Effets de l'ajout d'un antibiotique sur les fréquences de tricheurs et de coopérateurs dans les populations de bactéries Pseudomonas aeruginosa.
Coopération microscopique : la production de biens communs chez les bactéries
Loin d’être des individus solitaires, les bactéries participent à une multitude d’interactions et vivent souvent en groupes que l’on appelle des biofilms. Elles produisent notamment des molécules qui sont secrétées dans le milieu extérieur où elles deviennent accessibles à toutes les bactéries capables de les utiliser, qu’elles soient ou non de la même espèce. Lorsque ces molécules ont un effet bénéfique pour les bactéries, on les qualifie de biens communs (Figure 1). Il existe un large éventail de biens communs qui sont utilisés pour récupérer de la nourriture, survivre en conditions difficiles, coloniser de nouveaux habitats et se déplacer. Il arrive cependant, par hasard, qu’une mutation réduise ou stoppe totalement la production de biens communs chez un individu. Si cet individu est toujours capable d’utiliser les molécules produites par les autres, il possède alors un avantage car il bénéficie du bien commun sans participer à sa production donc sans en payer le coût métabolique (ou en payant un moindre coût). Lorsqu’un tel mutant est en compétition avec un individu producteur (qualifié de coopérateur), c’est le mutant qui a l’avantage et les chercheurs appellent ces mutants des tricheurs (Figure 1).
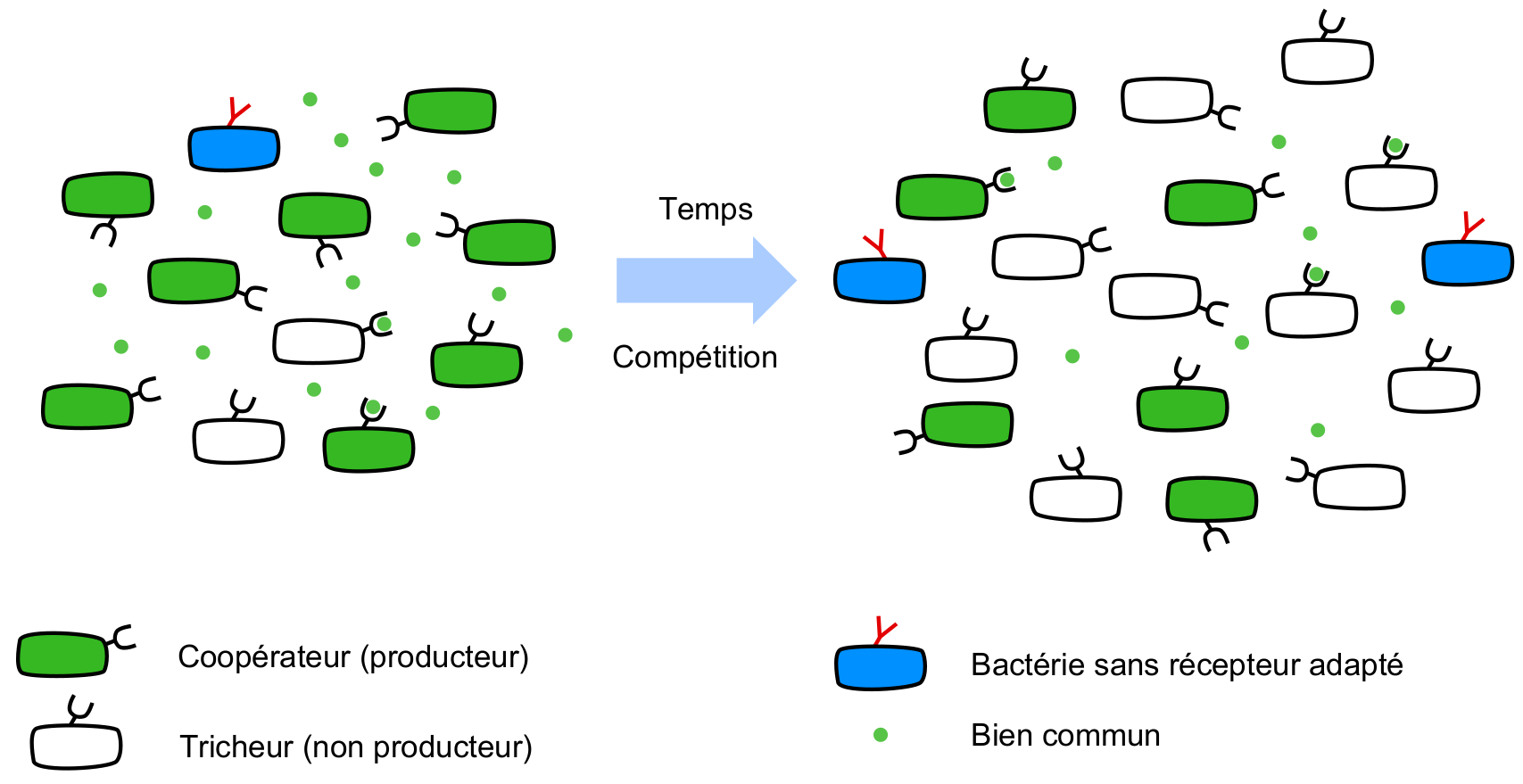
Les bactéries vertes sont les coopérateurs, elles produisent des biens communs et les utilisent (disques verts). Les bactéries blanches sont des tricheurs car elles ont le récepteur associé au bien commun mais ne le produisent pas. Les bactéries bleues ne peuvent utiliser le bien commun (récepteur triangle rouge). Après une période de croissance en compétition, les tricheurs (qui ont une croissance plus rapide) sont plus fréquents dans la population.
