
Les molécules d’ARN messager (ARNm) et les protéines qu’elles codent sont généralement représentées sur les schémas sans tenir compte de la taille relative de ces deux types de macromolécules. Combien mesurent et pèsent les ARN messagers, ainsi que les protéines qu’ils codent ? Et, entre les deux, qui est le plus gros : l’ARNm ou la protéine codée ?
L’expression des gènes codant des protéines nécessite un intermédiaire, l’ARN messager (ARNm). Dans les ouvrages traitant de biologie, qu’il s’agisse de manuels scolaires ou universitaires, ainsi que sur internet, les molécules d’ARNm sont souvent représentées par un simple trait ou par les nucléotides qui les composent, tandis que les protéines sont généralement dessinées sous la forme d’un collier d’acides aminés ou sous une forme globulaire (Figure 1). Quoi qu’il en soit, sur de tels schémas, les protéines sont généralement aussi grosses ou plus grosses que les ARNm qui ont servi à leur production. Or, en réalité, les ARN messagers sont plus grands et plus lourds que les protéines qu’ils codent. Pour le montrer, nous allons voir quels sont les ordres de grandeur de masse et de taille des protéines, puis des ARN messagers, pour terminer en comparant ces deux types de macromolécules.

Le gène, sous forme d’ADN, est d’abord transcrit en ARN prémessager qui, une fois maturé en ARN messager, est traduit en protéine.
La masse des protéines
Masses molaires des acides aminés
Tous les acides aminés possèdent un squelette commun, formé d’un carbone α, d’un groupement carboxyle COOH, d’un groupement amine NH2 et d’un atome d’hydrogène (Figure 2). De formule brute C2H4NO2, ce squelette pèse 74 g/mol1. Les masses des acides aminés varient alors en fonction de leurs chaînes latérales : la glycine est l’acide aminé le plus léger, tandis que le tryptophane est le plus lourd (Tableau 1).

Quatre acides aminés sont présentés à titre d’exemple. Les atomes sont représentés sous forme de boules (noir : carbone, bleu : azote, rouge : oxygène, blanc : hydrogène) reliés entre eux par des bâtonnets, symbolisant les liaisons chimiques. Sur toutes ces représentations, le groupement carboxyle COOH (ici sous forme carboxylate COO−, comme c’est le cas au pH cellulaire) est placé à droite. Il est relié au carbone α, lui-même lié au groupement amine (ici sous forme ammonium NH3+, comme c’est le cas au pH cellulaire) visible en bas à gauche de celui-ci. La chaîne latérale, variable selon les acides aminés, occupe donc la partie gauche de chaque molécule. Chez la glycine, cette chaîne latérale se limite à un atome d’hydrogène.
Source des images : aspartate, glutamine et tryptophane : Jynto, CC0, Wikimedia. Glycine : Ben Mills, domaine public, Wikimedia.
| Nom | Masse molaire 1 (g/mol) | Fréquence dans les séquences de la banque de données UniProtKB/Swiss-Prot 2 |
|---|---|---|
| Alanine | 89,09 | 8,26 |
| Arginine | 174,2 | 5,53 |
| Asparagine | 132,12 | 4,06 |
| Aspartate | 133,1 | 5,46 |
| Cystéine | 121,15 | 1,39 |
| Glutamate | 147,13 | 6,72 |
| Glutamine | 146,15 | 3,93 |
| Glycine | 75,07 | 7,07 |
| Histidine | 155,16 | 2,28 |
| Isoleucine | 131,17 | 5,91 |
| Leucine | 131,17 | 9,65 |
| Lysine | 146,19 | 5,8 |
| Méthionine | 149,21 | 2,41 |
| Phénylalanine | 165,19 | 3,87 |
| Proline | 115,13 | 4,75 |
| Sérine | 105,09 | 6,66 |
| Thréonine | 119,12 | 5,37 |
| Tryptophane | 204,23 | 1,11 |
| Tyrosine | 181,19 | 2,92 |
| Valine | 117,15 | 6,85 |
À partir de ce tableau, il est possible de déterminer la masse molaire moyenne d’un acide aminé. Le calcul de la moyenne brute donne un résultat de 136,90 g/mol. Toutefois, la pondération de la moyenne par la fréquence de chaque acide aminé au sein des protéines permet d’aboutir à une valeur de 129,11 g/mol.
