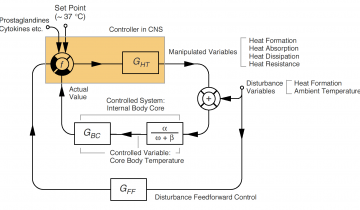
Article présentant les notions de base en cybernétique, permettant un meilleur enseignement de la physiologie. Ces principes sont ici appliqués au système de la régulation de la température et des variations hormonales chez l’homme et chez la femme. Les notions présentées peuvent également servir à l’enseignement de la régulation de la pression artérielle, de la glycémie…
Introduction
La cybernétique peut être définie comme l’ensemble des sciences du pilotage, c’est-à-dire les mécanismes commandant l’évolution d’un système vers un but défini. Ces mécanismes se rapportent donc à une notion de finalité, au sens cybernétique du terme : on dit qu’un système est finalisé quand il évolue vers un nouvel état antérieurement défini, quelles que soient les péripéties imprévisibles qui accompagnent cette évolution.
L’utilisation d’une approche cybernétique de l’étude de la physiologie est proposée dans le cadre des programmes de l’enseignement secondaire. Toutefois, ces notions restent inconnues pour bon nombre d’enseignants. Le présent article se propose donc d’introduire les grandes notions de cybernétique en physiologie animale, et d’illustrer ceci par trois exemples : le thermostat, exemple théorique simple permettant de comprendre ces notions, et l’axe gonadotrope chez l’homme et la femme.
Cet article s’appuie essentiellement sur l’ouvrage de B. Calvino, « Introduction à la physiologie – Cybernétique et régulations », publié chez Belin, collection Belin Sup Sciences (2003), qui aborde la physiologie animale sous l’angle de la cybernétique, à l’aide d’exemples variés.
En complément, vous pouvez également consulter cette réflexion sur l’emploi du terme rétroaction, vu sous une approche cybernétique.
Cybernétique et physiologie : les grands principes
Mise en relation de la cybernétique et de la physiologie
La cybernétique correspond aux sciences du pilotage. Il est ainsi possible d’étudier la physiologie d’un organisme par une approche cybernétique : Il s’agit là d’étudier comment évoluent les paramètres physiologiques de l’organisme, comment ils sont fixés.
Une approche cybernétique de la physiologie ne peut s’envisager qu’en rapport avec une modélisation des mécanismes physiologiques de l’organisme. En effet, une démarche de modélisation consiste à observer le fonctionnement d’un organisme, et ainsi mettre en évidence les paramètres qui décrivent cet organisme et leurs relations.
Pour modéliser la physiologie (et ainsi comprendre le fonctionnement de l’organisme), il est donc nécessaire d’utiliser un langage précis dans la description des relations entre les paramètres choisis, de leurs évolutions, etc. Comme nous allons le voir dans cet article, le langage cybernétique se révèle particulièrement adapté à cette tâche de modélisation.
Modélisation cybernétique : le concept de « boîte noire »
La démarche de modélisation consiste à construire un premier modèle, qui est progressivement corrigé et affiné par comparaison avec des résultats d’expérimentations. L’utilisation d’un langage cybernétique lors de cette démarche est à relier au concept de « boîte noire ». L’idée est de prendre en compte ce qui rentre et sort d’un organisme (ou d’un organe), sans se préoccuper des mécanismes précis qui ont lieu dans cet organisme (ou cet organe).
On étudie donc :
-
des paramètres d’entrée, décrivant les informations, molécules, paramètres physico-chimiques, etc., arrivant ou influençant l’activité de l’organisme,
-
et des paramètres de sortie, décrivant les informations, molécules émises par l’organisme, les paramètres physico-chimiques modifiés par l’organe, etc.,
-
ainsi que les relations entre ces paramètres.
Ces relations traduisent l’action de l’organisme ou de l’organe. L’important ici est donc de modéliser ces relations : on cherche à modéliser ce que fait l’organisme ou l’organe, de manière globale, mais sans chercher à comprendre quels phénomènes biologiques, au sein de cet organe, expliquent ces relations. C’est ainsi que l’on parle de « boîte noire » pour cet organisme ou cet organe, car on ne s’intéresse pas à ses mécanismes intimes.

Dans ce type de modélisation on ne s'intéresse pas aux mécanismes intimes du système effecteur.
Par exemple, dans le cas d’un centre nerveux, on cherche à modéliser quelles conséquences les informations reçues par ce centre ont sur les informations émises par ce même centre. Par contre, le détail des réseaux de neurones, des neurotransmetteurs mis en jeu au sein du centre nerveux, etc., restent « dans la boîte noire », et ne sont pas représentés.
Intuitivement, on peut ainsi comprendre que ce type de représentation permet d’étudier de manière simple (donc abordable par des non-spécialistes, tels que les élèves) mais scientifiquement exacte, les phénomènes de régulation au sein des organismes.
Homéostasie et cybernétique
La physiologie étudie les mécanismes de régulation des grandes fonctions qu’accomplissent les organismes vivants. Depuis Claude Bernard, au XIXe siècle, l’étude de la physiologie est liée à deux notions :
-
Le milieu intérieur : ensemble des liquides extracellulaires dans lesquels baignent toutes les cellules de l’organisme. Il leur permet d’être potentiellement en relation les unes avec les autres. Le support physiologique du milieu intérieur est constitué par les trois compartiments plasmatique, interstitiel et lymphatique (in « Introduction à la physiologie », B. Calvino, Belin 2003).
-
Le maintien de la constance du milieu intérieur, notion complétée ensuite par celle d’homéostasie : les paramètres du milieu intérieur sont maintenus constants dans l’organisme malgré les modifications induites par l’environnement extérieur ou l’activité de l’organisme. Cette stabilité est permise par des mécanismes régulateurs. Ainsi, les paramètres du milieu intérieur ne varient que dans d’étroites limites.
Ces valeurs fixes des paramètres du milieu intérieur constituent donc un état défini, vers lequel l’organisme tend à revenir dès qu’il s’en éloigne. Ce système est donc finalisé : on peut ainsi étudier ces mécanismes de régulation par une approche cybernétique.
La physiologie d’un organisme peut ainsi être étudiée à travers ses paramètres, stabilisés (l’homéostasie de l’organisme) par des systèmes effecteurs. Ces systèmes effecteurs (qui constituent ici nos « boîtes noires ») reçoivent des informations, correspondant aux paramètres d’entrée, ou grandeurs d’entrée (on parle aussi, cybernétiquement, d’input), et agissent sur des paramètres de sortie, ou grandeurs réglées (on parle, de même, d’output).

Ces systèmes effecteurs permettent la régulation physiologique de l’organisme. On peut aussi les qualifier de régulateurs. Schématiquement, on peut en distinguer deux types, selon les relations entre les paramètres d’entrée et de sortie. En effet, en cybernétique, un homéostat est un système asservi : les paramètres d’entrée (grandeur d’entrée) et de sortie (grandeur réglée) sont asservis, car les variations des uns sont sous le contrôle des variations des autres. Voyons maintenant ces deux types de régulateurs : en constance et en tendance.