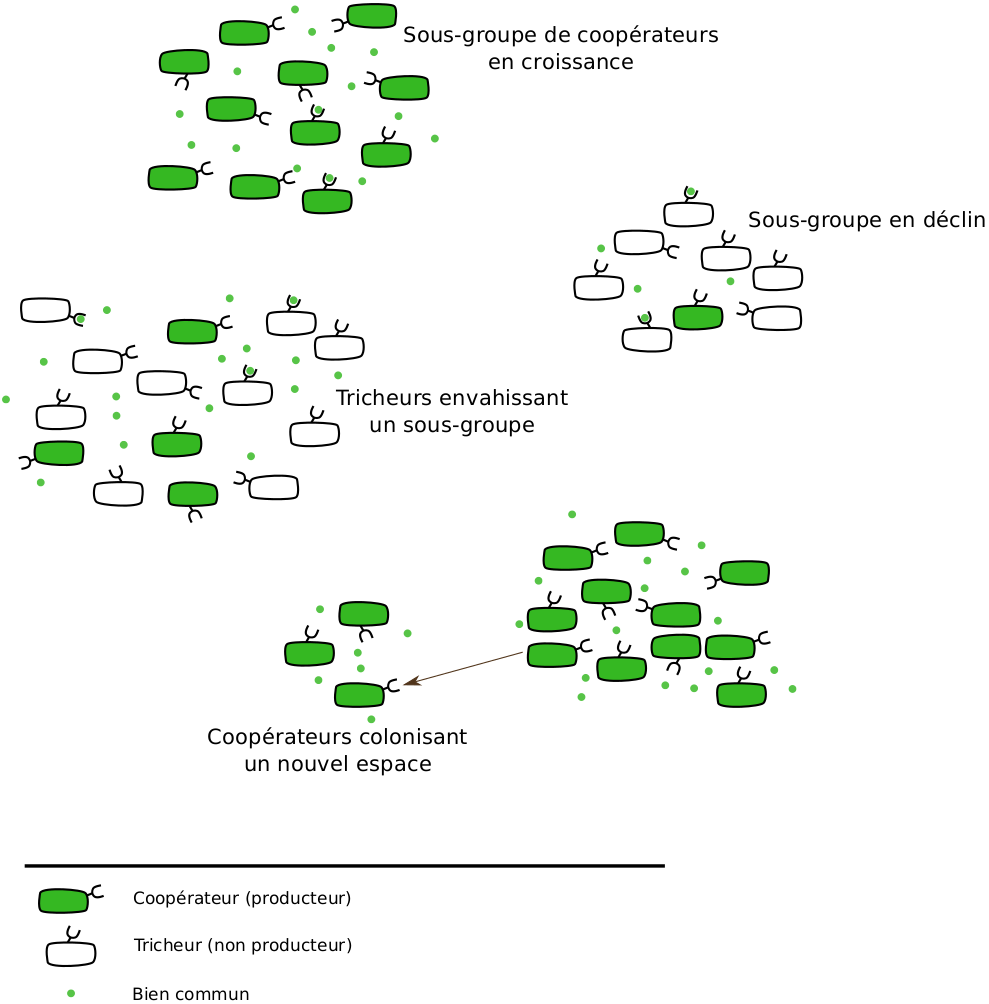
Mais alors, dans ce cas, comment peut-on observer des comportements coopératifs ? Pourquoi n’y a-t-il pas que des tricheurs ? Ce sont des questions qui ont fasciné (et qui fascinent toujours) des générations de scientifiques. Une des réponses prend la forme d’un apparent paradoxe. Imaginons une population avec des sous-groupes de différentes compositions en tricheurs et en coopérateurs. Au sein d’un sous-groupe, les coopérateurs sont désavantagés face aux tricheurs et voient leur fréquence diminuer. Cependant, si on considère la population dans son ensemble, la fréquence totale de coopérateurs peut augmenter. Cette expansion des coopérateurs, qui peut paraître surprenante, a été observée dans des études théoriques et expérimentales. Elle s’explique par la « performance » supérieure des sous-groupes avec davantage de coopérateurs. En effet, ces sous-groupes sont moins susceptibles de disparaître suite à une pénurie de biens communs et les individus en leur sein se reproduisent plus vite. Cet avantage global peut compenser voire supplanter le désavantage des coopérateurs à l’intérieur des sous-groupes et permettre le maintien de la coopération. De plus, l’extinction locale des sous-groupes comportant beaucoup de tricheurs crée de nouveaux espaces à occuper qui pourront être colonisés par des coopérateurs (Figure 2).
Face à l’ennemi : coopérer ou trahir ?
Les scientifiques qui se sont penchés sur la question du maintien de la coopération bactérienne ont élaboré de nombreuses expériences mettant en jeu des tricheurs et des coopérateurs. On sait par exemple que les tricheurs ont souvent un avantage plus grand lorsqu’ils sont en minorité que lorsqu’ils sont très fréquents dans un groupe, notamment parce les coopérateurs ne sont plus assez nombreux pour subvenir aux