Masses molaires des protéines
Pour calculer la masse molaire d’une protéine, il faut tenir compte des molécules d’eau (MH2O = 18,01 g/mol) éliminées lors de la formation des liaisons peptidiques entre les acides aminés.
La masse molaire moyenne d’une protéine formée par n acides aminées, et contenant donc (n − 1) liaisons peptidiques est :
Mprotéine = n × 129,11 − (n − 1) × 18,01
Mprotéine = n × 111,10 + 18,01
Mprotéine ~ n × 111 (Équation 1)
La masse moyenne d’un acide aminé présent dans une protéine est d’environ 111 g/mol.
Quelques exemples de masses molaires de protéines
Le site UniProt fournit de nombreuses informations sur les protéines dont, en particulier, leurs nombres d’acides aminés et leurs masses molaires 3. Les masses molaires estimées d’après l’Équation 1 ainsi que les masses molaires réelles de quatre protéines sont données dans le Tableau 2. La lecture de ce tableau montre que l’approximation Mprotéine ~ n × 111 donne des estimations très proches des valeurs réelles.
| Protéine | Référence UniProt | Organisme | Nombre d’acides aminés | Masse molaire estimée d’après l’Équation 1 (g/mol) | Masse molaire réelle (g/mol) |
|---|---|---|---|---|---|
| Prépro-insuline | P01308 | Homo sapiens | 110 | 12 210 | 11 981 |
| Sous-unité α 1 de l’hémoglobine | P69905 | Homo sapiens | 142 | 15 762 | 15 258 |
| Grande sous-unité de la rubisco | O03042 | Arabidopsis thaliana | 479 | 53 169 | 52 955 |
| Titine | Q8WZ42 | Homo sapiens | 34 350 | 3 812 850 | 3 816 030 |
La taille des protéines
La taille des protéines est intrinsèquement liée à leur forme. Les protéines peuvent très schématiquement être regroupées en deux catégories de formes : globulaires ou fibrillaires. Il existe également des protéines désordonnées, c’est-à-dire sans forme stable. Par ailleurs, les protéines globulaires et fibrillaires peuvent présenter des portions désordonnées. Les protéines globulaires à structure tertiaire mesurent généralement quelques nanomètres de diamètre. Par exemple l’insuline humaine (2HIU4) mesure environ 3 nm, tandis que la myoglobine de grand cachalot (1MBD) fait entre 4 et 5 nm de diamètre (Figure 3). Les protéines à structure quaternaire, composées de plusieurs sous-unités, sont logiquement plus grandes. Par exemple la rubisco d’épinard (1RCX) présente un diamètre de 12 nm environ, alors que le ribosome de levure de boulanger (6SNT) a une taille de 25 nm environ5.

Cette image a été produite à l’aide du logiciel Mol* grâce à des structures protéiques disponibles sur le site de la banque de données sur les protéines (PDB) : insuline humaine (2HIU), myoglobine de grand cachalot (1MBD), rubisco d’épinard (1RCX) et ribosome de levure de boulanger (6SNT). Sur ce dernier, les protéines sont colorées en jaune, les ARNr en orange et un morceau de l’ARN messager est visible en magenta.
Les protéines fibrillaires sont formées de sous-unités qui sont soit elles-mêmes fibrillaires, soit globulaires. Le premier cas de figure est représenté par le collagène, formé de trois hélices enroulées. Le tout mesure environ 300 nm de long pour un diamètre de 1,5 nm. Dans la seconde catégorie se trouvent par exemple les microfilaments d’actine et les microtubules. Les microfilaments d’actine se composent d’actine G, globulaire. Assemblées, ces sous-unités produisent des microfilaments d’actine pouvant atteindre plusieurs micromètres de long. Les microtubules sont quant à eux formés de 13 protofilaments eux-mêmes composés de protéines globulaires, les tubulines α et β (le dimère αβ est visible à l’entrée 1TUB de la banque de données sur les protéines). Les microtubules forment des tubes creux d’un diamètre de 25 nm et d’une longueur pouvant atteindre plusieurs micromètres.
Un aperçu de la diversité des protéines du vivant
Deux ressources en ligne permettent d’appréhender la diversité de tailles et de formes des protéines du vivant. La première, en deux dimensions, est hébergée par le site de la banque de données sur les protéines (PDB). La seconde, en trois dimensions, utilise le logiciel Mol* (à prononcer Molstar).
