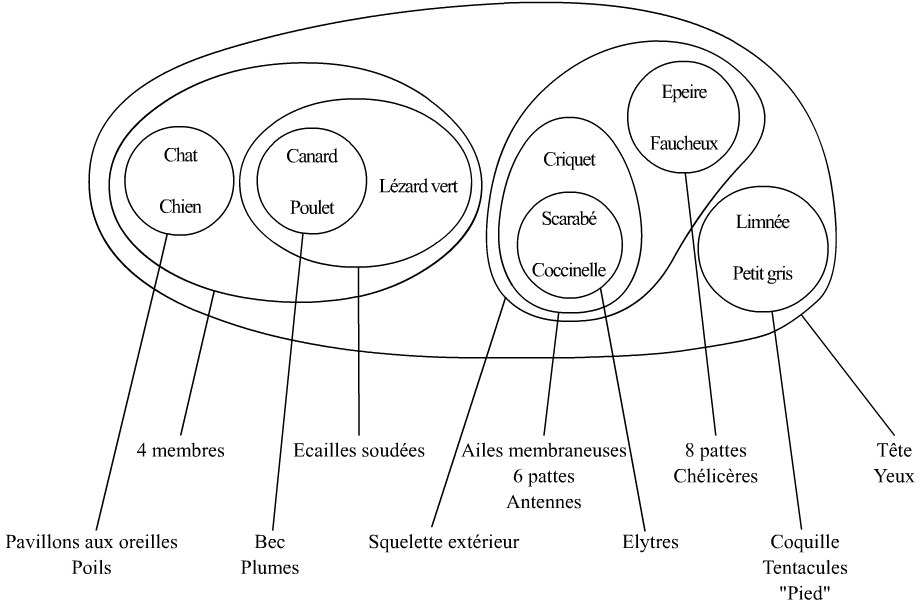
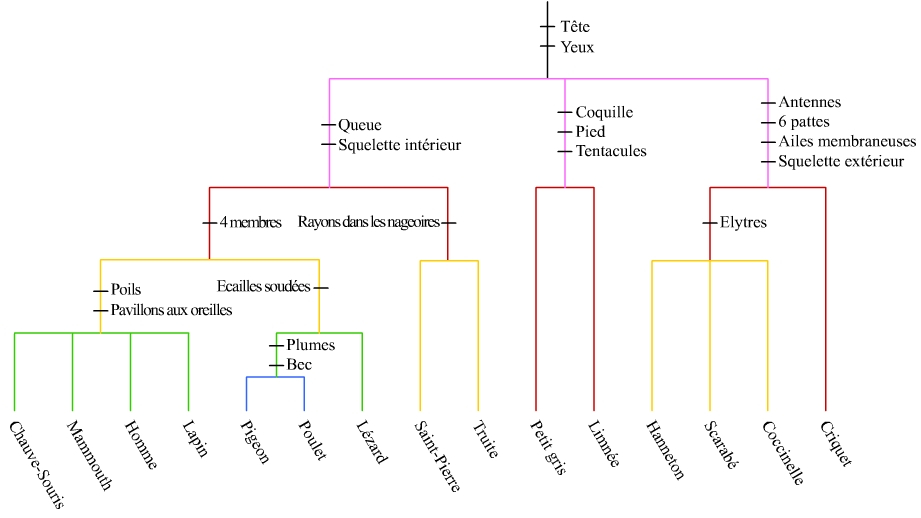
Ces arguments sont :
- (Hanneton, scarabée, coccinelle) : élytres
- Les précédents avec le criquet : six pattes, antennes, ailes membraneuses (éventuellement squelette à l’extérieur, argument surtout intéressant si on ajoute deux araignées)
- Limnée, escargot petit gris : tentacules, coquille, pied
- Pigeon, poulet : plumes, bec
- Pigeon, poulet, lézard vert : écailles soudées
- Chat, homme, lapin : pavillon aux oreilles, poils
- Chat, homme, lapin, lézard vert, pigeon, poulet : quatre membres
- Saint-pierre, truite : rayons dans les nageoires
- Saint pierre, truite, chat, homme, lapin, lézard vert, pigeon, poulet : Squelette à l’intérieur, queue
- Tous : tête, yeux
On constate les emboîtements.