besoins de tous. Ce que l’on connaît moins, c’est comment ces interactions tricheurs-coopérateurs sont modifiées par l’environnement dans lequel elles ont lieu. Ainsi, les bactéries sont très fréquemment confrontées à des stress ou des ennemis tels que des prédateurs, des compétiteurs ou des antibiotiques qui ont des conséquences dramatiques sur la dynamique des populations. Pourtant, on sait très peu de choses sur la manière dont ces stress affectent l’avantage dont bénéficient les tricheurs, en particulier lorsque la coopération n’est pas directement liée à un mécanisme de résistance ou de tolérance à ces stress. C’est précisément ce que nous avons voulu étudier.
Notre expérience : la production de biens communs en présence d’antibiotiques
Nous avons utilisé une bactérie appelée Pseudomonas aeruginosa (voir Encadré 1) qui est une des stars des expériences sur la coopération. Cette bactérie produit des molécules appelées sidérophores qui s’attachent au fer environnemental. Le fer est un élément important du métabolisme bactérien et il se trouve rarement sous une forme directement disponible dans l’environnement. Les sidérophores se lient au fer pour le rendre utilisable et permettent donc aux bactéries de subvenir à leurs besoins en fer. La production de sidérophores est un exemple classique pour étudier la production de biens communs chez les bactéries. En effet, le sidérophore principal de P. aeruginosa, la pyoverdine, a la propriété d’être fluorescent et sa concentration dans le milieu est donc mesurable avec un appareil adapté, appelé spectro-fluorimètre. La pyoverdine donne également une couleur vert-jaune aux colonies sur boîtes de Pétri ce qui permet de reconnaître les tricheurs des coopérateurs. Dans notre expérience, les tricheurs ne produisent pas du tout de pyoverdine et apparaissent donc comme des colonies blanches sur les boîtes de Pétri.