Régulateurs en constance
En physiologie, le système effecteur correspond à un homéostat, et peut donc être assimilé à un régulateur en constance : la finalité du système effecteur est de maintenir la grandeur réglée, ou grandeur de sortie (« output »), à une valeur constante. Cette valeur constante est dénommée la grandeur de consigne, fixée au préalable, et caractéristique du régulateur en constance.
Cette régulation est réalisée en s’opposant aux variations de la grandeur d’entrée, qui ont pour effet d’éloigner la grandeur de sortie de cette grandeur de consigne. Il s’agit donc d’un effet en retour s’opposant aux variations observées : ce système fonctionne grâce à un rétrocontrôle négatif (feedback négatif).
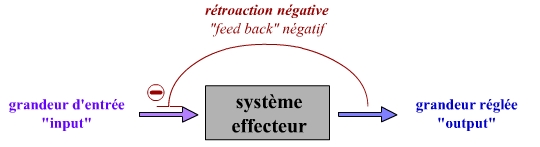
Ce régulateur est caractérisé par une rétroaction négative.
Régulateurs en tendance
Dans de rares situations physiologiques, au contraire, le système effecteur amplifie les variations de la grandeur réglée : il agit alors comme un régulateur en tendance.
La finalité d’un tel régulateur est d’entretenir une variation de la grandeur réglée, soit dans le sens d’un accroissement continue, soit dans le sens d’une diminution continue. Ce système fonctionne, au contraire d’un régulateur en constance, grâce à un rétrocontrôle positif (feedback positif), chaque variation renforçant ce qui l’a déterminée.

Ce régulateur est caractérisé par une rétroaction positive.
Il est important de noter qu’un système contrôlé par un régulateur en tendance, laissé tout seul, ne peut qu’aboutir à une impasse, soit en augmentant jusqu’à sa destruction, soit en diminuant jusqu’à son blocage complet. Ainsi, ces régulateurs sont en fait toujours couplés par un servomécanisme avec des régulateurs en constance : ceci évite que le système aboutisse à une telle impasse, dans l’organisme.
Bilan : modélisation cybernétique d’un homéostat
En cybernétique, un homéostat peut être décrit d’une manière un peu plus détaillée, par l’association de deux systèmes :
- Un système réglé : le compartiment dans lequel est définie la grandeur réglée, caractérisé en particulier par ses grandeurs d’entrée (paramètres affectant la grandeur réglée) et par sa grandeur de sortie (la grandeur réglée).
- Un système réglant : il agit en permanence sur le système réglé afin de maintenir la grandeur réglée aussi proche que possible de la valeur de consigne. Il est composé de trois éléments :
- Un émetteur, ou émetteur-capteur : détecte les variations de la grandeur réglée par rapport à la grandeur de consigne, et traduit cet écart en une information codée, transmise par le transmetteur.
- Un transmetteur : transmet l’information codée par l’émetteur au récepteur de la voie de communication.
- Un récepteur : c’est l’effecteur du système réglant. Il traduit le message codé qu’il reçoit en une action capable de rectifier la valeur de la grandeur réglée.
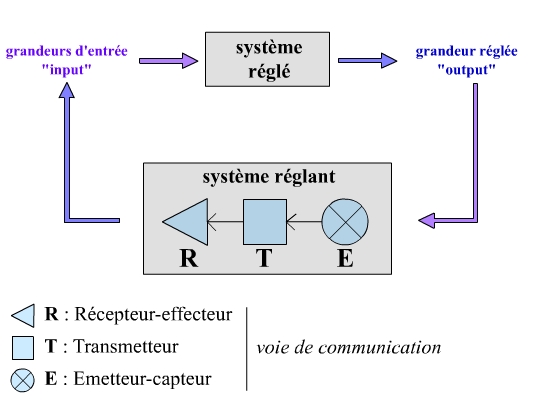
Noter l’intervention de deux systèmes, un « réglant » et un « réglé ». Grâce à une voie de communication, le système réglant peut agir sur les grandeurs d’entrée du système réglé, et ainsi positionner la grandeur réglée au niveau de la valeur de consigne.
Les servomécanismes
Dans la très grande majorité des cas, les paramètres physiologiques du milieu intérieur d’un organisme tel que l’être humain sont maintenus constants : il s’agit d’homéostats, fonctionnant grâce à des régulateurs en constance. Toutefois, dans certains cas, des régulateurs en tendance peuvent intervenir : ils résultent de la modification, de manière active, de la grandeur réglée, dans le sens d’une diminution ou d’une augmentation. Cependant, ces modifications ne sont pas infinies et sont limitées dans le temps : elles sont la conséquence du déplacement transitoire de la valeur de la grandeur de consigne vers une nouvelle valeur bien déterminée.
Dans ces phénomènes, des servomécanismes interviennent : il s’agit de dispositifs permettant de modifier la valeur de la grandeur de consigne. Ils mettent en jeu des régulateurs en tendance pour modifier la grandeur réglée, et des régulateurs en constance pour la maintenir ensuite à la nouvelle grandeur de consigne. Il s’agit donc ici d’une notion de pilotage : le servomécanisme permet une variation précise, commandé par un niveau d’organisation supérieur de l’organisme.
Premier exemple : le thermostat
Afin de comprendre plus clairement ce qu’est une représentation cybernétique, nous présentons ici un exemple simple : le thermostat. Cet exemple simple est choisi dans l’ouvrage de B. Calvino, « Introduction à la physiologie – Cybernétique et régulations », publié chez Belin, collection Belin Sup Sciences (2003).
Il s’agit d’un ensemble permettant de maintenir un système réglé à une température constante et définie, grâce à l’action d’un système réglant. Dans un premier temps, nous verrons le cas d’un bain-marie, maintenu à 37 °C par un thermostat physique. Ensuite, nous verrons comment représenter ce type de mécanismes dans le cas du corps humain, maintenu lui aussi à 37 °C.
Le thermostat du bain-marie
Le thermostat d’un bain-marie est un système asservi, dont la finalité (cybernétique) est de maintenir la température d’une enceinte constante. Ici, notre thermostat est réglé à 37 °C. On voit d’ailleurs bien ici la notion de finalité, cybernétiquement parlant : cette valeur de 37 °C est fixé à l’origine, et l’ensemble a donc un « but » : que l'eau soit à la température de 37°C.
Le thermostat d’un bain-marie se compose donc de deux systèmes : un système réglé et un système réglant.
Un système réglé : l’enceinte contenant l’eau

Sa grandeur réglée est la température de l’eau.
On peut distinguer deux grandeurs d’entrée :
- Le débit d’entrée : il s’agit de l’apport en calories fourni par la résistance du bain-marie. Ce débit d’entrée permet de chauffer l’eau et donc de faire monter sa température.
