Cet article présente une comparaison d'Homo sapiens et d'Homo neanderthalensis, notamment au niveau de l'ADN mitochondrial (séquences téléchargeables), et discute des relations de parenté possibles entre ces deux types d'individus.
Homo neanderthalensis et Homo sapiens
Si de nos jours seule l’espèce Homo sapiens est présente sur Terre, les études paléoanthropologiques montrent que plusieurs espèces du genre Homo ont existé par le passé, et sur des périodes de temps qui parfois se recouvrent. Ainsi, l’homme de Néandertal (Homo neanderthalensis) a vécu de – 200 000 ans B.P. à – 30 000 ans B.P. environ (B.P. : « before present », âge par rapport à nos jours) ; dans le même temps, l’homme moderne (Homo sapiens) est apparu il y a probablement environ 130 000 ans (les premiers Homo sapiens sont couramment nommés « hommes de Cro-Magnon »). De ce fait, ces deux types d’homme ont coexisté pendant près de 100 000 ans.
De nombreuses questions se posent donc quant aux relations précises entre l’homme de Néandertal et l’homme de Cro-Magnon (voir la bibliographie pour une hypothèse sur la disparition de H. neanderthalensis).
La première de ces questions consiste à se demander s’ils étaient interféconds quand ils vivaient dans les mêmes lieux. Dans l’affirmative, ils ne pourraient être classés comme deux espèces différentes et devraient être appelés respectivement Homo sapiens neanderthalensis et Homo sapiens sapiens. Dans le cas contraire, formant deux espèces distinctes, ils mériteraient bien les deux patronymes d’Homo neanderthalensis et d’Homo sapiens.
Une deuxième question, directement reliée à la première, est en rapport avec l’origine de l’homme moderne : est-il apparu d’abord en Afrique, d’où il aurait ensuite migré vers le reste de la planète ? Ou bien l’homme moderne est-il le fruit de croisements, éventuellement multiples, entre des hommes de Néandertal et des Homo sapiens ancestraux ? Ces questions reviennent en fait à nous demander si nous, hommes modernes, portons dans nos gènes un héritage de l’homme de Néandertal.