Encadré 1 - Notre modèle biologique : Pseudomonas aeruginosa
Pseudomonas aeruginosa est une bactérie commune dans les sols et les milieux humides et qui peut, si elle en a l’opportunité, infecter des hôtes et notamment des humains. C’est donc une bactérie pathogène opportuniste. Elle est particulièrement dangereuse chez les patients immunodéprimés et ceux atteints de mucoviscidose. Chez ces derniers, elle se développe dans les poumons et peut conduire au décès du patient.
P. aeruginosa est fréquemment exposée à des antibiotiques, que ceux-ci soient d’origine humaine lorsqu’il s’agit de traiter une infection, ou microbienne quand les antibiotiques sont des médiateurs de compétition dans les communautés. De plus, on ne connaît aucun lien entre la résistance aux antibiotiques et la pyoverdine, ce qui nous a permis d’explorer les liens entre coopération et résistance dans un contexte où la coopération ne permet pas directement de résister aux antibiotiques. Notre question de recherche était double : comment la présence d’antibiotiques modifie les interactions entre tricheurs et coopérateurs ? et comment ces interactions sociales affectent l’évolution de la résistance aux antibiotiques ?
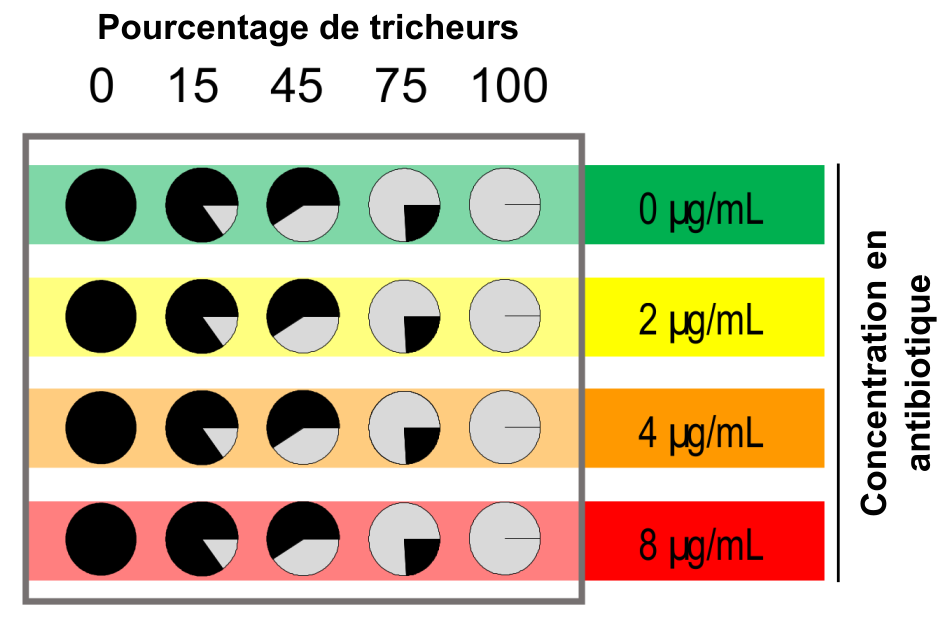
Des populations formées de différents pourcentages d’individus coopérateurs (en noir) et de tricheurs (en gris) ont été cultivées avec différentes concentrations d’antibiotique. Ces différentes populations ont ensuite été échantillonnées à t = 0, 10, 24, 34 et 48 heures.