- Le débit de sortie : la cuve du bain-marie est un système ouvert. Ainsi, il y a une perte de chaleur, de manière passive (ou perte de calories). Cette perte fait baisser la température de l’eau.
Un système réglant
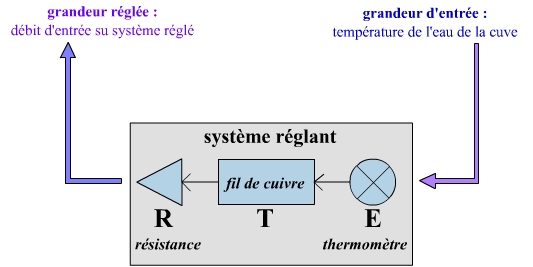
Sa grandeur d’entrée est la température de l’eau du bain-marie. Sa grandeur réglée est le débit d’entrée du système réglé : il peut ainsi agir, en régulant l’apport de calories, pour maintenir la température au niveau de la grandeur de consigne de 37 °C.
Cette action est rendue possible par une voie de communication, formée de trois éléments :
- Un émetteur-capteur : il s’agit du thermomètre, qui mesure la température de l’eau de la cuve du bain-marie, et la compare à la valeur de consigne (37 °C). Il transmet cet écart sous la forme d’une série d’impulsions électriques au transmetteur.
- Un transmetteur : il s’agit d’un fil de cuivre, qui transmet le message codé sous forme d’impulsion électriques de l’émetteur au récepteur.
- Un récepteur-effecteur : il s’agit de la résistance électrique, qui chauffe l’eau du bain-marie.
En résumé, suite à la détection d’un écart de température par rapport à la valeur de consigne (détecté par l’émetteur – le thermomètre), le récepteur (la résistance), informée via le transmetteur (le fil de cuivre), chauffe l’eau. Tant que la température atteint 37 °C, le transmetteur n’a pas d’impulsions électriques à transmettre (ou insuffisamment), et en conséquence la résistance s’arrête de chauffer.
La réunion de ces deux systèmes permet la représentation cybernétique du thermostat associé au bain-marie (Figure 8).

Le thermostat du corps humain
On peut aisément transposer cet exemple simple à un cadre physiologique : le corps humain. Dans ce cas, le système réglé correspond au milieu intérieur. De même que dans l’exemple précédent, l’organisme cherche à le maintenir à une température constante de 37 °C : la grandeur réglée de ce système est donc la température centrale de l’organisme, et la grandeur de consigne de 37 °C. Comme pour le bain-maire, ce système réglé présente deux grandeurs d’entrée : un débit d’entrée (augmentant la température) et un débit de sortie (diminuant la température).
En parallèle, un système réglant permet de maintenir la température centrale au niveau de la grandeur de consigne, en agissant sur les deux grandeurs d’entrée du système réglé (et non pas une seule comme dans le cas du bain-marie) :
- Les phénomènes de thermogenèse au niveau des muscles, et de limitations des pertes thermiques par vasoconstriction des vaisseaux sanguins, permettent d’augmenter le débit d’entrée et donc de réchauffer l’organisme.
- Les phénomènes augmentant les pertes thermiques par vasodilatation des vaisseaux sanguins, et de refroidissement par la transpiration, permettent eux de réduire le débit de sortie et ainsi de refroidir l’organisme.
On peut donc retrouver la voie de communication de ce système réglant :
- émetteur-capteur : les thermorécepteurs, cellules nerveuses sensorielles sensibles à la température. Ce sont ici les thermorécepteurs centraux (situés principalement dans l’hypothalamus) qui sont les plus importants, la grandeur réglée étant la température centrale.
- transmetteur : le système nerveux : les nerfs sensoriels transmettent l’information des capteurs vers un centre intégrateur (l’hypothalamus), et les voies nerveuses efférentes qui transmettent l’information générée par ce centre intégrateur vers les effecteurs. Le centre intégrateur est considéré comme un élément associé au transmetteur.
- récepteurs-effecteurs : il s’agit ici d’un ensemble varié d’effecteurs pouvant agir sur la température de l’organisme : les muscles squelettiques (leur travail dégage de la chaleur), les muscles lisses (qui permettent vasodilatation et vasoconstriction), les glandes sudoripares (l’évaporation de la sueur qu’elles produisent permet un refroidissement du corps).
On peut donc représenter le thermostat associé au maintien de la température centrale chez l’homme (comme chez les autres Vertébrés homéothermes) d’une manière similaire au thermostat associé au bain-marie.
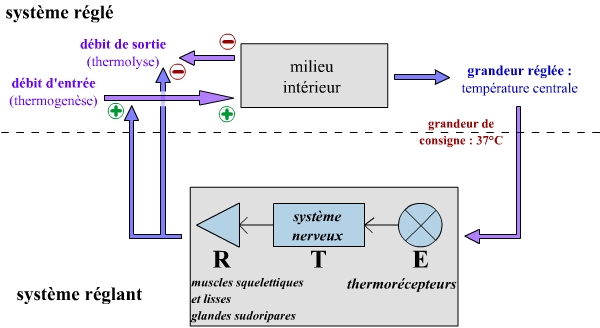
On a donc représenté ainsi cybernétiquement le système suivant :

La comparaison de ces deux schémas (cybernétique et analogique) illustre l’intérêt de la démarche cybernétique : cette représentation modélisée permet une conceptualisation « simple » d’un mécanisme physiologique par ailleurs très complexe, et dont la pleine compréhension nécessite de nombreuses connaissances en anatomie, physiologie et neurologie. En particulier, le fonctionnement précis du centre nerveux intégrateur est ici totalement occulté (grâce au concept de « boîte noire »), ce qui rend la compréhension du thermostat accessible au plus grand nombre. Les principes de fonctionnement de cet homéostat, par contre, ressortent clairement sur le schéma cybernétique.
Deuxième exemple : l’axe gonadotrope chez l’homme
L’exemple simple du thermostat permet de comprendre l’intérêt d’une représentation cybernétique. Dans le cas de phénomènes physiologiques plus complexes, cette représentation permet de conceptualiser les différents mécanismes et régulations intervenant. C’est par exemple le cas dans le contrôle de la fonction de reproduction. Chez les Mammifères, ce contrôle fait intervenir un ensemble de structures, « l’axe gonadotrope », regroupant hypothalamus, hypophyse, gonades et autres organes sexuels.
Chez l’être humain, la situation est sensiblement différente, selon que l’on étudie l’homme ou la femme. Nous traiterons ici tout d’abord du cas de l’homme, plus simple, car correspondant à une régulation en constance. Le cas de la femme, qui fait intervenir des régulateurs en tendance et en constance, reliés par des servomécanismes, est plus complexe, et sera étudié plus loin dans le document.