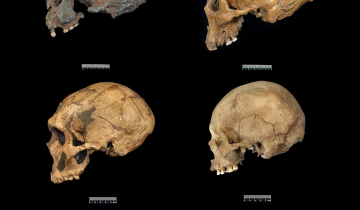
Pendant longtemps, le seul moyen de répondre à ces questions a été d’effectuer des comparaisons anatomo-morphologiques entre Homo neanderthalensis et Homo sapiens, sur la base des fossiles disponibles (voir tableau et figure ci-dessous).
| Homo neanderthalensis | Homo sapiens | ||
|---|---|---|---|
| Habitat | Tempéré ou froid entre 35°N et 50°N | Tous les habitats | |
| Stature | Trapue hommes : 1,65 m pou 90 kg femmes : 1,55 m pour 70 kg |
Longiligne et robuste (H.s. ancien) hommes : 1,70 m pour 70 kg femmes : 1,65 m pour 55 kg |
Longiligne et gracile (H.s. sapiens) hommes : 1,63 pour 63 kg femmes : 1,56 pour 50 kg |
| Boîte crânienne | Volumineuse, en bombe 1500 à 1750 cm3 |
Volumineuse, sphérique chez H.s. sapiens 1650 (H.s. ancien) à 1350 cm3 (H.s. sapiens) |
|
| Face | Nez très grand et saillant, pommettes et menton absents | Nez étroit, présence de pommettes saillantes et d’un menton | |
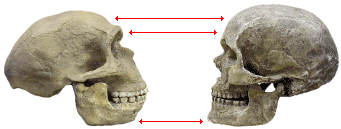
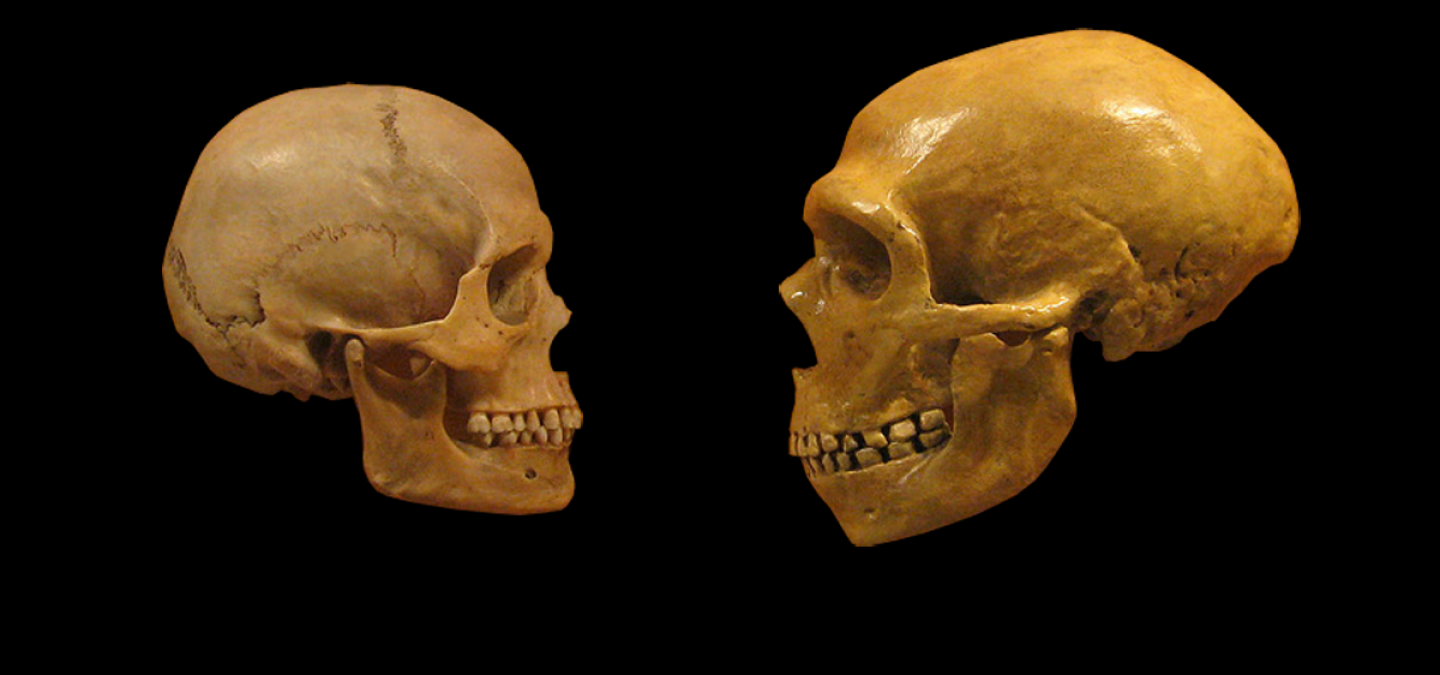
Le crâne d’H. sapiens est celui de droite.
Les flèches soulignent trois points de comparaison possibles entre ces deux crânes : le front, l’arcade sourcilière et le menton.
Noter aussi la présence du « chignon occipital », excroissance du crâne au niveau le plus postérieur, chez H. neanderthalensis.
Image reproduite avec l’aimable autorisation de Dennis O'Neil – « Evolution of modern humans tutorial »
Utilisation de l’ADN mitochondrial
Au-delà des études purement morphologiques, la biologie moléculaire est un autre moyen de déchiffrer les rapports entre H. neanderthalensis et H. sapiens.
En effet, de l’ADN a pu être extrait de fossiles d’hommes de Néandertal et d’Homo sapiens ancestraux. Il s’agit d’un ADN particulier : celui des mitochondries, organites qui ont l’avantage d’être présents en de multiples exemplaires dans une même cellule et de contenir chacun de nombreuses molécules d’ADN similaires (alors qu’on ne trouve qu’un seul noyau par cellule, ne contenant qu’un unique exemplaire de chaque ADN paternel et maternel).
