

Après avoir présenté l'ensemble des phénomènes à l'origine d'une variation génétique dans une population, à l'échelle du génome puis du gène, cet article explique la notion de dérive génétique, les facteurs qui l'influencent, ses implications dans la diversité allélique d'une espèce et les conséquences sur la biodiversité.
Toutes les espèces présentes à l’heure actuelle sur notre planète sont caractérisées par une information génétique, contenue dans leur génome, qui leur est propre. Cela dit, le génome de ces espèces n’est pas figé. Divers mécanismes comme les duplications, les fusions ou les cassures de chromosomes, les mutations géniques (insertions, délétions, substitutions…) contribuent à faire évoluer ces génomes, conduisant, à terme, à partir d’une espèce ancestrale, à l’apparition de groupes d’individus qui, du fait des nombreuses modifications au sein de leur génome, ne sont plus interféconds et constituent donc de nouvelles espèces. C’est le phénomène de spéciation qui ne se produit pas à l’échelle de la vie humaine mais sur des centaines de milliers d’années.
Évolution des génomes
Duplications génomiques
Deux types de duplication génomique sont discernables, dépendant du mécanisme mis en jeu pour leur apparition.
-
S’il y a non-disjonction des chromosomes de la lignée germinale en méiose, cela va permettre la création de gamètes diploïdes à 2n chromosomes (au lieu de n normalement). S’il y a fusion entre deux gamètes de ce type, on va obtenir un zygote à 4n chromosomes qui aura donc subi un doublement complet de son génome. C’est ce que l’on appelle l’autopolyploïdie.
-
Lors de l’hybridation interspécifique entre 2 espèces donnant 2 lots chromosomiques distincts, le nombre de chromosomes de l’hybride F1 est alors la somme des chromosomes de chaque espèce. Il y aura création d’une nouvelle espèce si l’hybride est fertile, c’est l’allopolyploïdie (voir figure ci-dessous).
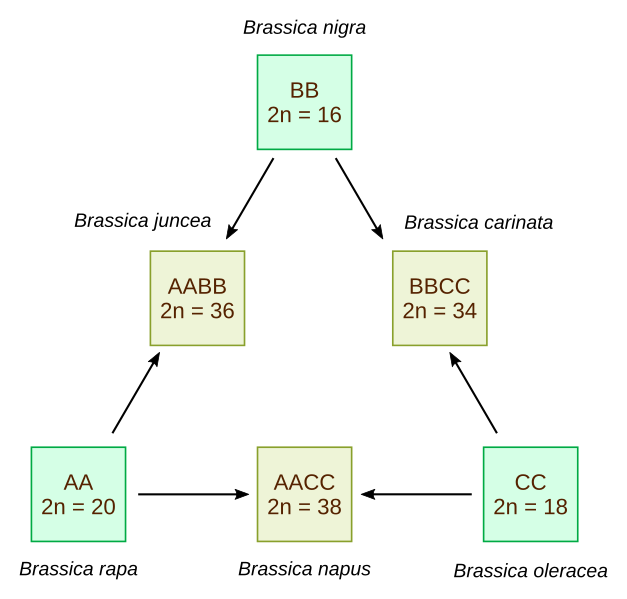
Le "triangle de U" décrit les relations génétiques entre 3 espèces de Brassica diploïdes (B. oleracea, B. rapa et B. nigra) et 3 espèces allotétraploïdes (B. napus, B. juncea et B. carinata) résultant d'hybridations entre les diploïdes (hypothèse publiée par Woo Jang-choon en 1935 et confirmée ensuite par des études portant sur l'ADN et les protéines).
Brassica nigra = moutarde noire, Brassica carinata = moutarde d'Éthiopie, Brassica juncea juncea = moutarde brune, Brassica oleracea botrytis = chou-fleur, Brassica rapa rapa = navet, Brassica napus napus = colza.
Adapté d'après Adenosine, Wikimedia.
Duplications segmentales et géniques
Plusieurs mécanismes cellulaires comme les mécanismes de réparation suite à une cassure de l’ADN, les rétrotranspositions d’éléments transposables (séquences virales contenues dans nos génomes qui ont gardé la faculté de se dupliquer puis de s’insérer ailleurs dans le génome), les enjambements inégaux au niveau de chiasmas méiotiques (échange non équilibré de matériel génétique entre deux chromosomes homologues menant à la duplication d’une séquence sur un des chromosomes et à sa délétion sur l’autre) peuvent induire une duplication d’une région génomique (duplication segmentale) ou de tout ou partie d’un gène.