Rappel : l’axe hypothalamo-hypophysaire
L’hypothalamus est une structure nerveuse du cerveau, formée à partir du diencéphale au cours du développement embryonnaire (voir à ce sujet l’article Vésicules embryonnaires du système nerveux des Vertébrés). Cette structure intervient dans le contrôle de nombreux paramètres physiologiques. Cette grande importance de l’hypothalamus peut être reliée à sa structure, et en particulier à ses neurones : les neurones hypothalamiques, en effet, peuvent projeter leurs terminaisons axonales jusqu’au niveau de l’hypophyse, petite glande accrochée à la base du cerveau, juste sous l’hypothalamus. Ces neurones se projettent alors dans la partie postérieure de l’hypophyse, aussi nommée neurohypophyse. Les molécules libérées par les terminaisons axonales de ces neurones sont distribuées dans tout l’organisme par la circulation sanguine : on parle alors de neurohormones.
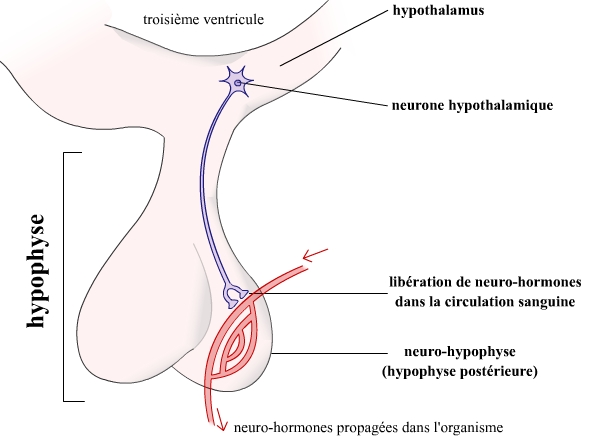
L’importance des rôles de l’hypothalamus est due à la capacité de sécrétion de neurohormones par ses neurones. Ces molécules vont pouvoir transmettre une information à distance, et ainsi réguler de nombreux mécanismes.
Toutefois, tous les neurones hypothalamiques ne se projettent pas au niveau de la neurohypophyse. Certains de ces neurones présentent des terminaisons axonales à proximité de capillaires sanguins situés non pas au niveau de l’hypophyse, mais directement dans l’hypothalamus. Ce sont ces neurones qui vont particulièrement nous intéresser ici. Les neurohormones ainsi libérées vont avoir une action à courte distance, grâce à une organisation originale du système sanguin à ce niveau : le système porte hypothalamo-hypophysaire (SPHH).
Ce système porte hypothalamo-hypophysaire est constitué de deux réseaux capillaires en série : un premier réseau de capillaires, au niveau de l’hypothalamus « reçoit » les neurohormones libérées par les neurones hypothalamiques. La veine porte achemine ces neurohormones vers le deuxième réseau de capillaires. Un deuxième réseau de capillaires permet la diffusion des neurohormones hypothalamiques vers leurs récepteurs spécifiques, portés par des cellules endocrines (sécrétrices d’hormones) situées dans l’hypophyse antérieure, ou adénohypophyse.
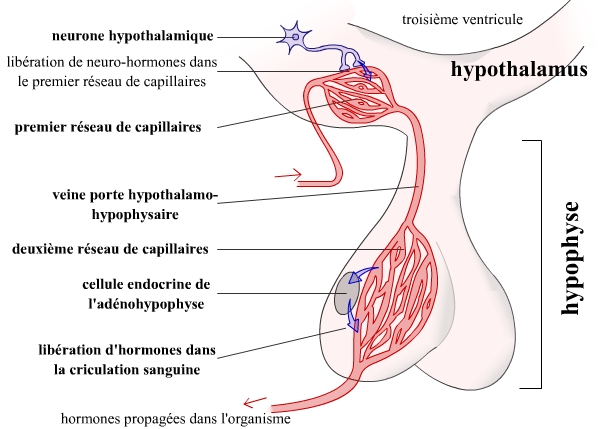
Le système porte hypothalamo-hypophysaire (SPHH) permet la transmission d’informations depuis l’hypothalamus jusqu’à l’adénohypophyse. Ces informations sont véhiculées sous forme de neurohormones, sécrétées par les neurones hypothalamiques, et captées par les cellules endocrines de l’hypophyse antérieure.
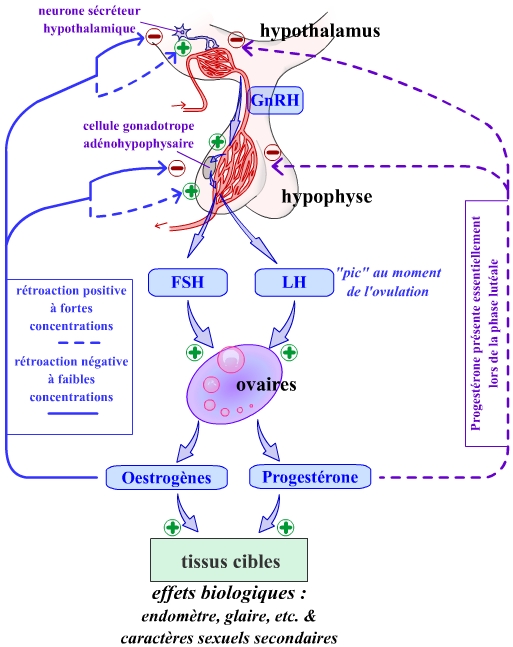
En résumé, en réponse à certains stimuli (éloignement d’une grandeur réglée de sa valeur de consigne par exemple), les neurones hypothalamiques peuvent libérer des neurohormones, véhiculées ensuite jusqu’aux cellules sécrétrices de l’adénohypophyse. Les hormones libérées par ces cellules endocrines agissent sur un effecteur, en général constitué par une glande endocrine périphérique, qui, grâce à d’autres hormones, peut alors agir sur la valeur réglée. Ces hormones périphériques exercent un rétrocontrôle sur les cellules endocrines de l’adénohypophyse et de l’hypothalamus.
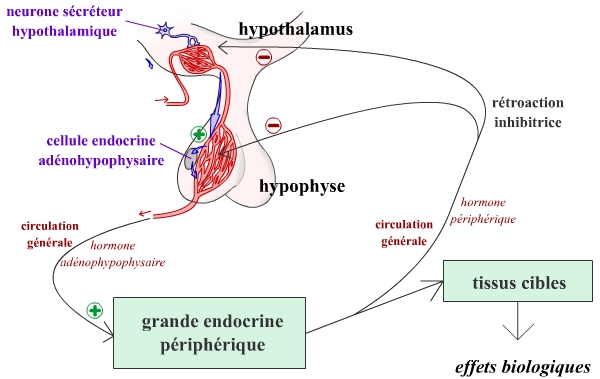
On a représenté ici le cas d’un fonctionnement en constance, grâce à une rétroaction inhibitrice. Cette rétroaction est exercée par les hormones périphériques sur les neurones sécréteurs hypothalamiques et sur les cellules endocrines adénohypophysaires.