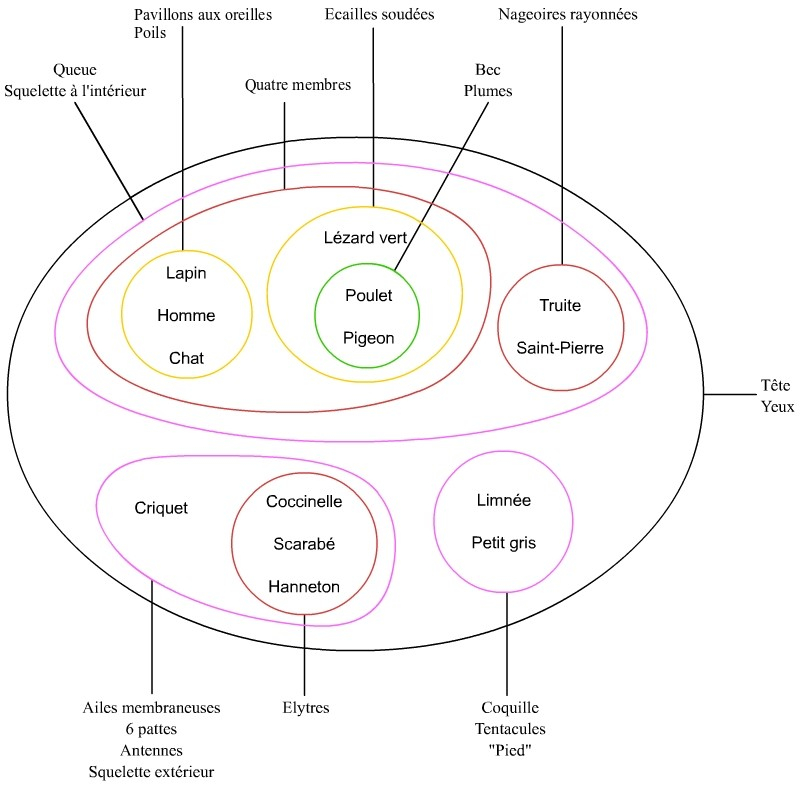
- Remarque n°1 :
Dans une première version de cet échantillon ne figurait qu’un seul oiseau, le pigeon. Le fait que l’oiseau soit le seul à représenter des plumes ne pose pas de problème particulier, on considèrera qu’il forme un ensemble à lui tout seul. Si cela pose problème, il suffit de rajouter un poulet dans l’échantillon, comme suggéré ici. - Remarque n°2 :
Le point fondamental de la séance est d’éviter de classer les animaux sur la base de ce qu’ils n’ont pas. Si l’on fait des ensembles sur des attributs-arguments constatés, il n’y a pas lieu de réunir limnée, petit gris, criquet, hanneton, coccinelle, scarabée dans un ensemble « invertébrés ». L’expérience montre d’ailleurs que, spontanément, et même sans consigne particulière, les enfants ne classent pas sur la base de ce que les animaux n’ont pas. Les groupes privatifs sans valeur scientifique comme « invertébrés » ou « agnates » sont des « pollutions culturelles » qui posent un problème plus pour les adultes que pour les enfants. - Remarque n°3 :
On n’est pas obligé de faire tous les ensembles possibles de l’échantillon. Pour un nombre donné d’objets à classer, on atteint la classification la plus précise lorsque chaque ensemble est constitué de deux objets ou deux ensembles. Dans ce cas la classification comportera le maximum d’information. Mais on peut très bien laisser un ensemble constitué de trois ensembles ou davantage. - Remarque n°4 :
Une fois les ensembles constitués, on peut amener une espèce « problématique », par exemple une chauve-souris. Cette espèce est problématique parce qu’elle amène de l’homoplasie, c’est-à-dire de la ressemblance trompeuse. Elle est traitée en assignation. On constate que la classe aura relevé deux arguments pour placer la chauve-souris avec le chat, l’homme et le lapin, tandis qu’un seul argument la place avec le pigeon et le poulet. On placera donc la chauve-souris avec ceux pour lesquels on peut énoncer le plus d’arguments.