La masse des ARN messagers
Masses molaires des nucléotides
Les ARN messagers sont formés de quatre ribonucléotides monophosphates différents (Figure 4 et Tableau 3).
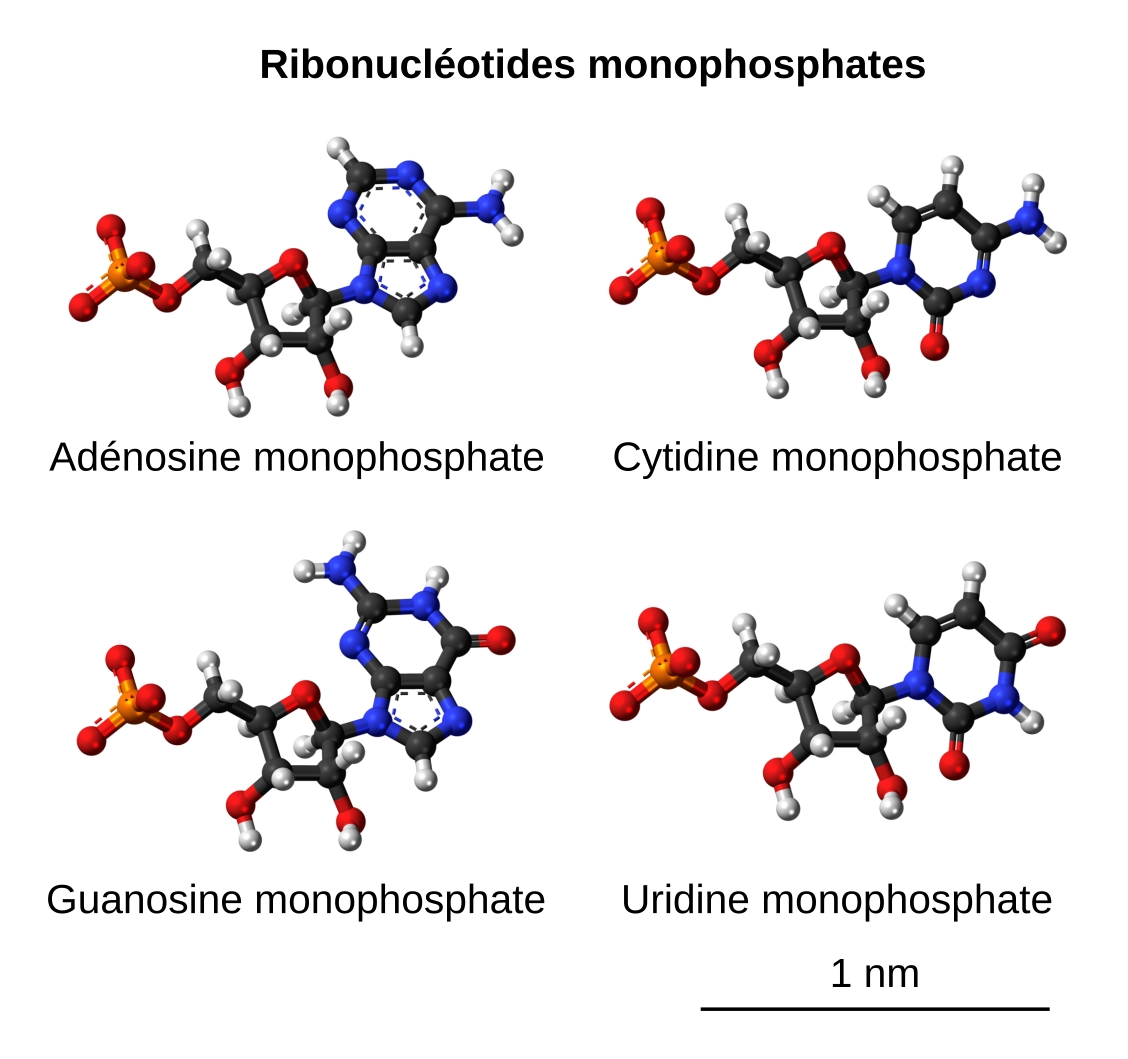
Les atomes sont représentés sous forme de boules (noir : carbone, bleu : azote, rouge : oxygène, blanc : hydrogène, orange : phosphore) reliés entre eux par des bâtonnets, symbolisant les liaisons chimiques. Sur toutes ces représentations, le phosphate est à gauche, le ribose au centre et la base azotée à droite.