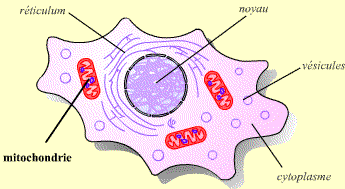
Les mitochondries sont présentes en grand nombre dans le cytoplasme (la représentation donnée ici n’est pas à l’échelle, les mitochondries étant en réalité bien plus petites).
Une des plus grandes difficultés de cette manipulation est d’éviter, par l’application de règles absolument draconiennes, la contamination de l’ADN fossile par de l’ADN actuel, contamination qui ruinerait toute comparaison…
Outre l’avantage du nombre, l’ADN mitochondrial (ADNmt), qui est d’héritage presque uniquement maternel, a l’intérêt d’être garanti contre les recombinaisons entre chromosomes paternels et chromosomes maternels existant au sein de l’ADN nucléaire.
L’ADN mitochondrial permet ainsi de pister l’histoire évolutive sans le brouillage des recombinaisons. Avec la limite, toutefois, que cette histoire évolutive reste cantonnée à une histoire essentiellement maternelle.
Lors de la fécondation d’un ovule, les mitochondries du spermatozoïde ne sont pas retrouvées dans l’œuf fécondé. Ainsi, les mitochondries du futur organisme sont d’origine maternelle. Or, les mitochondries possèdent un ADN qui leur est propre, présent en de nombreux exemplaires au sein de leur matrice. Cette information génétique mitochondriale n’est donc pas transmise de manière mendélienne, mais par les mères.
Une fois l’ADN mitochondrial extrait des fossiles (ou directement d’hommes modernes), il faut choisir une séquence nucléotidique particulière pour comparer ces ADN. Pour cela, après amplification par réaction en chaîne de la polymérase (ou PCR), on séquence de courtes régions hypervariables connues pour leur fréquence de mutation élevée. Cela permet de suivre une histoire évolutive concentrée sur une période de temps relativement courte (quelques centaines de milliers d’années seulement).
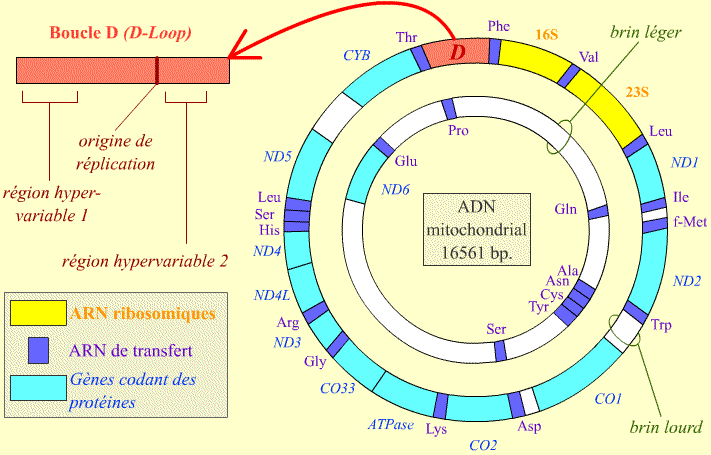
Le génome d'une mitochondrie code pour des ARN de transfert, les ARN ribosomiques et quelques protéines comme le cytochrome b (CYB) et l’ATP synthase (ATPase). Les deux brins complémentaires de l’ADN mitochondrial sont représentés par convention, et en référence à une légère différence de poids moléculaire, comme « brin léger » et « brin lourd ». La boucle D (« D-Loop ») comporte l’origine de réplication, divers éléments de régulation de l’expression des gènes mitochondriaux, et des régions hypervariables.
Séquence à télécharger :
- hs-mitochondrie-cplete.adn : ADN mitochondrial humain complet ; fichier .adn utilisable directement sous Anagène.