Description fonctionnelle
L’axe gonadotrope hypothalamo-hypophysaire contrôle la fonction de reproduction chez l’homme. Cet axe s’organise de manière assez simple, très semblable au schéma donné ci-dessus.
Des neurones hypothalamiques sécréteurs libèrent une neurohormone, la gonadolibérine (GnRH – Gonadotrophin Releasing Hormone), qui est véhiculée par le système porte hypothalamo-hypophysaire jusqu’à l’adénohypophyse. À ce niveau, la GnRH se fixe sur des récepteurs spécifiques, portés par les cellules gonadotropes de l’adénohypophyse (ou hypophyse antérieure). On peut remarquer que la sécrétion de GnRH présente la particularité de ne pas être continue, mais pulsatile, par pics toutes les 60 à 90 minutes.
La fixation de la GnRH sur son récepteur stimule la sécrétion de deux hormones par les cellules gonadotropes hypophysaires : la FSH (Follicle Stimulating Hormone, hormone folliculo-stimulante) et la LH (Luteinizing Hormone, hormone lutéinisante). Ces deux hormones sont véhiculées par la circulation générale jusqu’à leurs cellules cibles, situées dans les testicules.
La LH agit sur les cellules interstitielles, ou cellules de Leydig, situées entre les tubes séminifères. Elle stimule la production de testostérone par ces cellules. Cette nouvelle hormone agit à divers niveaux, permettant la formation des spermatozoïdes, gamètes mâles, et permettant l’obtention des caractères sexuels secondaires masculins.
La FSH agit sur les cellules de Sertoli des tubes séminifères. Elle permet de faciliter la fixation de la testostérone au niveau de ces cellules. FSH et testostérone permettent ainsi de contrôler l’activité sécrétrice des cellules de Sertoli, activité qui permet la maturation des spermatozoïdes.
En retour, on observe une double rétroaction négative :
- stimulées par la LH, les cellules de Leydig sécrètent de la testostérone, qui inhibe les productions de GnRH, FSH et LH ;
- stimulées par la FSH, les cellules de Sertoli sécrètent des inhibines, qui inhibent la production de FSH.
En conclusion, l’action de l’axe gonadotrope permet de maintenir une concentration plasmatique constante de testostérone, en particulier à l’origine d’une production constante de spermatozoïdes par les testicules, gonades mâles.
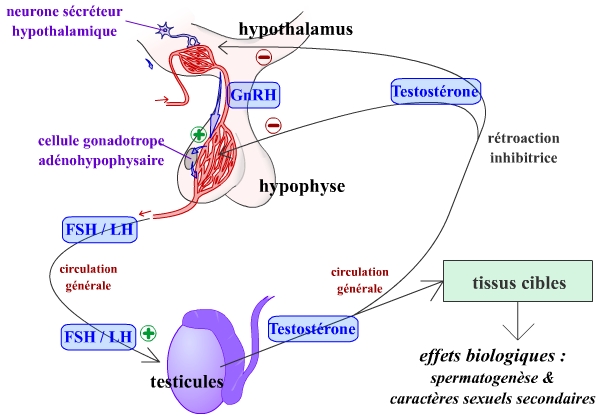
Les neurones hypothalamiques sécrètent la GnRH, qui active la libération de FSH et LH par les cellules gonadotropes de l’adénohyphyse.
FSH et LH, en agissant sur les testicules, permettent de maintenir constante la concentration plasmatique de testostérone, et ainsi les caractères sexuels masculins.
La testostérone limite en retour les sécrétions de GnRH, FSH et LH.
Schéma cybernétique
L’axe gonadotrope hypothalamo-hypophysaire peut être assimilé à un régulateur en constance de la concentration plasmatique en testostérone. On peut décomposer ce régulateur en trois systèmes, correspond aux trois organes impliqués dans l’émission de messages : hypothalamus, adénohypophyse et testicules. On peut en effet faire une représentation cybernétique de chacune des voies de communication de ces trois systèmes :

Pour chaque voie de communication, les émetteurs, transmetteurs et récepteurs sont précisés. On peut noter que les hormones sont véhiculées par le transmetteur, mais qu’elles sont en même temps dégradées par le catabolisme.
SPHH : Système porte hypothalamo-hypophysaire.
Ces trois systèmes agissent en série dans l’axe gonadotrope chez l’homme. La rétroaction négative permet un fonctionnement de l’ensemble comme un régulateur en constance.
On peut ainsi représenter l’ensemble de l’axe cybernétique sur un seul schéma, en disposant en série les schémas des trois systèmes présentés ci-dessus. On obtient alors sensiblement le même schéma que pour le fonctionnement en constance de l’axe gonadotrope chez la femme (en remplaçant œstrogènes et progestérone par la testostérone, et les ovaires par les testicules).
Ce système permet donc de maintenir une concentration plasmatique de testostérone constante dans l’organisme. Ceci permet une production constante de gamètes, tout au long de la vie de l’individu (à partir de la maturité sexuelle, bien entendu). On peut donc noter ici que la représentation cybernétique permet une modélisation simple de l’axe gonadotrope chez l’homme.
Certaines variations, que nous ne traiterons pas ici, peuvent toutefois exister. Ainsi, la concentration plasmatique de testostérone n’est pas constante au cours de la journée, par exemple. Ces variations sont dues à un changement de la valeur de consigne de l’homéostat, imposé à l’hypothalamus par d’autres structures cérébrales.
Complément : l’exemple de la régulation de la concentration plasmatique en testostérone permet de réfléchir de manière utile et efficace sur le sens du terme « rétroaction ». Voir pour cela le document Sous le terme de rétroaction, désigne-t-on toujours la même chose ? qui reprend en particulier la modélisation cybernétique de cet homéostat en termes de système réglant et système réglé.
On retrouve chez la femme de nombreux points communs, mais aussi quelques différences fondamentales d’un point de vue cybernétique.

Troisième exemple : l’axe gonadotrope chez la femme
L’axe hypothalamo-hypophysaire intervient dans le contrôle de la fonction de reproduction dans l’espèce humaine, aussi bien chez l’homme que chez la femme. Chez l’homme cet axe gonadotrope fonctionne comme un régulateur en constance de la concentration plasmatique de la testostérone. Chez la femme, au-delà de différences au niveau des hormones sexuelles impliquées, le schéma cybernétique présente des différences notables, à savoir une variabilité au cours du temps, et un fonctionnement alternativement en constance et en tendance.
Nous allons donc rappeler ici les grands principes du fonctionnement de l’axe gonadotrope chez la femme, avant d’en donner une interprétation cybernétique.