En primaire seulement : aborder la causalité sous-jacente (la classification dit quelque chose sur le monde)
On peut demander à la classe pourquoi les espèces ont des choses en commun. Les réponses sont du type :
- Parce que Dieu les a faits comme cela (à l’occasion, on pourra définir ce qui est propre aux affirmations scientifiques)
- Parce qu’ils sont dans un même environnement
- Parce qu’ils font des bébés
- Parce qu’ils sont de la même famille
- Parce qu’ils sont cousins
- Parce qu’ils viennent du ventre de la même mère, mais cette mère vivait au temps de la préhistoire…
On peut amener les enfants à se souvenir que le déroulement de l’histoire d’une famille sur de très grandes périodes de temps, ça s’appelle la généalogie. Le mot peut d’ailleurs venir de la classe. Qu’ y a-t-il dans une généalogie ? : des ancêtres.
Pourquoi le chat et le lapin ont en commun des poils et des pavillons aux oreilles, alors qu’ils ne font plus de bébés ensemble ? Parce qu’ils l’ont hérité de leurs ancêtres, et plus précisément d’ancêtres communs.
Dès que le mot « généalogie » est lâché, les enfants parlent spontanément d’arbre. Certains enfants viennent même à dire que les espèces se transforment. Derrière les ensembles emboîtés, il y a la transformation passée des animaux au cours de leur généalogie.
Une fois que les mots-clés « ancêtres », « cousins », « généalogie », « transformation » ou « évolution », voire « arbre » ont émergé, tout est prêt pour expliquer que ce qu’ils ont en commun (et que d’autres n’ont pas), ils l’ont parce qu’ils l’ont hérité d’ancêtres communs à eux seuls (c’est-à-dire que ce ne sont pas les ancêtres des autres). Par exemple, les six pattes ont été hérités d’un animal qui a été l’ancêtre du hanneton, de la coccinelle, du scarabée, des deux papillons, mais qui n’est pas l’ancêtre des autres (chat, lapin, etc.), sinon les autres auraient six pattes également.
En primaire seulement : de la classification à l’arbre
Des ensembles emboîtés représentent un arbre vu du dessus. Chaque ensemble est une branche. Plus l’ensemble est inclusif et plus la branche correspondante est profonde. Deux ensembles de même niveau hiérarchique sont groupes-frères. Une série d’ensembles, par projection dans la troisième dimension, devient un arbre.
Du point de vue pratique, on peut :
- Soit laisser l’enseignant dessiner l’arbre derrière les ensembles
- Soit utiliser un mobile
- Soit créer une séance spéciale pour laisser les enfants dessiner l’arbre eux-mêmes. Dans ce cas, une couleur différente par ensemble et sa branche correspondante aidera à resituer visuellement les branches. On pourra proposer à l’élève qui a dessiné le bon arbre de venir l’expliquer ; puis vérifier qu’aucune information n’a été perdue des ensembles vers l’arbre.
- Soit donner d’emblée l’arbre correspondant aux ensembles dessinés au tableau et laisser les enfants placer eux-mêmes les animaux au bout des branches conformément à ces ensembles, exercice qui passe nécessairement par la compréhension du rapport qu’il y a entre les branches et les ensembles.
On peut replacer ensuite les arguments sur les branches de l’arbre (Figure 10). Là, on pourra constater que les oiseaux et la chauve-souris qui avait été donnée en assignation ont appris à voler deux fois indépendamment.
Assignation : placer une espèce nouvelle dans la classification
On peut proposer de placer un mammouth dans l’arbre final. Il suffit que les enfants vérifient les attributs (arguments) que présente le mammouth et le placent en fonction : il a une tête, des yeux, un squelette à l’intérieur, une queue, quatre membres, des pavillons aux oreilles et des poils. Ce qu’on montre là, c’est que le fossile ne sera pas rangé à un nœud de l’arbre mais au bout d’une branche, comme les animaux actuels.