- hs-mitochondrie-cplete.txt : ADN mitochondrial humain complet ; fichier texte comportant les annotations de la séquence (référence GenBank : HUMMTCG).
Pour importer les fichiers, enregistrer la cible des liens.
ADN mitochondriaux d’Homo sapiens et Homo neanderthalensis : résultats
Que révèle alors la biologie moléculaire sur l’histoire des relations entre Néandertal et sapiens ?
David Caramelli de l’université de Florence et ses collaborateurs ont comparé en mars 2003 ces séquences hypervariables de deux hommes de Cro-Magnon de la grotte de Paglicci en Italie du sud datés de – 24 720 et – 23 000 ans B.P., de quatre hommes de Néandertal proches géographiquement et datés de – 42 000 à – 29 000 ans B.P., de quatre hommes préhistoriques datés de -14 000 à -5 500 ans B.P., et enfin de celles figurant dans une banque de données d’ADN mitochondrial de 2 566 hommes actuels d’Europe et du Moyen-Orient.
Les séquences des hommes de Cro-Magnon sont très semblables à celles des hommes préhistoriques plus récents et à celles des hommes actuels. Elles se distinguent en revanche nettement de celles des néandertaliens, voisins pourtant par la géographie et, pour certains, par l’âge.
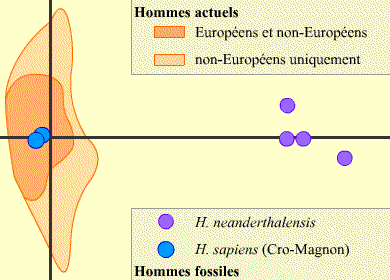
Le graphique correspond à une représentation (Multidimensional scaling, ou MDS) dans laquelle la distance entre deux points est proportionnelle à la distance génétique entre les deux séquences correspondantes.
Sont représentés les résultats de la comparaison de la région hypervariable 1 de la boucle D de l’ADN mitochondrial de six hommes fossiles (quatre H. neanderthalensis et deux H. sapiens ancestraux), sous forme de cercles, et de nombreux Homo sapiens actuels.
Pour des raisons de simplification, les points des hommes actuels ne sont pas indiqués, mais leurs emplacements sont matérialisés par une aire orangée.
On observe que les H. sapiens sont très proches les uns des autres, qu’ils soient fossiles ou pas, Européens ou non. Au contraire, les H. neanderthalensis sont proches entre eux, mais éloignés des H. sapiens ancestraux ou actuels.
Ces résultats sont un argument supplémentaire, en plus des comparaisons morphologiques fines de Katerina Harvati (université de New-York), pour estimer que les hommes de Néandertal ne sont pas une sous-espèce d’Homo sapiens mais bien une espèce à part entière, Homo neanderthalensis. On peut ainsi supposer qu’il n’y a pas eu de mélanges génétiques entre ces deux espèces humaines, malgré parfois leur proximité géographique.
De plus, les études réalisées de manière plus générale sur le génome mitochondrial de l’homme actuel suggèrent qu’il est âgé (sous sa forme actuelle) d’environ 163 000 ans. Or la séparation initiale entre les lignées ayant donné H. sapiens et H. neanderthalensis est estimée à environ 500 000 ans B.P. (de 365 à 853 000 ans B.P. selon les estimations les plus larges) d’après des calculs d’horloge moléculaire. Cela confirme l’idée d’un génome mitochondrial ayant évolué de manière séparée chez ces deux espèces, et donc l’absence de mélanges génétiques entre sapiens et Néandertal.
ADN mitochondriaux d’Homo sapiens et Homo neanderthalensis : conclusions
Les diverses études menées sur les régions hypervariables de la boucle D de l’ADN mitochondrial convergent toutes vers la même conclusion : il n’y a probablement pas eu de mélanges génétiques entre Homo sapiens et Homo neanderthalensis. L’hypothèse selon laquelle ces individus correspondent bien à deux espèces humaines distinctes est ainsi confortée. De plus, de tels résultats confortent la théorie dite « Out of Africa », selon laquelle l’homme moderne serait apparu en Afrique, puis aurait migré vers le reste de la planète (au contraire des autres modèles, proposant une apparition multiple et locale de l’homme moderne).