Source des images : Jynto, CC0, Wikimedia : adénosine monophosphate, cytidine monophosphate, guanosine monophosphate, uridine monophosphate
| Ribonucléotides monophosphates | Formules brutes | Masses molaires (g/mol) |
|---|---|---|
| Adénosine monophosphate | C10H12N5O7P2− | 345,21 |
| Uridine monophosphate | C9H11N2O9P2− | 322,17 |
| Cytidine monophosphate | C9H12N3O8P2− | 321,18 |
| Guanosine monophosphate | C10H12N5O8P2− | 361,21 |
Les formules brutes sont données ici pour des nucléotides dont le groupement phosphate porte deux charges négatives, ce qui est le cas au pH physiologique.
La masse molaire moyenne d’un nucléotide monophosphate, calculée à partir des quatre valeurs du tableau, vaut 337,44 g/mol.
Masses molaires des ARNm
Pour calculer la masse molaire d’une molécule d’ARN, il faut tenir compte de l’élimination d’un groupement OH (MOH = 17,01 g/mol) lors de la formation des liaisons phosphodiesters.
La masse molaire d’une molécule d’ARN formée par n ribonucléotides, et contenant donc (n − 1) liaisons phosphodiesters est :
MARN = n × 337,44 − (n − 1) × 17,01
MARN = n × 320,43 + 17,01
MARN ~ n × 320 (Équation 2)
La masse moyenne d’un ribonucléotide présent dans un ARN est d’environ 320 g/mol.
Quelques exemples de masses molaires d’ARNm
L’outil Nucleotide du site du National Center for Biotechnology Information fournit de nombreuses informations sur les gènes et leurs transcrits dont, en particulier, la longueur (en nombre de nucléotides) des ARNm. Les masses molaires estimées d’après l’Équation 2 ainsi que les masses molaires réelles de quatre ARNm sont données dans le Tableau 4. L’approximation MARN ~ n × 320 donne des estimations très proches des valeurs réelles.
| ARNm | Référence | Organisme | Nombre de nucléotides | Masse molaire estimée d’après l’Équation 2 | Masse molaire réelle, d’après la séquence |
|---|---|---|---|---|---|
| Insuline (variant 1) | NM_000207.3 | Homo sapiens | 465 | 148 800 | 148 796 |
| Sous-unité α 1 de l’hémoglobine | NM_000558.5 | Homo sapiens | 577 | 184 640 | 183 553 |
| Grande sous-unité de la rubisco | L24073.1 | Oryza sativa | 1355 | 433 600 | 432 851 |
| Titine, isoforme N2-A | NM_133378 | Homo sapiens | 101 520 | 32 486 400 | 32 510 844 |
Notez que les banques de données fournissent le nombre total de nucléotides de l’ARNm (sans la queue polyA toutefois, puisque celle-ci n’est pas issue de la transcription du génome, mais ajoutée après la transcription) et non pas uniquement de la séquence codante. Autrement dit, la longueur des extrémités non traduites en 5’ (5’UTR) et en 3’ (3’UTR) sont également prises en compte (Tableau 5).
| ARNm | Référence | Nombre de nucléotides de l’extrémité 5’UTR | Nombre de nucléotides de la séquence codante (y compris codon stop) | Nombre de nucléotides de l’extrémité 3’UTR | Nombre de nucléotides de l’ARNm mature (sans la queue polyA) |
|---|---|---|---|---|---|
| Insuline (variant 1) | NM_000207.3 | 59 | 333 | 73 | 465 |
| Sous-unité α 1 de l’hémoglobine | NM_000558.5 | 37 | 429 | 111 | 577 |
| Grande sous-unité de la rubisco | L24073.1 | 34 | 1 206 | 115 | 1 355 |
| Titine, isoforme N2-A | NM_133378 | 225 | 100 272 | 1023 | 101 520 |
Chez les êtres vivants pluricellulaires, la longueur moyenne de l’extrémité 5’ non traduite est d’environ 300 nucléotides, tandis qu’elle est d’environ 700 nucléotides pour l’extrémité 3’ non traduite (calculs réalisés grâce au site de la banque de données sur les extrémités non traduites, UTRdb).
De plus, les ARNm peuvent porter des modifications post-transcriptionnelles, dont certaines sont réversibles, qui ne sont pas prises en compte ici. La plus étudiée d’entre elles est l’addition d’un groupement méthyle en position 6 sur les adénines, modification qui régule la traduction et le métabolisme des ARNm.