Apprendre des mots nouveaux
Éventuellement, on peut apprendre les noms des ensembles constitués en classe :
- Tête, yeux : animaux (nom scientifique : métazoaires)
- Six pattes, antennes, ailes membraneuses : insectes (se confond avec arthropodes en absence de crustacés ou d’araignée)
- Élytres : coléoptères
- Coquille, tentacules, pied : mollusques
- Squelette à l’intérieur, queue : vertébrés
- Rayons dans les nageoires : actinoptérygiens
- Quatre membres : tétrapodes
- Plumes, bec : oiseaux
- Écailles soudées : sauropsides
- Pavillons aux oreilles, poils : mammifères
Précautions
Par cette séquence, on a atteint un certain nombre d’objectifs :
- On a parlé de classification phylogénétique sans le dire ;
- On a évacué les groupes privatifs anthropocentriques (invertébrés, agnathes, etc.) ;
- On a développé une démarche classificatoire qui va de l’observation vers la classification, et non la démarche inverse utilisant les a priori, c’est-à-dire lorsqu’on projette ce qu’on croit savoir sur notre perception du vivant pour conforter une classification mentale souvent rassurante mais fausse du point de vue phylogénétique ;
- On a posé l’idée qu’une classification dit quelque chose sur le monde (transformation au cours de la généalogie), et la métaphore de l’arbre y contribue ;
- On a fourni des résultats compatibles avec ce que la science produit aujourd’hui.
Des dizaines de séances différentes peuvent être inventées sur ce modèle, en fonction de la collection d’espèces à classer. On peut collecter des espèces au bord de la mer, en forêt, au bord des étangs, etc. et construire une telle séance avec un nombre limité d’échantillons. Cependant, les difficultés principales, pour l’enseignant, viendront du rapport qu’il y a entre les ressemblances dues à des convergences évolutives et celles qui attestent une réelle parenté. En d’autres termes, le risque est que la démarche puisse être réussie et comprise en classe, tout en aboutissant à une classification fausse du point de vue phylogénétique. La préparation d’une telle séance demande donc beaucoup de travail documentaire en amont, pour doser la difficulté provenant des arguments potentiellement contradictoires entre eux, et vérifier que ceux qui ont le plus de chances d’émerger de la classe soient réellement des arguments conduisant à une classification phylogénétique.
En science, c’est le cadre théorique de l’évolution biologique qui fixe son « cahier des charges » à la classification. Dans le cadre scolaire élémentaire français, on apprend la classification alors que l'évolution biologique n'est pas encore assimilée : la classification est prescrite pour aller vers l'évolution ; tandis qu'en science moderne, l'évolution est un préalable à la classification. C’est la raison pour laquelle l’enseignant ne classera jamais qu’une collection contrôlée d’organismes tant qu’il ne peut pas construire une phylogénie comme cela a été fait en section 4.2. Cela dit, cela n’est pas complètement absurde car permet d’étager nos objectifs pédagogiques. Les premiers objectifs de la classification sont cognitifs et philosophiques avant que d’être biologiques :
- Cognitifs : distinguer les opérations de tri (séparation), d’assignation (rangement) et de classification (regroupement, agglomération : faire des ensembles).
- Philosophiques : c’est nous qui classons, selon certains besoins (décrire et comprendre le monde qui nous entoure parce qu’on est en science) et certaines règles du jeu (on va classer sur ce que les espèces partagent en commun) : posture nominaliste.
- Biologiques : si on classe sur la base de ce que les organismes vivants ont en commun (et non sur ce qu’ils font, ou là où ils habitent…), c’est qu’on cherche à retracer les ascendances communes (selon Darwin, 1859).