Organes et hormones impliqués dans l’axe gonadotrope chez la femme
On retrouve chez la femme un axe gonadotrope d’organisation similaire à celui de l’homme :
- Des neurones hypothalamiques secrètent une neurohormone, la GnRH (Gonadotrophin Releasing Hormone). Cette sécrétion présente, comme chez l’homme, la particularité d’être pulsatile : elle se réalise par « pics » toutes les 60 à 90 minutes en moyenne.
- La GnRH est véhiculée par le système porte hypothalamo-hypophysaire (SPHH) de l’hypothalamus à l’hypophyse antérieure, ou adénohypophyse.
- La GnRH stimule la sécrétion de deux hormones par les cellules endocrines gonadotropes de l’adénohypophyse : la FSH (Follicle Stimulating Hormone) et la LH (Luteinizing Hormone).
- FSH et LH sont véhiculées dans l’ensemble de l’organisme par la circulation sanguine.
- FSH et LH stimulent les cellules endocrines de l’ovaire. Ces cellules ovariennes secrètent deux types d’hormones : les œstrogènes (dont l’œstradiol), et la progestérone.
- Œstrogènes et progestérone, véhiculées par la circulation sanguine, agissent au niveau de divers organes cibles, permettant la réalisation de la fonction de reproduction. De plus, ces hormones agissent en rétroaction au niveau de l’hypothalamus et de l’hypophyse.
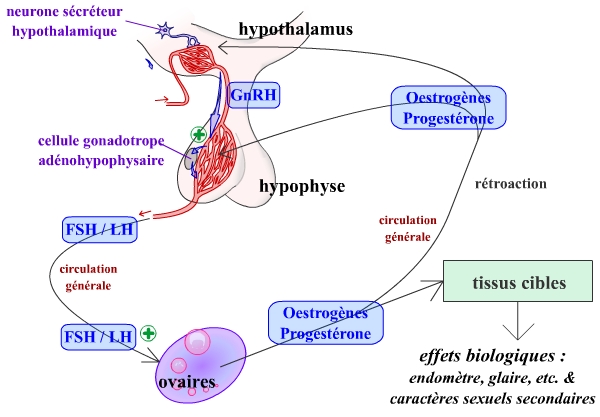
Les paramètres fluctuants ne sont pas représentés ici, ni la nature des rétroactions.
Alors que ce schéma est « stable » chez l’homme, de nombreuses variations cycliques sont observables chez la femme. Ainsi, la progestérone est par moment quasiment absente. De même, la nature de la rétroaction des hormones ovariennes varie en fonction de leur concentration. Ces variabilités expliquent un fonctionnement cyclique de l’appareil sexuel féminin.
Fonctionnement cyclique de l’axe gonadotrope
Le fonctionnement cyclique de l’appareil sexuel féminin peut être remarqué par deux évènements marquants :
- Les menstruations, desquamations de l’endomètre utérin se traduisant par des pertes sanguines.
- L’ovulation, qui correspond à l’expulsion d’un ovocyte mûr de l’ovaire. Cet ovocyte pénètre dans les trompes de Fallope, où il peut être fécondé par un spermatozoïde.
Ces deux évènements sont deux manifestations parmi de nombreuses autres du cycle sexuel féminin. Ce cycle présente une période de 28 jours en moyenne. Classiquement, on fait débuter un cycle au moment des menstruations. L’ovulation se situe environ au milieu du cycle, vers le jour 14.
On distingue ainsi deux phases :
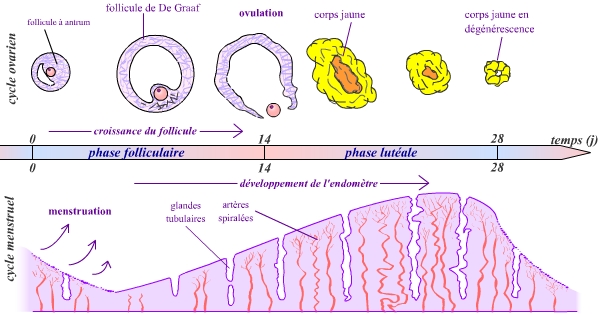
Les principaux évènements ovariens (au niveau du follicule) et menstruels (au niveau de l’endomètre) sont représentés. Un cycle très important se réalise aussi au niveau du mucus de la glaire cervicale.
Follicules et phase folliculaire : quelques précisions
Au début de la phase folliculaire, c’est en fait une cohorte d’une dizaine de follicules antraux qui amorcent leur développement. Ces follicules sont sortis du stock de follicules primaires quelques mois auparavant, et se sont développés jusqu’à ce stade en même temps que les cycles menstruels précédents se déroulaient.
Tous ces follicules antraux ne sont pas strictement identiques dans leur capacité à répondre aux gonadostimulines, et à la FSH en particulier. De ce fait, l’un deux se développe en général plus rapidement pendant la première semaine de la phase folliculaire, arrivant ainsi seul au stade de follicule pré-ovulatoire : c’est le follicule dominant.
Les autres follicules, qui se sont moins développés pendant cette première semaine de la phase folliculaire, dégénèrent pendant la deuxième semaine (c’est le phénomène d’atrésie). Ainsi, un seul follicule (en général) arrive jusqu’à l’ovulation, à la fin de cette phase folliculaire.
Au niveau de l’axe gonadotrope, le cycle féminin se caractérise par des variations cycliques des concentrations hormonales. La seule exception est la GnRH, dont les pics de sécrétion restent constant. Toutefois, la fréquence de ces pics augmente au cours de la phase folliculaire, ce qui correspond à une augmentation de la stimulation hypophysaire par la GnRH.

Courbes représentant les concentrations plasmatiques moyennes mesurables chez la femme. Ces courbes sont exactement à la même échelle temporelle que la représentation schématique du cycle sexuel proposée plus haut.
- Une phase pré-ovulatoire du jour 1 au jour 14, ou phase folliculaire. Au niveau des organes sexuels, cette phase est marquée par une croissance du follicule ovarien contenant le futur ovule (voir précisions ci-dessous), et par un développement important de l’endomètre utérin.
- Une phase post-ovulatoire du jour 14 au jour 28, ou phase lutéinique. L’endomètre continue à se développer, et peut recevoir un embryon si l’ovule a été fécondé. Le follicule ovarien (privé maintenant de son ovocyte) se transforme en corps jaune. En cas de fécondation et d’implantation de l’embryon dans l’endomètre, une grossesse commence ; le corps jaune persiste alors. Dans le cas contraire, il régresse à la fin du cycle, et l’endomètre utérin se desquame, marquant ainsi le début d’un nouveau cycle.
Les variations observées peuvent s’expliquer par divers mécanismes :
- Phase folliculaire (première moitié) : la progestérone est quasiment absente. La GnRH stimule la production de FSH et de LH, qui stimulent la production d’œstrogènes par les cellules de la thèque interne et de la granulosa du follicule ovarien. Ces œstrogènes inhibent en retour l’axe hypothalamo-hypophysaire.