Pour y répondre, nous avons préparé cinq types de populations (Figure 3) : des monocultures de tricheurs, des monocultures de coopérateurs et trois cultures mixtes avec respectivement 15 %, 45 % et 75 % de tricheurs. Les cultures étaient agitées en permanence pour homogénéiser l’environnement. Ces populations ont été exposées à des concentrations de 2 µg/mL, 4 µg/mL ou 8 µg/mL d’antibiotique ou cultivées sans antibiotique. Nous avions déterminé au préalable qu’une concentration de 10 µg/mL empêchait la croissance des bactéries (voir Encadré 2). Les concentrations utilisées dans notre expérience ne sont donc pas létales et l’antibiotique agit comme un bactériostatique c’est-à-dire qu’il empêche les bactéries de se reproduire sans les tuer.
Encadré 2 - Estimer le niveau de résistance des bactéries aux antibiotiques
Une manière courante de mesurer le niveau de résistance des bactéries consiste à déterminer la concentration à partir de laquelle la croissance d’une population est nulle. On appelle cette concentration la concentration minimale inhibitrice (CMI, ou MIC en anglais pour minimum inhibitory concentration). Classiquement, on soumet des populations de bactéries à des concentrations croissantes d’antibiotiques en multipliant par deux la dernière concentration testée. Si je commence par 1 µg/mL et que je souhaite tester 10 concentrations par exemple, je vais utiliser les concentrations 1, 2, 4, 8, 16, 32, 64, 128, 256 et 512 µg/mL, sans oublier d’avoir un témoin sans antibiotique pour vérifier que mes populations croissent correctement. Après une période de croissance (qui dépend du type de bactérie, chez P. aeruginosa c’est en général 24 heures), je mesure la densité des populations et la compare avec la densité initiale. La CMI correspond à la concentration la plus faible pour laquelle je mesure une croissance nulle.
Nous avons échantillonné ces populations au début de l’expérience et après 10 heures, 24 heures, 34 heures et 48 heures. Ces échantillons nous permettent d’estimer la densité totale de bactéries dans chacune des populations ainsi que la proportion de tricheurs et de coopérateurs (Encadré 3). Au début et à la fin de l’expérience, nous avons également étalé nos échantillons sur des boîtes contenant du milieu nutritif et l’antibiotique. Dans ce cas, seules les bactéries résistantes se sont reproduites et ont formé des colonies. En faisant le ratio de la densité de colonies résistantes (sur les boîtes avec antibiotique) sur la densité de colonies totales (sur les boîtes sans antibiotique), on peut calculer la proportion de bactéries résistantes.
Encadré 3 - Estimer la densité d’une population bactérienne par étalement sur boîtes de Pétri
Pour estimer une densité de bactéries par étalement, on effectue des dilutions en série d’échantillons des populations et on étale les différentes dilutions sur un milieu nutritif dans des boîtes de Pétri. Après 24 heures, on observe des colonies et on fait l’hypothèse que chaque colonie provient d’une bactérie unique qui s’est ensuite reproduite dans la boite et qui forme une colonie visible à l’œil nu. On appelle ces colonies des « unités formant colonies » (ou CFU en anglais pour colony forming unit). On réalise des dilutions en série pour obtenir un nombre de colonies par boîte de Pétri qui soit suffisamment grand pour limiter les effets stochastiques et suffisamment petit pour qu’il n’y ait pas de chevauchement entre colonies. En général, on choisit des boîtes qui contiennent entre 30 et 300 colonies. En comptant ces colonies et en prenant en compte le facteur de dilution, on estime la densité de chaque population c’est-à-dire le nombre de bactéries par millilitre de milieu.
Les antibiotiques favorisent les tricheurs… ?
Notre premier objectif était d’étudier comment les antibiotiques affectent les interactions sociales dans les populations de bactéries. Dans toutes les populations mixtes, c’est-à-dire contenant à la fois des coopérateurs et des tricheurs, c’est la fréquence de ces derniers qui augmente au cours de l’expérience, et ce quelle que soit la concentration d’antibiotique dans l’environnement. Ce que montre la figure 4, cependant, c’est qu’en présence d’antibiotiques la fréquence de tricheurs augmente encore plus vite que sans antibiotique. Les antibiotiques modifient donc les interactions entre coopérateurs et tricheurs en accentuant l’avantage des tricheurs.