La taille des ARN messagers
Les ARN sont composés d’un enchaînement de ribonucléotides monophosphates. Chaque nucléotide occupe une « hauteur » de 0,5 nm environ. Cependant, il n’est pas possible de déterminer simplement la longueur totale d’un ARN messager en multipliant la place occupée par un nucléotide par le nombre de nucléotides. En effet, les ARN messagers, comme les autres ARN (ARNt, microARN…) présentent des structures secondaires par appariement de nucléotides (structure tige-boucle par exemple). Dans les ARNm, ces structures secondaires sont séparées par des régions non structurées et la flexibilité qui en résulte rend difficile la détermination expérimentale de la structure tridimensionnelle des ARNm1.
Pour se représenter convenablement les ARN messagers dans un contexte cellulaire il faut également considérer que ces molécules ne sont pas nues, mais associés à de nombreuses protéines. Ainsi, chez les Eucaryotes, durant la transcription, des protéines se lient au niveau des jonctions introns-exons et contrôlent l’épissage. Les ARN messagers matures sont également liés à de nombreuses protéines : protéines s’associant à la coiffe 5’ et à la queue polyA en 3’ et circularisant les ARN messagers, protéines du complexe de la jonction exonique et, bien entendu, ribosomes.
Bilan
Taille relative des nucléotides et des acides aminés
Les nucléotides, qui mesurent environ 1,1 nm, sont à peu près 1,5 fois plus grands que la taille moyenne d’un acide aminé, de 0,7 nm (Figure 5). Par ailleurs, leur masse molaire est environ trois fois plus élevée (320 g/mol en moyenne pour un ribonucléotide, contre 111 g/mol pour un acide aminé).
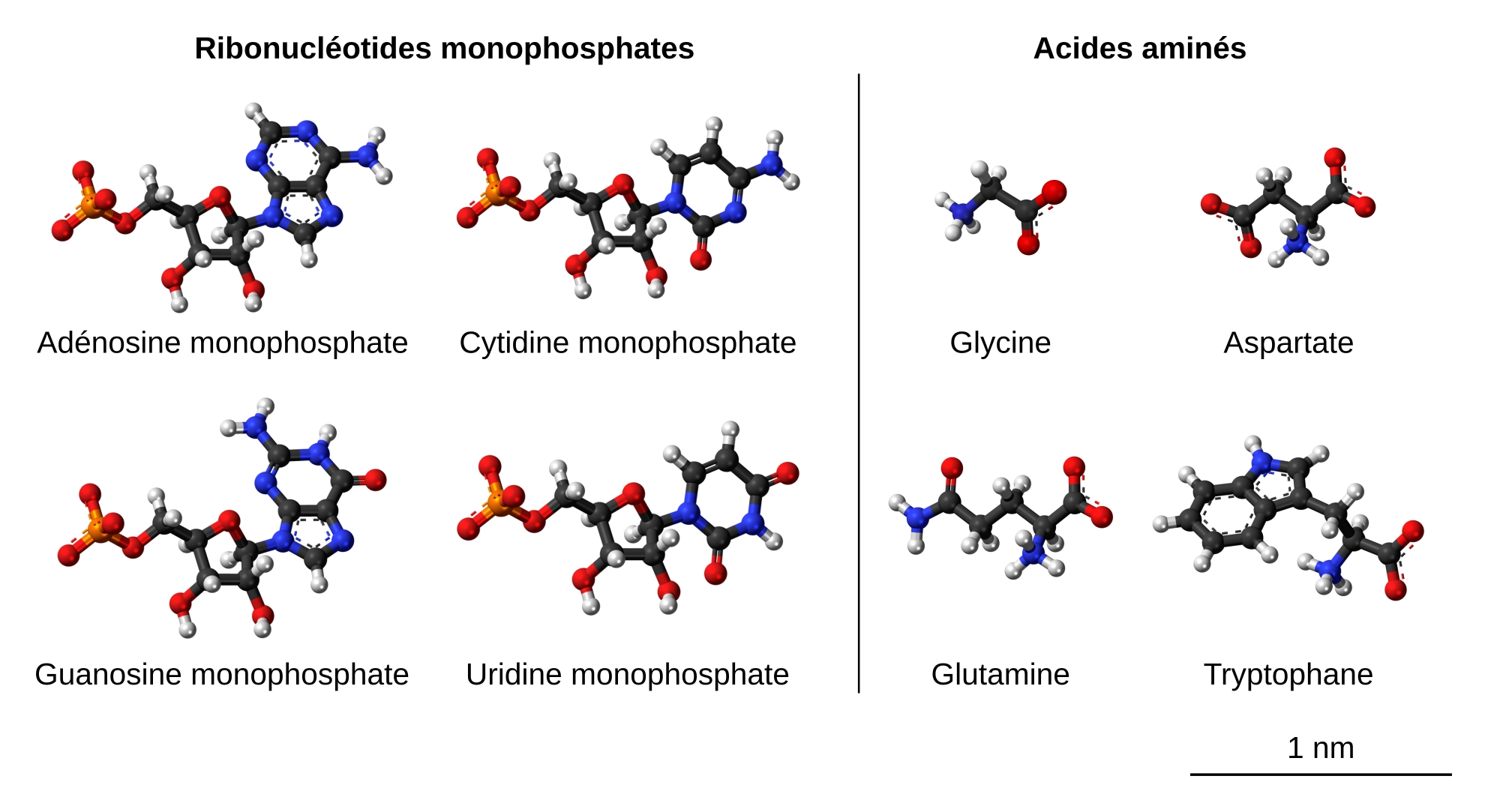
Les différents nucléotides sont représentés sous leur forme ionisée. Ils mesurent environ 1,1 nm. Le plus petit des acides aminés, la glycine, a une longueur d’à peu près 0,4 nm tandis que le plus grand, le tryptophane, mesure environ 1 nm.
Source des images : Jynto, CC0, Wikimedia : adénosine monophosphate, cytidine monophosphate, guanosine monophosphate, uridine monophosphate, acide aspartique, glutamine, tryptophane. Ben Mills, domaine public, Wikimedia : glycine