Comparaison entre espèces
Du fait de ces divers mécanismes, deux espèces ayant divergé à partir d’un ancêtre commun vont donc avoir gardé certaines similitudes au niveau de leurs chromosomes mais vont également accumuler des dissemblances. La figure ci-dessous compare par exemple les structures chromosomiques de deux génomes murin (de la souris) et humain qui ont évolué à partir d’un même ancêtre commun. Les blocs de chromosomes transmis sans modification d’une espèce à l’autre sont appelés bloc de synténie.
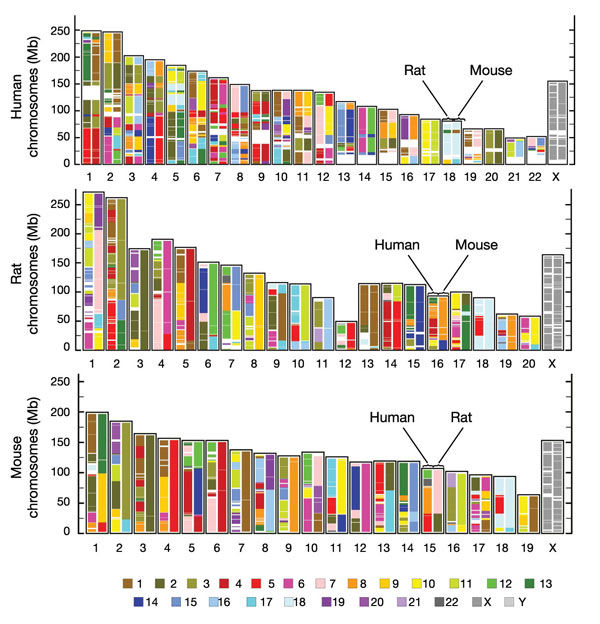
Ces cartes représentent la répartition des régions conservées, c'est-à-dire des zones de synténie (synténie = conservation de l'ordre des gènes) entre les génomes de l'homme, du rat et de la souris. Une couleur a été attribuée à chaque chromosome humain (en bas de la figure) et les localisations des fragments chromosomiques humains (zones de synténie humaines), sont indiquées dans les chromosomes du rat et de la souris, selon ce code couleur. On peut voir que le chromosome 1 humain se retrouve, par exemple dans le génome murin, en quatre fragments distincts, au niveau des chromosomes 1,3 et 4.
Évolution des gènes
Jusqu’à présent nous avons surtout évoqué les modifications concernant le nombre et la constitution des chromosomes ou des séquences géniques. Il ne faut toutefois pas négliger un autre moteur important de l’évolution des génomes, la mutation au sein des gènes.
Notion d’allélisme
Les gènes présents au sein des génomes peuvent évoluer, se modifier du fait des forces évolutives par des mécanismes aléatoires (mutations, recombinaisons…) et d’autres non aléatoires (sélection naturelle sexuelle, sélection artificielle…). La modification de ces gènes va entraîner l’apparition de plusieurs séquences nucléotidiques (l’enchaînement des nucléotides au sein d’un chromosome est caractéristique pour chaque gène) très similaires, mais qui présentent de petites variations au sein d’une population, les allèles.
La dérive génétique
Les différents allèles d’un gène n’apparaissent pas en même temps durant l’évolution d’une espèce. De plus, ils peuvent conférer un avantage ou un désavantage aux individus qui les portent. Leurs fréquences vont donc changer au cours du temps.
De manière générale, l’ensemble des mécanismes décrits comme étant des forces évolutives (sélection naturelle, mutation, recombinaison, dérive, migration…) contribue à faire varier les fréquences alléliques des gènes. Pour la plupart de ces forces, un mécanisme moléculaire est mis en jeu. Mais la dérive génétique concerne l’évolution au sein d’une population, ou d’une espèce, de la fréquence des allèles ou des génotypes (combinaison des deux allèles d’un gène héritée des chromosomes paternel et maternel au moment de la fécondation) d’un gène, causée par des phénomènes aléatoires et impossibles à prévoir, donc indépendamment des mutations, de la sélection naturelle et des migrations. Citons pour exemple le hasard des rencontres des spermatozoïdes et des ovules (panmixie) dans le cas d’une reproduction sexuée.