Chaque panneau correspond à une proportion initiale de tricheurs et les couleurs représentent les différentes concentrations d’antibiotiques.
Notre hypothèse pour expliquer ce résultat est que les coopérateurs sont plus vulnérables que les tricheurs aux antibiotiques. Autrement dit, les antibiotiques auraient un effet plus délétère sur les coopérateurs que sur les tricheurs. En effet, la production de biens communs a un coût métabolique et les coopérateurs ont ainsi moins de ressources que les tricheurs à investir dans la résistance aux antibiotiques. Les antibiotiques constituent alors un coût supplémentaire pour les coopérateurs qui se manifeste par un bénéfice accru pour les tricheurs (Figure 5).
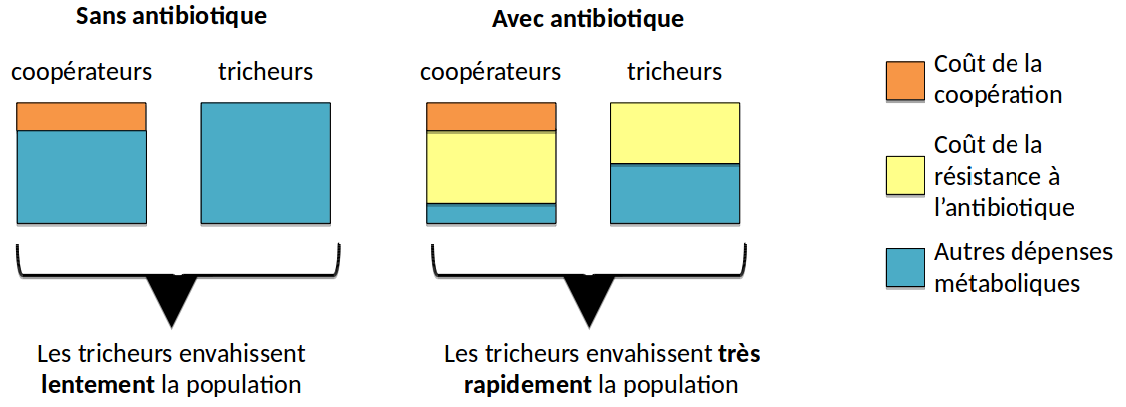
Chaque carré représente l’ensemble des dépenses métaboliques d’un type de bactérie. Les tailles relatives des zones colorées ne sont pas à l’échelle et servent à montrer les différences entre tricheurs et coopérateurs.
Notre expérience montre ainsi que les interactions sociales peuvent être considérablement modifiées par des environnements stressants. Des stress tels que les antibiotiques augmentent l’avantage des tricheurs, rendant encore plus épineuse la question du maintien et de l’évolution de la coopération dans les populations bactériennes. La structure des populations, qui fait que tout le monde n’interagit pas avec tout le monde, joue certainement un rôle crucial. Comme nous l’avons expliqué précédemment, les groupes où il y a beaucoup de coopérateurs se portent mieux que les groupes comportant beaucoup de tricheurs. Ce que nous observons dans notre expérience s’apparente aux interactions qui ont lieu au sein de tels groupes. On peut donc imaginer que les antibiotiques accélèrent le déclin des groupes de tricheurs au profit des groupes de coopérateurs et contribuent au maintien global de la coopération.
Évolution de la résistance à l’antibiotique dans les populations bactériennes
Notre second objectif était d’examiner l’évolution de la résistance bactérienne et l’influence des interactions sociales sur la résistance.