Bibliographie sélective en langue française
- Guillaume Lecointre. 2011. Filiation. Pp. 271-334 In HEAMS, T., HUNEMAN, P., LECOINTRE, G., SILBERSTEIN M. (Dir.). Les Mondes Darwiniens. L’évolution de l’évolution (Tome I). Syllepse. Paris. Seconde édition, 1576 pp.
- Lecointre Guillaume (dir.). (2004). Comprendre et enseigner la classification du vivant. Belin.
- Lecointre Guillaume (dir.). (2020). Guide critique de l’évolution. Belin-Éducation.
- Tassy Pascal. (1991). L’arbre à remonter le temps. Christian Bourgois.
- Lecointre Guillaume. (2002). La classification du vivant. In « Graines de Sciences 4 » (sous la direction de D. Wilgenbus, J.-M. Bouchard et P. Léna). Le Pommier, Paris, pp. 173-218.
- Linné : classer la Nature. Les génies de la science n°26. Pour La Science. (2006).
Pour aller plus loin
- Darlu Pierre et Tassy Pascal. (1993). Reconstruction phylogénétique. Seconde édition. Éditions Matériologiques (2019).
- Lecointre Guillaume et Le Guyader Hervé. (2016). Classification phylogénétique du Vivant. Tome I. Quatrième édition. Belin.
- Lecointre Guillaume et Le Guyader Hervé. (2017). Classification phylogénétique du Vivant. Tome II. Quatrième édition. Belin.
Glossaire
Remarque : les noms de taxons ne sont pas reportés dans le glossaire (agnathes, archosaures, lépidosaures, amniotes…). Pour leur définition, on se reportera à Lecointre et Le Guyader (2001). Les noms de structures anatomiques sont accessibles dans tout livre d’anatomie ou dictionnaire médical (os dentaire, articulaire, angulaire, carré, tympanique, métatarsien, marteau, enclume, radius, amnios).
| Mot | Définition |
|---|---|
| Anthropocentrisme | Attitude consistant à mettre l’Homme au centre de toute chose, auquel toute réalité du monde doit se rapporter. |
| Attribut | Structure portée par un organisme. |
| Caractère | Attribut d’un organisme ou d’un taxon par lequel il diffère d’autres taxons, et sur lequel on peut formuler une hypothèse d’homologie. |
| Clade | Groupe monophylétique |
| Cladistique | Synonyme de « Systématique phylogénétique ». |
| Classer | Créer des taxons dont l’emboîtement traduit la hiérarchie de la distribution des caractères dans la Nature. |
| Classification | Système de taxons emboîtés. |
| Espèce | Ensemble monophylétique d’individus se reconnaissant comme partenaires sexuels, et capables de donner une descendance féconde. |
| Extra-groupe | Espèce ou Taxon que l’on suppose a priori extérieur à l’ensemble des espèces que l’on se propose de classer. Synonyme de « groupe extérieur ». En systématique phylogénétique, les extra-groupes font partie des postulats de départ de toute activité classificatoire. |
| Fixisme | Doctrine stipulant que les espèces sont immuables. |
| Généalogie | Figure arborescente dans laquelle les lignes traduisent des liens génétiques d’ancêtres à descendants, et dans laquelle les ancêtres sont identifiés individuellement. |
| Groupes-frères | Au sein d’un échantillon donné, deux groupes (ou taxons) sont qualifiés de frères lorsqu’ils partagent un ancêtre commun exclusif, c’est-à-dire non partagé par le reste de l’échantillon. |
| Homologie | Similarité chez plusieurs espèces d’organes ou de parties d’organes dont on peut faire le pari qu’elle est héritée par ascendance commune. |
| Homologie primaire | Sont homologues deux attributs qui, pris chez des espèces différentes, entretiennent avec les structures voisines les mêmes connexions topologiques, et ceci quelles que soient leurs formes et leurs fonctions. |
| Homologie secondaire | Deux attributs sont homologues s’ils sont hérités par voie d’ascendance commune. C’est l’arbre phylogénétique qui, au final, établit l’homologie secondaire. |
| Homoplasie | Similarité chez plusieurs espèces d’attributs qui n’est pas héritée par ascendance commune. C’est l’arbre phylogénétique qui, au final, établit l’homoplasie. |