Taille relative des ARNm et des protéines qu’ils codent
Une fois polymérisés, la masse molaire d’un nucléotide ou d’un acide aminé est un peu plus faible que celle de sa version libre, à cause de l’élimination d’un groupement hydroxyle ou d’une molécule d’eau, respectivement. Cependant, la masse molaire d’un nucléotide reste toujours trois fois supérieure à celle d’un acide aminé (Mnucléotide = 320 g/mol ; Macide aminé = 111 g/mol).
Par ailleurs, comme il faut un triplet de nucléotides pour coder un acide aminé, cela signifie que la masse d’un ARNm est au moins neuf fois supérieure à celle de la protéine qu’il code. En réalité, ce rapport est souvent encore plus élevé, car il faut également tenir compte de la masse des séquences non traduites (5’ UTR et 3’UTR ; Tableau 6). Ainsi, dans les cellules, la notice de fabrication (l’ARNm) est bien plus lourde que le produit lui-même (la protéine) !
| Masse ARNm | Masse protéine | Rapport | |
|---|---|---|---|
| Insuline (variant 1) | 148 796 | 11 981 | 12 |
| Sous-unité α 1 de l’hémoglobine | 183 553 | 15 258 | 12 |
| Grande sous-unité de la rubisco | 432 851 | 52 955 | 8 |
| Titine, isoforme N2-A | 32 510 844 | 3 816 030 | 9 |
L’observation d’une unité de traduction en microscopie électronique à transmission permet de se rendre compte de la taille relative d’un ARN messager et de la protéine qu’il code (Figures 6 et 7). Il est également possible de transmettre cette idée sous forme de schéma (Figure 8).
Cette image, obtenue à partir d’une glande salivaire de chironome (Chironomus sp.) permet d’apprécier la taille relative d’un ARN messager (long filament occupant toute la largeur de l’image), des ribosomes qui le décodent (boules jaunes le long de l’ARNm) et des protéines ainsi produites (courts filaments émergeant des ribosomes).
Cette image, obtenue à partir d’une glande salivaire de chironome (Chironomus sp.) permet d’apprécier la taille relative d’un ARN messager (en rose), des ribosomes qui le décodent (en bleu) et des protéines ainsi produites (en vert).