Les effets de la dérive génétique sont d’autant plus importants que la population est petite, car les écarts observés d’une génération à l’autre par rapport aux fréquences alléliques y sont d’autant plus perceptibles. La dérive génétique concerne surtout les allèles neutres, qui ne confèrent ni avantage ni désavantage sélectif. La dérive génétique est un des mécanismes majeurs de l’évolution.
Principe de la dérive génétique
Alors que dans des populations de taille infinie, en l’absence de sélection et de mutation, les fréquences alléliques sont stables au cours des générations (principe de Hardy-Weinberg, voir « Pour aller plus loin »), elles vont varier aléatoirement dans des populations diploïdes (possédant deux copies complètes de leur génome) de taille finie. Ceci est dû à la variabilité du tirage aléatoire des allèles d’une génération à l’autre, résultant du mécanisme de formation des gamètes.
Principe de Hardy-Weinberg
Le principe de Hardy-Weinberg porte sur l’existence d’un équilibre des fréquences alléliques et génotypiques au cours des générations, pour une population. Afin que cet équilibre puisse exister cette population doit suivre certains critères bien précis
-
La population doit être de taille infinie (dans la pratique à partir de 1000 individus on peut approximer la taille de la population comme infinie)
-
L’espèce dont est issue la population doit être diploïde
-
L’espèce dont est issue la population doit avoir une reproduction sexuée
-
Les croisements entre les individus au sein de la population doivent se faire au hasard (principe de panmixie)
-
Il ne doit pas y avoir de forces évolutives telles que la mutation, la migration ou la sélection en jeu dans cette population
Dans ces conditions, pour un gène possédant deux allèles différents, les allèles A et a, de fréquences respectives p et q, la loi de Hardy-Weinberg postule que les fréquences des génotypes homozygotes AA (individus possédants deux fois la forme « A » du gène dans leur génome), aa (individus possédants deux fois la forme « a » du gène dans leur génome) et hétérozygote Aa (individus possédants les deux variants « A » et « a » du gène dans leur génome) peuvent être estimées comme suit :
- f(AA) = p2
- f(Aa) = 2pq
- f(aa) = q2
Lors de la fécondation, la rencontre de deux gamètes haploïdes paternel et maternel va donner naissance à un nouvel individu diploïde dont la totalité du génome est présent sous forme de deux copies ; paternelle et maternelle. Si pour un gène donné les deux copies provenant des deux parents sont identiques, on dit que l’individu est homozygote. Par contre si ces deux copies sont des variants alléliques différents, l’individu est dit hétérozygote.
C’est lors de la séparation des chromosomes homologues qu’il va y avoir une répartition, un tirage, aléatoire des chromosomes allant dans chaque gamète. En se basant sur ce mode de formation des gamètes, un individu ne se reproduisant qu’une fois, ne va transmettre à son descendant qu’un seul des deux allèles de chacun de ses gènes. Pour qu’il puisse transmettre à coup sûr la totalité de ses allèles, il faudrait que son nombre de descendants tende vers l’infini (d’où la condition de taille infinie dans la loi de Hardy-Weinberg, voir « Pour aller plus loin »). A l’échelle d’une population, il est donc statistiquement inévitable que certains allèles ne soient pas transmis d’une génération à l’autre. Ce phénomène est encore amplifié si l’on tient compte du fait que certains adultes n’auront pas de descendance.
Les fréquences alléliques d’un gène vont donc varier d’une génération à l’autre, certains allèles voyant leur fréquence diminuer ou augmenter au sein de la population. Ces variations peuvent aller jusqu’à la disparition de certains allèles, ce qui a pour conséquence directe la diminution du nombre d’allèles du gène dans la population.
La dérive génétique est donc un mécanisme aléatoire (stochastique) qui va modifier la fréquence des allèles d’un gène dans la population en l’augmentant ou en la diminuant. En fonction de la taille de la population et de la fréquence des allèles à l’origine, le temps de fixation (temps que vont mettre les allèles pour atteindre un point d’équilibre avec un nombre d’allèles fixé et des fréquences alléliques stables) va varier (voir figure ci-dessous). L’effet de taille de la population est particulièrement visible lorsqu’un faible groupe d’individus d’une espèce va coloniser un autre milieu. Plus la taille de la population est faible, plus la dérive est rapide. A plus ou moins long terme la dérive génétique va entraîner une diminution de la variabilité génétique (voir ci-après).