Précisions : la concentration plasmatique de FSH augmente légèrement pendant cette période : elle se maintient ainsi au-dessus du seuil nécessaire au développement des jeunes follicules à antrum. En effet, les follicules à antrum ont besoin d’être stimulés par une concentration suffisante de FSH ; si la FSH se situe en dessous de cette valeur seuil, les follicules antraux ne peuvent se développer. - Phase folliculaire (deuxième moitié) : la fréquence des pics de GnRH augmente progressivement. Ceci conduit à une légère augmentation de la sécrétion de FSH et de LH, et ainsi à une hausse de la sécrétion d’œstrogènes. Quand les œstrogènes atteignent la concentration plasmatique seuil de 200 pg/mL, ils exercent une rétroaction positive sur l’axe hypothalamo-hypophysaire.
Précisions : le follicule dominant, plus en avance dans sa croissance que les autres follicules issus de la cohorte de follicules antraux de la première moitié de la phase folliculaire, secrète davantage d’œstradiol. Ceci a pour conséquence d’engendrer, par rétroaction négative, une baisse de la sécrétion de FSH.
La FSH passe ainsi sous le seuil nécessaire à la croissance folliculaire : les autres follicules subissent alors le phénomène d’atrésie. Le follicule dominant, quant à lui continue à se développer, malgré la concentration trop faible de FSH : en effet, les cellules de la granulosa ont acquis des récepteurs à LH. Le follicule dominant est donc capable de répondre à la LH, ce qui lui permet de poursuivre son développement, et de secréter ainsi de plus en plus d’œstradiol. - Ovulation : la rétroaction positive des œstrogènes (en concentration plasmatique supérieure à 200 pg/mL) induit une forte stimulation des sécrétions de GnRH, FSH et LH. Ceci conduit à un pic de LH très important, et dans une moindre mesure à un pic de FSH. Le pic de LH a pour conséquence de déclencher l’ovulation.
- Phase lutéinique : le follicule ovarien transformé en corps jaune produit de la progestérone. Cette hormone exerce une rétroaction négative sur l’axe hypothalamo-hypophysaire, réduisant ainsi les sécrétions de GnRH, FSH et LH. Dans le même temps, la synthèse d’œstrogènes par l’ovaire diminue : leur concentration plasmatique repasse sous la valeur-seuil de 200 pg/mL, et ces hormones retrouvent un rôle de rétroaction négative.
Précisions : les hormones ovariennes inhibent la sécrétion de FSH et de LH. La concentration de FSH est ainsi maintenue sous le seuil nécessaire à la croissance des follicules. La concentration de LH, par contre, reste à un niveau suffisant pour assurer la stimulation de la production des hormones ovariennes (notamment de progestérone par le corps jaune). - Fin de la phase lutéinique : l’arrêt de l’activité du corps jaune entraîne une chute des concentrations plasmatiques des hormones ovariennes (notamment de progestérone), et donc une forte diminution de l’inhibition qu’elles exercent sur l’axe hypothalamo-hypophysaire. Les concentrations de FSH et de LH peuvent ainsi recommencer à augmenter : un nouveau cycle débute.
En cas de fécondation, l’activité du corps jaune se maintient, évitant ainsi le démarrage d’un nouveau cycle.
Comme on le voit ici, la nature même des rétroactions qui ont lieu est primordiale dans le déroulement de ce cycle. Une représentation cybernétique, qui permet de mettre l’accent sur l’importance de ces rétroactions est donc particulièrement adaptée.
Modélisation cybernétique : principes chez la femme
Il est possible de reprendre la représentation cybernétique de l’axe gonadotrope chez l’homme, pour l’adapter à ce qui se passe chez la femme. D’un point de vue cybernétique, en effet, ce qui change fondamentalement chez la femme, c’est :
- que l’on remplace la testostérone par les œstrogènes (et que l’on rajoute la progestérone en phase lutéinique) ;
- qu’en fin de phase folliculaire, juste avant l’ovulation, lorsque la concentration en œstrogènes plasmatiques augmente, au-delà d’un certain seuil (la valeur exacte n’importe pas pour une représentation cybernétique), le sens de la régulation exercée par les œstrogènes s’inverse.
Ce seuil atteint, un servomécanisme (changement du pilotage, de la commande de la régulation) se met en place (par un mécanisme que l’on ignore) au niveau de l’hypothalamus. Il permet l’inversion du sens de la régulation : la régulation négative exercée par les hormones œstrogéniques (régulation en constance) devient positive (régulation en tendance) : alors que les œstrogènes inhibent la libération de GnRH le reste du temps, ils la stimulent maintenant.
Cette augmentation des œstrogènes est elle-même initiée à l’origine par une augmentation de l’activité sécrétrice pulsatile des neurones hypothalamiques par un mécanisme encore mal compris. On suppose que ce mécanisme pourrait être relié à l’horloge biologique interne. On peut noter à ce propos que ceci dégage l’importance du contrôle de cet axe gonadotrope par des niveaux cérébraux supérieurs.
Par voie de conséquence, la GnRH (qui stimule la sécrétion de FSH et LH par les cellules gonadotropes de l’adénohypophyse) induit le pic de LH et FSH, à l’origine de l’ovulation.
Après l’ovulation, la concentration plasmatique en œstrogènes s’effondre transitoirement et le sens de la régulation s’inverse : de positive, la rétroaction exercée par les hormones plasmatiques redevient négative. Le servomécanisme rétablit une grandeur de consigne fixe. La concentration en œstrogènes remonte un peu pendant la phase lutéinique, mais sans atteindre la valeur-seuil de déclenchement du servomécanisme : on revient en situation basale de régulation en constance négative exercée par les œstrogènes (et la progestérone, qui est désormais aussi présente) sur les neurones hypothalamiques compétents et sur les cellules gonadotropes de l’adénohypophyse.
Modélisation cybernétique : schémas
En somme, on peut représenter l’axe gonadotrope chez la femme par trois schémas cybernétiques :
- un schéma de régulateur en constance pendant l’essentiel de la phase folliculaire ;
- un schéma de régulateur en tendance à la fin de la phase folliculaire, juste avant l’ovulation ;
- un schéma de régulateur en constance pendant la phase lutéinique.
Or, la seule différence entre le premier et le troisième schéma se situe au niveau des hormones ovariennes impliquées : œstrogènes seuls en phase folliculaire, œstrogènes et progestérone en phase lutéinique. D’un point de vue cybernétique, œstrogènes et progestérones remplissent les mêmes rôles, pendant la phase lutéinique : rétroaction négative sur l’axe hypothalamo-hypophysaire et actions variées sur les organes sexuels.
Il est donc possible de simplifier cette représentation par deux schémas, correspondants aux deux situations cybernétiques :
- un schéma de régulateur en constance pendant l’essentiel du cycle ;
- un schéma de régulateur en tendance juste avant l’ovulation.