| Monophylétique | Se dit d’un groupe comprenant un ancêtre commun hypothétique et la totalité de ses descendants connus. Exemple : les oiseaux. |
| Ontogénèse | Processus du développement de l’organisme, de l’œuf fécondé jusqu’à l’adulte. |
| Paraphylétique | Se dit d’un groupe comprenant un ancêtre commun hypothétique et une partie seulement de ses descendants connus. Exemple : les poissons. |
| Polyphylétique | Se dit d’un groupe ne comprenant pas d’ancêtre commun hypothétique. Exemple : les pachydermes. |
| Phylogénétique | Se dit d’un arbre traduisant des relations de parenté, c’est-à-dire des relations de groupes-frères (qui est plus proche de qui). En d’autres termes, l’arbre phylogénétique dit quels groupes ont des ancêtres hypothétiques communs et exclusifs. |
| Phylogénie | Au sens large, c’est-à-dire au sens de Haeckel (1866), le cours historique de la descendance des êtres organisés. C’est l’abre qui montre la genèse (Genesis) des lignées (Phylum). Au sens strict, relations de parenté : la phylogénie dit quels taxons sont plus étroitement apparentés entre eux qu’à d’autres. |
| Phylogénèse | Processus d’individualisation des lignées évolutives. |
| Polarisation (des caractères) | Etablir l’état primitif et l’état (ou les états) dérivé(s) d’un caractère, en d’autres termes établir le sens de la transformation d’un caractère. |
| Saut adaptatif | Discontinuité dans l’organisation générale des êtres vivants, interprétée comme étant due à une adaptation brusque à de nouvelles conditions ou niches écologiques. Ces discontinuités se sont avérées être des artéfacts d’échantillonnage. Par exemple, la classe des « reptiles » n’existe que parce qu’on a cru détecter une discontinuité d’organisation entre des amniotes poïkilothermes (« à sang froid » : tortues, lézards, crocodiles…) et des amniotes à sang chaud (mammifères et oiseaux). En fait, les « reptiles mammaliens » et les découvertes récentes de dinosaures à plumes montrent que cette discontinuité n’existe pas. |
| Systématique | Science des classifications biologiques. |
| Systématique éclectique | Ecole traditionnelle de la systématique évolutionniste classant êtres vivants sur la base des « affinités évolutives » (la phylogénie) autant que sur la base des « sauts adaptatifs », lesquels prenaient en compte à la fois le « degré de complexité » et des critères écologiques. Cette école admettait comme valides les groupes paraphylétiques et les groupes monophylétiques. |
| Systématique phylogénétique | Méthode de classification évolutionniste, initiée par Willi Hennig, se fondant sur l’apparentement phylogénétique et lui seul. La cladistique n’admet donc comme valides que les taxons monophylétiques. |
| Taxon | Ensemble d’organismes regroupés en tant qu’unité formelle. Par exemple, les canidés (chien, loup, renard, fennec, etc.) constituent un taxon de rang familial. |
| Transformisme | Théorie de la transformation des espèces au cours du temps. |
Quelques idées fausses et les arguments pour les démonter
Idée fausse n°1
Les étiquettes de « groupes » dans les clés de détermination justifient les « invertébrés », les « poissons » et les « reptiles » qui sont des points de repère pratiques.
Idée fausse n°2
Une recherche sur le mot-clé « actinoptérygiens » via Google donne 48 100 pages ; la même recherche sur le mot « poissons » donne plus de seize millions de pages. Ce n’est pas la peine de changer notre enseignement.
Réponse couplée :
On peut objecter à plusieurs niveaux à ces remarques, du plus particulier au plus général :
- Google enregistre le mot « poissons » pour des sites culinaires et de nombreux sites d’astrologie.
- De tels « tests » n’ont que peu de valeur diagnostique, car ils mélangent des sites aux qualités extrêmement hétérogènes.