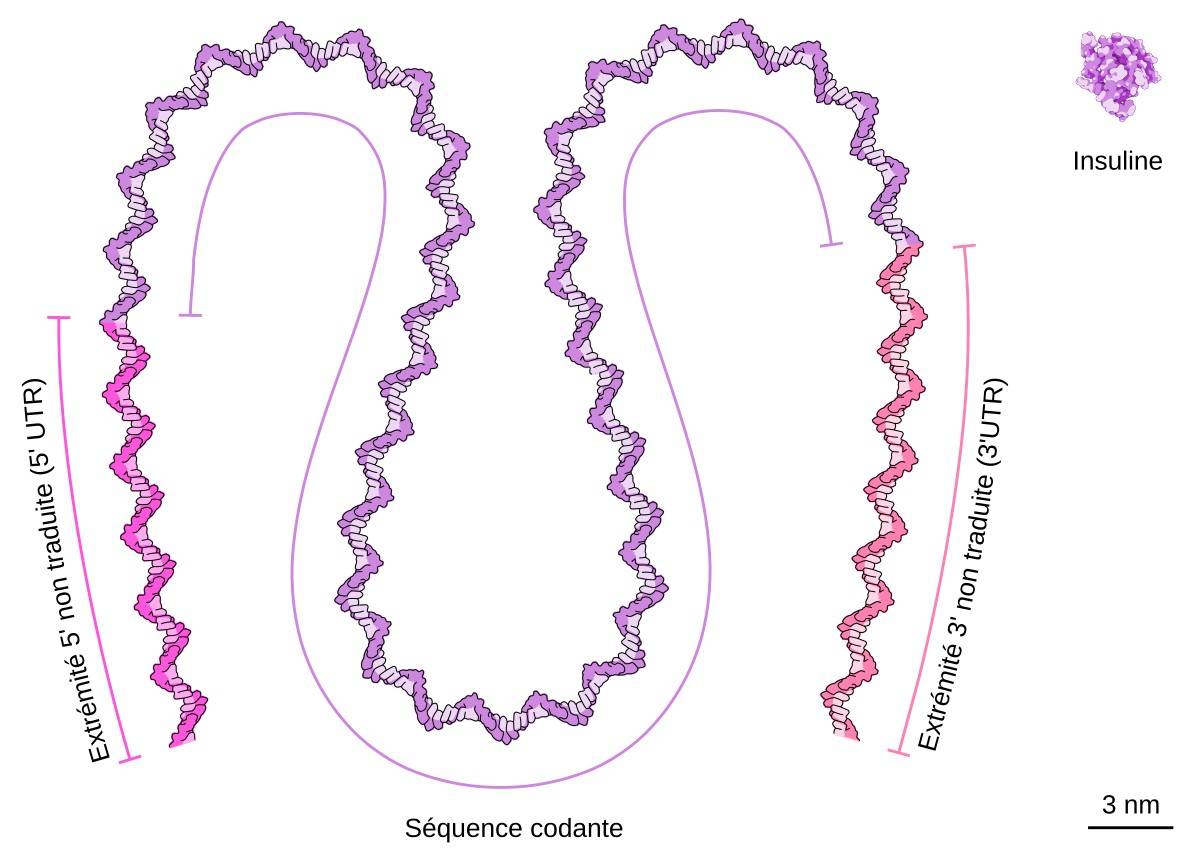
L’insuline est une petite protéine (51 acides aminés) codée par un ARNm relativement court (465 nucléotides se décomposant en 59 nucléotides à l’extrémité 5’ UTR, 333 nucléotides au niveau de la séquence codante et 73 nucléotides à l’extrémité 3’UTR). La queue polyA n’est pas représentée. La traduction de l’ARNm donne la prépro-insuline, longue de 110 acides aminés, qui est ensuite maturée en insuline (51 acides aminés). NB : L’ARN est ici représenté sous forme simple brin, sans former de structure secondaire, ce qui n’est probablement pas le cas physiologiquement, où les ARN messagers ne sont pas nus, mais associés à de nombreuses protéines. Le schéma présenté ici ne vise donc pas à rendre compte de la structure réelle de l’ARNm, mais uniquement de sa différence de taille par rapport à la protéine qu’il code.
Conclusion
Il ne s’agit bien sûr pas ici de remettre en cause la pertinence de schémas fonctionnels comme celui présenté Figure 1. Cependant, de tels schémas gagneraient à être complétés par des images obtenues en microscopie électronique à transmission (Figures 6 et 7), ou encore par des schémas dans lesquels les tailles relatives des différents éléments sont respectées (Figure 8, qui présente cependant comme limite de ne pas donner la structure physiologique de l’ARN messager, et vise uniquement à rendre compte de la différence de taille entre ARNm et protéine). De telles représentations traduisent en effet mieux le volume réellement occupé par les différentes molécules.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}