À la fin de l’expérience, la fréquence de bactéries résistantes dans les populations est d’autant plus grande que les bactéries ont été soumises à une concentration d’antibiotiques élevée. En d’autres termes, la part de bactéries sensibles à l’antibiotique est plus faible lorsque que la concentration d’antibiotique est élevée. Cela signifie que les bactéries résistantes ont un plus grand avantage sur les bactéries sensibles en présence des concentrations plus élevées d’antibiotiques. Il faut ici se souvenir que nous avons utilisé des concentrations non létales et que, par conséquent, les bactéries sensibles ne sont pas tuées par l’antibiotique.
Ces bactéries résistantes sont issues d’au moins une bactérie mutante qui était soit initialement présente dans la population soit qui est apparue au cours de l’expérience. Dans les deux cas, si on fait l’hypothèse que le taux de mutation est le même chez les coopérateurs et les tricheurs, c’est le type le plus nombreux qui aura la plus grande probabilité de contenir cette bactérie mutante. Par exemple, si chaque bactérie a une chance sur 10 000 d’avoir une mutation qui confère une résistance, dans un groupe de 1000 bactéries cette mutation apparaîtra avec une probabilité de 1000 × (1/10 000) = 0,1 alors que dans un groupe de 100 bactéries cette probabilité sera de 100 × (1/10 000) = 0,01.
Pour autant, la fréquence de bactéries résistantes dans la population ne dépend pas uniquement de la probabilité d’apparition d’une mutation qui confère une résistance. La fréquence de bactéries résistantes dépend également de la vitesse à laquelle ces bactéries se reproduisent. Ainsi, si une population contient par hasard un tricheur résistant et un coopérateur résistant au début de l’expérience, à la fin l’expérience il y aura davantage de tricheurs résistants que de coopérateurs résistants car les tricheurs se reproduisent plus vite que les coopérateurs (car ils ne paient pas le coût de la coopération).
Dans les populations mixtes, les tricheurs croissent non seulement plus vite que lorsqu’ils sont seuls, car ils ont accès au bien commun, mais ils sont aussi plus rapides que les coopérateurs en monocultures, car ils ne payent pas le coût de production du bien commun. Par conséquent, ce sont les populations mixtes qui contiennent les fréquences de bactéries résistantes les plus élevées et celles-ci sont en très grande majorité des tricheurs. Par contre, s’il y a initialement beaucoup de tricheurs dans la population, la fréquence finale de résistants n’est pas plus élevée dans les populations mixtes que lorsqu’ils sont tous seuls. En effet, la production de bien commun par la faible minorité de coopérateurs n’est probablement pas suffisante pour subvenir aux besoins de toute la population et celle-ci s’apparente presque à une monoculture de tricheurs.
Points-clés :
- Les populations bactériennes ne sont pas toujours homogènes et peuvent contenir, par exemple, des tricheurs et des coopérateurs.
- Des doses non létales d’antibiotiques augmentent l’avantage des tricheurs en compétition.
- Les populations mixtes contenant des tricheurs et des coopérateurs évoluent davantage de résistance que les monocultures.
Pour finir : des bactéries tricheuses ?
Si l’on parle de tricheurs et de coopérateurs pour décrire des comportements de bactéries c’est avant tout pour se faire comprendre en utilisant un vocabulaire imagé. Nous ne prêtons pas d’intentions, qu’elles soient bonnes ou mauvaises, aux bactéries. De plus, ce que nous appelons un coopérateur ou un tricheur se focalise sur un trait de coopération particulier, dans des conditions environnementales données. Le même individu peut donc être qualifié de tricheur dans un environnement et de coopérateur dans un autre. De même, il est tout à fait possible qu’un tricheur pour la pyoverdine produise davantage d’un autre bien commun auquel nous ne nous sommes pas intéressés.
Référence
Marie Vasse, Robert J. Noble, Andrei R. Akhmetzhanov, Clara Torres-Barceló, James Gurney, Simon Benateau, Claire Gougat-Barbera, Oliver Kaltz, and Michael E. Hochberg
Antibiotic stress selects against cooperation in the pathogenic bacterium Pseudomonas aeruginosa
PNAS 2017 114 (3) 546-551; published ahead of print January 3, 2017, doi:10.1073/pnas.1612522114