Les deux figures ci-dessous donnent une idée du temps de fixation d'un allèle dans une population en fonction de taille de celle-ci, une population de 10 individus pour la première figure et de 1000 individus, pour la deuxième.
Elles sont extraites d'une animation en ligne permettant de simuler les évolutions des fréquences des deux allèles A1 et A2 d'un hypothétique gène A. Chaque figure présente deux repères, portant en abscisse le temps, en nombre de génération, et en ordonnée, soit la fréquence de l'allèle A1 (repère du haut), soit celle de l'allèle A2 (repère du bas), qui se déduit aisément de celle de A1. Chacun des deux graphiques ci-dessous présente 5 exemples d’évolution des fréquences alléliques (représentés par 5 courbes de couleurs différentes), au cours de 200 générations . Ces courbes ont été obtenues par simulation informatique à partir d’une valeur initiale de 0,5 pour la fréquence de l’allèle A1 et donc également 0,5 pour la fréquence de l'allèle A2. Dans ces simulations deux autres paramètres sont fixés : les avantages selectifs des différents génotypes (fitness) sont identiques et les taux de mutation des deux allèles sont nuls.
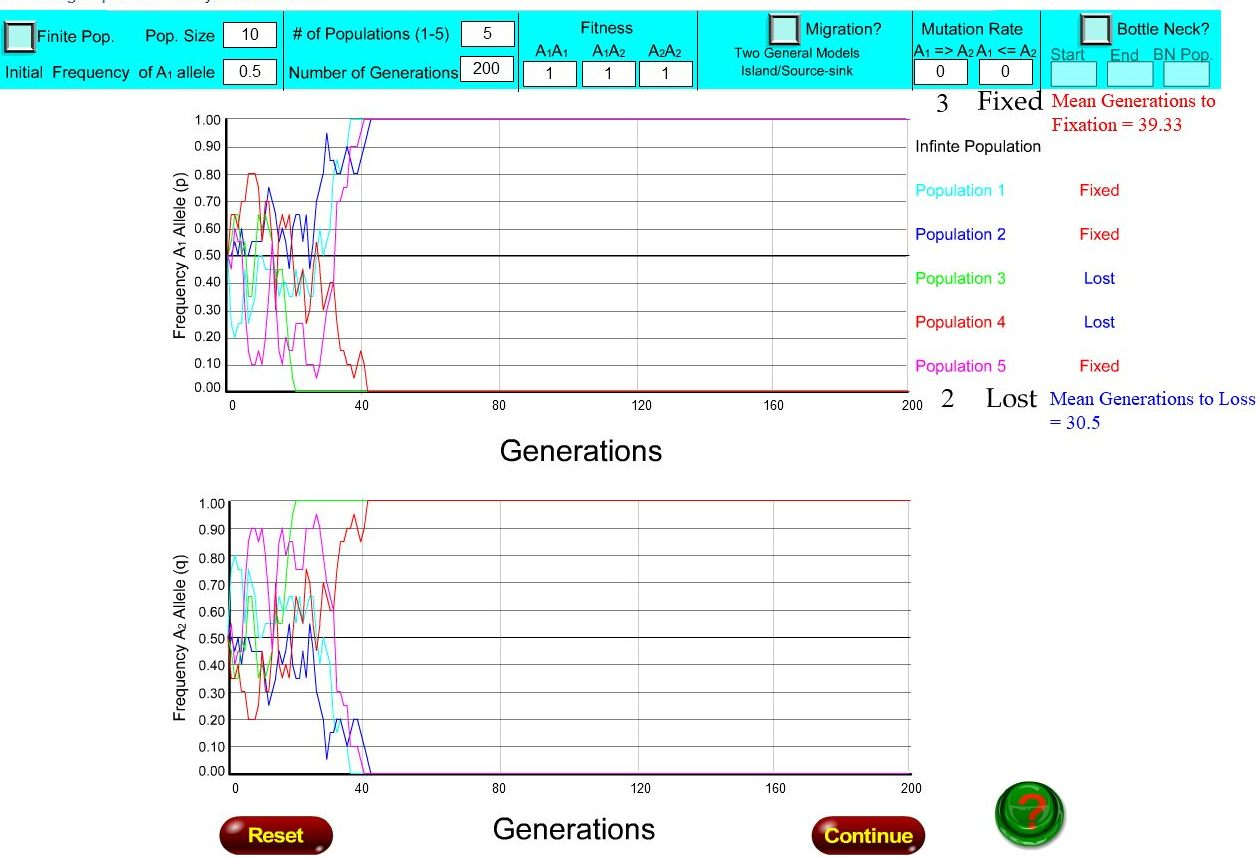
Sur deux repères représentant les fréquences des allèles A1 et A2 du gène A, les courbes de couleur correspondent à 5 simulations différentes effectuées pour une population de 10 individus, sur 200 générations. Ces deux allèles ont une fréquence de 0,5 au départ, et la somme des deux fréquences étant toujours égale à 1, les courbes de couleur du graphique du haut (A1) et du bas (A2) sont complémentaires (fréquence de A1 = p et fréquence de A2 = 1-p). Les simulations montrent, selon les cas (courbes de différentes couleurs), que l'allèle A1, par exemple, va rapidement, en 40 générations, disparaître (fréquence égale à 0) ou être fixé (fréquence égale à 1), alors qu'à l'origine on lui avait donné une fréquence de 0,5. On note aussi que l'amplitude des changements de fréquence d'une génération à l'autre est alors très grande.