- Ne s’agirait-il que de zoologie, la plupart des créateurs de sites n’ont eu à l’école primaire et secondaire que des cours traditionnels, puisque les programmes viennent de changer.
- Si l’on se renseigne vraiment sur des sites spécialisés (et là il faudrait souvent passer du français à l’anglais), on constate que le grand projet de classer le vivant sur la base de sa phylogénie est international.
Idée fausse n°3
Il faut bannir le mot « poissons ».
Réponse :
Il n’en a jamais été question. Le mot « poissons » restera dans la langue. Il renvoie à un registre culinaire, et à un registre mythologique. L’objectif des classifications pertinentes est d’utiliser les bons mots dans les registres adéquats. L’idée générale est qu’on n’utilise pas le mot « poissons » dans un registre (dit aussi « cahier des charges ») phylogénétique, pas plus qu’on ne dira « actinoptérygiens » chez le poissonnier.
En sciences, la justification du mot « poissons » dans un cahier des charges écologique est loin d’être évidente. Il existe une diversification écologique considérable des poissons, qui les rend extrêmement hétérogènes sur ce plan. Si l’on disait simplement que les poissons sont les vertébrés aquatiques, ce serait insuffisant. Il faudrait amener toute suite une longue liste d’exceptions, c’est-à-dire de vertébrés aquatiques qu’on ne met pas dans les poissons : cétacés, phoques, morses, otaries, loutres, siréniens, manchots, pingouins, cormorans, crocodile marin, serpent marin (Laticauda), tortues marines, ichtyosaures, sauroptérygiens, mosasaures, etc.
Idée fausse n°4
Il ne faut pas aller contre les représentations des élèves.
Réponse :
Cette vision est liée au refus ou à l’échec pédagogique que constituerait une superposition brutale d’une vision du monde (scientifique) sur une autre (non scientifique).
En sciences, il faut d’abord distinguer les discours de valeurs et les discours sur les faits. Les discours sur les valeurs ne sont pas gérés par la science. En cours de sciences, on enseigne le résultat d’une investigation rationnelle menée sur le monde réel à l’aide d’expériences reproductibles. Si la science dit que l’homme est un animal, ce n’est pas pour le dégrader (perception par les valeurs), c’est une décision nomenclaturale fondée sur des observations : pourvu de muscles et de nerfs, l’homme possède les attributs de tous les métazoaires (= animaux). Si la science dit que l’homme est un singe, c’est une décision nomenclaturale fondée sur des observations : pourvu d’os frontaux fusionnés, l’homme possède les attributs de tous les simiiformes (= singes vrais). Au mieux, il faut prendre les représentations des élèves sur les faits pour les transformer.
Tout scientifique et tout enseignant de sciences se doit d’initier les élèves à des réalités du monde qui échappe à nos sens. C’est ce que font les sciences depuis leurs origines. On n’enseigne plus que le soleil tourne autour de la terre, et pourtant chaque élève voit chaque jour le soleil se lever à l’Est et le soir se coucher à l’Ouest.
Idée fausse n°5
Une phylogénie est un arbre fondé sur les gènes.
Réponse :
Faux : Le mot est inventé en 1866 par Ernst Haeckel. Il signifie l’arbre qui raconte la genèse (genesis) des lignées (phylum). Cela étant dit, certaines phylogénies modernes peuvent être fondées sur la comparaison entre espèces de séquences d’ADN. Mais la phylogénie peut autant être fondée sur l’anatomie comparée.
Idée fausse n°6
Le bouleversement de la classification est dû aux phylogénies moléculaires.
Réponse :
En partie faux. Les classifications ont toujours changé. Le bouleversement massif et récent des classifications a débuté avec les approches purement phylogénétiques de la classification, dès 1950 ; et plus massivement à partir de 1966. Les premiers bouleversements ont été fondés sur de l’anatomie comparée. L’accès à la comparaison de molécules pour faire des phylogénies a débuté timidement dans les années 1965-1970 ; et beaucoup plus massivement à partir de 1990, pour des raisons techniques.