Chacun de ces schémas est établi à partir des schémas des trois systèmes impliqués (hypothalamus émetteur, adénohypophyse émetteur, et ovaires émetteurs – voir dans l’exemple de l’axe gonadotrope de l’homme une représentation similaire de ces trois systèmes).
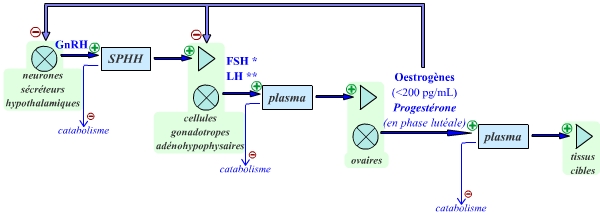
Les périodes de fonctionnement en constance de l'axe gonadotrope chez la femme sont la phase folliculaire, à l’exclusion de la période pré-ovulatoire, et la phase lutéinique.
FSH : la FSH ne stimule la croissance folliculaire qu’au-dessus de sa valeur seuil, pendant la première moitié de la phase folliculaire.
LH : La LH n’agit que sur le follicule dominant et le corps jaune.
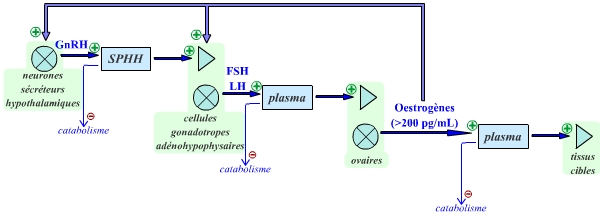
La période de fonctionnement en tendance de l'axe gonadotrope chez la femme correspond à la phase pré-ovulatoire du cycle menstruel.
L’existence d’un phénomène cyclique s’explique donc par l’existence d’un servomécanisme, inactif pendant la majeure partie du cycle, mais qui permet un fonctionnement en tendance positive (d’où la très forte augmentation de FSH et surtout de LH) à un moment donné du cycle.
La représentation cybernétique pendant le fonctionnement en constance présentée ici fait figurer sur un même schéma des hormones agissant à différents moments du cycle, et sur différentes cibles. Il s’agit donc d’une représentation simplifiée. Pour plus de détails sur les seuils de FSH et d’œstrogènes, ainsi que sur les cibles de la LH, revoir les explications données sur le fonctionnement cyclique de l’axe gonadotrope ci-dessus. On peut d’ailleurs noter que si l’accent est souvent mis sur l’importance du rétrocontrôle positif permettant le déclenchement de l’ovulation, le rétrocontrôle négatif, notamment vis-à-vis de FSH, est lui aussi fondamental d’un point de vue fonctionnel ; en particulier en intervenant dans les mécanismes permettant l’arrivée à maturité d’un seul follicule issu de la cohorte de follicules antraux.
Différences cybernétiques entre l’homme et la femme
D’un point de vue cybernétique, il est donc intéressant de constater que les différences entre l’homme et la femme ne sont finalement que minimes :
- Dans les deux cas, l’axe gonadotrope fonctionne essentiellement en constance, l’hormone périphérique étant la testostérone chez l’homme, les œstrogènes et par moment la progestérone chez la femme.
- Alors que le fonctionnement chez l’homme reste en constance, un servomécanisme permet un fonctionnement en tendance transitoire chez la femme, qui permet l’ovulation.
L’existence de ce servomécanisme est donc suffisant pour permettre de passer d’une production continue de gamètes, chez l’homme, à une production périodique, chez la femme.
Conclusion
La cybernétique permet de modéliser les processus physiologiques. L’intérêt principal de ce type d’approche est d’intégrer la finalité du système, en particulier par les notions de grandeur de consigne et de servomécanisme.
Définitions et conventions
Pour finir, nous proposons ici quelques définitions générales de termes utilisés couramment en cybernétique et en physiologie (d’après B. Calvino, « Introduction à la Physiologie » (224 pages), publié chez Belin (Paris) dans la collection Belin Sup Sciences (2003) – d’autres exemples et des précisions sont présentes dans cet ouvrage).
- Cybernétique : science du pilotage et du gouvernement. Elle traite des processus de communication et de régulation dans les êtres vivants comme dans les machines.
- Émetteur-capteur : premier élément d’une voie de communication. Il transforme les variations d’un paramètre physique ou physiologique en un message codé qu’il émet dans le transmetteur de la voie de communication.
- Grandeur de consigne : pour un homéostat, grandeur fixée autour de laquelle varie en permanence la grandeur réglée.
- Grandeur d’entrée : facteur agissant sur le système réglé d’un régulateur, et intervenant dans les variations de la grandeur de sortie.
- Grandeur réglée : dans le système réglé d’un homéostat, paramètre défini et maintenu à une valeur aussi constante que possible par l’action du système réglant. Il constitue la grandeur de sortie d’un régulateur.
- Homéostasie : état stable d’un système maintenu semblable à lui-même, et caractérisé par la stabilité des paramètres le définissant. L’homéostasie caractérise donc la stabilisation de paramètres d’un système autour de valeurs fixes, hors d’un équilibre thermodynamique.
- Homéostat : système régulé et asservi construit dans le but de maintenir la grandeur réglée le caractérisant aussi constante que possible, à une valeur oscillant autour de la grandeur de consigne qui lui a été préalablement fixée. On peut le modéliser par deux systèmes : un système réglé, dont la grandeur de sortie est maintenue constante par l’action d’un système réglant sur sa (ses) grandeur(s) d’entrée.
- Récepteur – effecteur : élément terminal d’une voie de communication qui reçoit du transmetteur le message codé émis par l’émetteur, puis le décode. Le récepteur est aussi un effecteur : il agit sur les grandeurs d’entrée du système réglé, dans le but de maintenir la grandeur réglée à une valeur fixée.
- Régulateur : système muni d’un dispositif de contrôle construit dans le but d’intervenir pour modifier des facteurs (ou grandeurs d’entrée) à l’origine de variations de la grandeur réglée.
- Rétroaction ou feedback : action en retour déclenchée par les variations de la grandeur réglée d’un régulateur, qui modifie les facteurs à l’origine de ces variations. Les rétroactions négatives correspondent à un fonctionnement en constance ; les positives à un fonctionnement en tendance.
- Servomécanisme : système asservi, sous le contrôle d’une information permettant de piloter au cours du temps des variations de la grandeur de consigne d’un homéostat.
- Système asservi : système maintenu sous le contrôle d’un asservissement qui pilote son fonctionnement. Il peut s’agir d’un régulateur en constance, d’un régulateur en tendance, ou d’un servomécanisme.
- Transmetteur : élément intermédiaire d’une voie de communication qui transmet au récepteur le message codé émis par l’émetteur.
- Voie de communication : ensemble constitué d’un émetteur-capteur, d’un transmetteur et d’un récepteur-effecteur. Il assure la communication entre deux ensembles en permettant le transfert d’un message codé.
Crédits