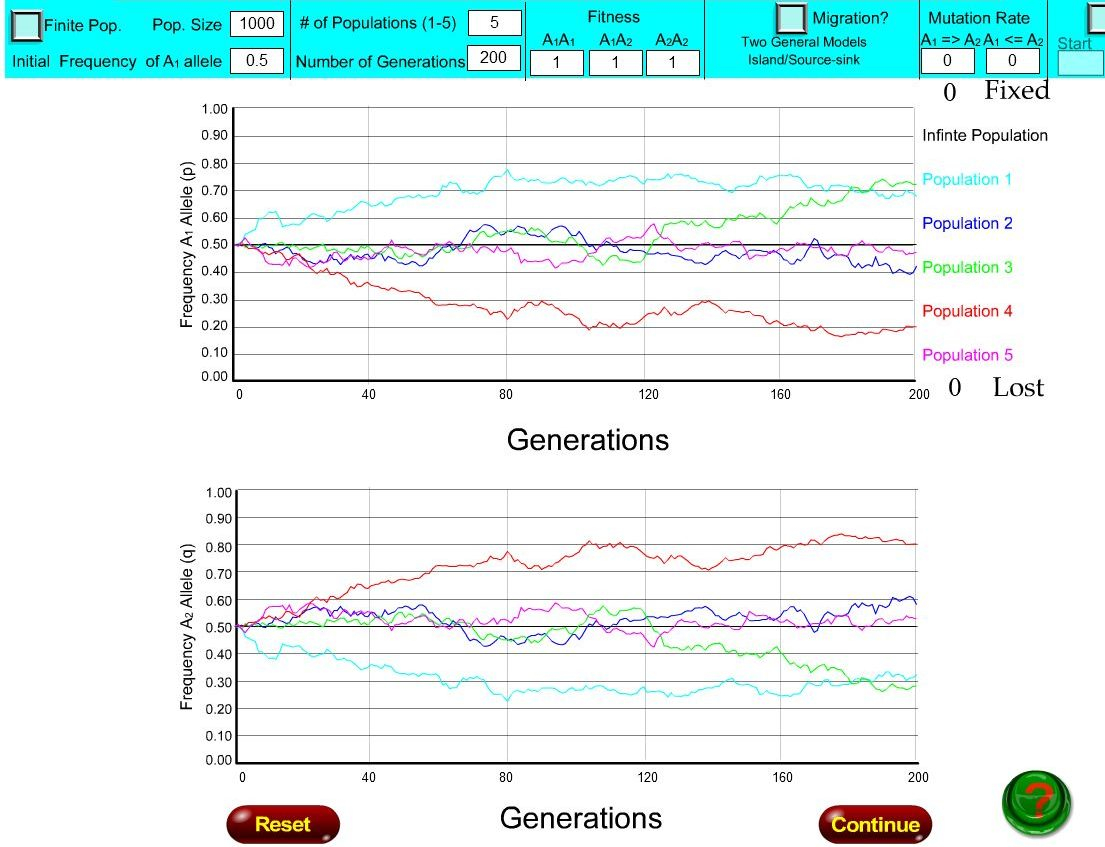
Ces 5 simulations sont effectuées avec les mêmes paramètres que dans la figure précédente, mais sur une population de 1000 individus.
Ces 5 simulations montrent une évolution globalement similaire : la fréquence de l'allèle A1 varie relativement faiblement (donc A2 aussi, mais en sens inverse) d'une génération à l'autre.
La fréquence allélique, au bout de 200 générations, tend vers une valeur stable qui n'est pas très éloignée de sa valeur de départ, l'allèle A1 n'a pas été fixé ni perdu.
De manière générale, plus la population est de taille importante moins le risque de disparition de certains allèles est grand. On tend alors vers un équilibre des fréquences alléliques et donc génotypiques (selon les prévisions de la loi de Hardy-Weinberg).
Dérive génétique et effet fondateur
Lorsqu’une sous-partie d’une population se sépare de la population initiale de taille beaucoup plus vaste, lors d’une migration pour coloniser un nouveau milieu par exemple, la population pionnière, ou fondatrice, n’est pas le reflet exact de la population de départ. Cette sous-population ne va « prendre » qu’un échantillon du pool d’allèles disponible dans la population mère « dans ses bagages » et ce, de manière aléatoire. Elle peut donc avoir des fréquences alléliques fort différentes de la population initiale. C’est ce que l’on appelle « l’effet fondateur ». Plusieurs exemples types d’effet fondateur ont été décrits dans l’histoire humaine, comme lors de la colonisation de l’Islande ou de la région québécoise du Saguenay-Lac-Saint-Jean.
Il est cependant difficile de déterminer la part de chaque force évolutive dans l’évolution des fréquences alléliques d’une population. Tous ces mécanismes vont agir conjointement et contribuer à cette évolution. Si nous reprenons l’exemple de la région québécoise du Saguenay-Lac-Saint-Jean, la population autochtone y souffre de certaines maladies génétiques (ataxie spastique, syndrome d’Andermann…) avec des fréquences beaucoup plus élevées que dans d’autres parties du monde. Pour comprendre l’origine de ces fréquences élevées, il faut étudier plus en détail l’histoire de cette population. La région a initialement été colonisée au XVIIe siècle par quelques milliers d’individus qui ont constitué la population fondatrice. Suite à ce début de colonisation, peu de nouveaux colons sont venus s’ajouter au groupe fondateur qui s’est alors reproduit durant un certain nombre d’années en « vase clos », entraînant une forte consanguinité. La consanguinité va intervenir sur les fréquences alléliques des gènes en amplifiant le taux d’homozygotie (tous les individus d’une famille consanguine sont issus du même ancêtre commun et ont de fortes chances de recevoir les mêmes allèles de cet ancêtre unique). En construisant les arbres généalogiques des familles dans lesquelles sévissent à l’heure actuelle ces maladies, on se rend compte que pour chacune il existe un ancêtre fondateur unique qui a introduit la mutation délétère. C’est uniquement par hasard que des porteurs de ces allèles délétères se sont trouvés dans la population fondatrice. Par la suite le mode de reproduction en milieu confiné, accroissant la consanguinité, a amplifié la fréquence de ces allèles délétères dans les familles. Ceci a entraîné l’apparition d’un nombre élevé d’individus porteurs de deux copies délétères du gène (individus malades).
Dérive génétique et érosion de la biodiversité
Les études de fixation d’allèles par dérive génétique ont montré que ce mécanisme peut aboutir à la fixation d’un allèle du gène au détriment des autres, dans une population. Cette constatation est particulièrement vraie pour de petits échantillons mais l’effet est de plus en plus limité lorsque la population s’accroît (comparer dans les figures ci-dessus les simulations pour 10 et 500 individus). Si l’on fixe un allèle dans la population cela va forcément entraîner au sein de celle-ci une baisse de la diversité génétique qui est pourtant essentielle à l’adaptabilité des espèces aux changements de milieu, de conditions de vie… Ainsi, plus une population va être petite, plus les effets de la dérive génétique seront importants et plus la diversité génétique dans la population sera menacée. La dérive génétique est donc un mécanisme défavorable à l’adaptation des espèces.
La perte de diversité génétique par dérive génétique se produit aléatoirement mais peut être amplifiée par des effets artificiels comme la réduction des effectifs (prédation, déforestation, utilisation agricole) ou la consanguinité.
Références
- Livre
Human molecular genetics, Strachan et Read, 3e édition. - Site
Génétique des populations appliquée aux données moléculaires (réseau GENET d’enseignement de la génétique)




{kind=link}