Cette figure est une synthèse des conclusions de Krings et al. (1997, 1999 et 2000) et Ovchinnikov (2000), citées dans « Aux origines de l’humanité – La génétique au service de la quête de nos origines », p 503).
Les séquences de chimpanzés sont utilisées comme groupe extérieur pour enraciner l’arbre.
Toutefois la prudence s’impose, car ces séquences hypervariables, longues d’à peine quelques centaines de nucléotides, ne peuvent raconter la même histoire que celle de l’ADN du noyau riche de ses trois milliards de nucléotides. Les mitochondries ne se transmettant que par les mères, ces études sur l’ADN mitochondrial suggèrent qu’il n’y aurait pas de femmes de Néandertal parmi nos ancêtres. À cette restriction près, apportée par de récentes études, qu’il existe de rares transmissions paternelles des mitochondries.
Une autre limite importante de ces résultats est le très faible nombre d’échantillons fossiles utilisés. Seuls quelques hommes de Néandertal et quelques hommes de Cro-Magnon ont en effet été analysés. Et il n’est pas impossible que ce faible échantillonnage ait entraîné un biais, non détecté encore…
De plus, il est à noter que les ADN extraits de ces fossiles peuvent très bien avoir subi des mutations au cours des milliers d’années qui se sont écoulées. Toutefois, la très forte homologie entre les hommes de Cro-Magnon étudiés et les hommes actuels, de même que la similarité des résultats entre des hommes de Néandertal issus de différents sites, suggèrent que ce phénomène reste négligeable, au moins pour ces durées de conservation et ces séquences.
Ainsi, même si le modèle le plus probable aujourd’hui paraît être celui d’une colonisation de la planète par des Homo sapiens issus d’Afrique, remplaçant les Homo neanderthalensis sans mélanges génétiques, il n’est pas possible à l’heure actuelle d’apporter une réponse définitive à cette question.
L’idéal serait évidemment de disposer un jour d’un ADN nucléaire, au lieu des seuls ADN mitochondriaux. Cette éventualité n’est peut-être pas hors de portée puisque, récemment, un tel ADN a pu être extrait d’un ours des cavernes de Croatie, vieux de 33 000 ans.
ADN mitochondriaux d’Homo sapiens et Homo neanderthalensis : séquences utilisables
Téléchargement
Le thème d’étude suivant est ouvrable sous Anagène (version 1 ou supérieure) :
Pour visualiser ces séquences, on peut utiliser le logiciel Anagène. Pour importer le fichier, enregistrer la cible du lien.
Explications du fichier
Le fichier D-loop.edi, une fois ouvert sous Anagène, présente 9 séquences d’ADN mitochondrial. Ces séquences proviennent d’Homo neanderthalensis et d’Homo sapiens fossiles, ainsi que d’Homo sapiens modernes. Elles correspondent soit à la région hypervariable 1, soit à la région hypervariable 2 de la boucle D de l’ADN mitochondrial. La datation approximative des spécimens d’où ont été extraits les ADN mitochondriaux est indiquée.
Liste des 9 séquences d’ADN mitochondrial disponibles dans le fichier D-loop.edi
| Séquence | Espèce | Région de la boucle D | Datation du specimen | Remarques |
| hn-D1-1997.adn | H. neanderthalensis | hypervariable 1 | 29 000 ans | spécimen de vallée de Néander en Allemagne, trouvé en 1856 (Ref. 1) |
| hn-D1-2000.adn | H. neanderthalensis | hypervariable 1 | 28-29 000 ans | spécimen de la grotte de Vindija en Croatie (Ref. 2) |
| hs-D1-2003-12.adn | H. sapiens | hypervariable 1 | 24-25 000 ans | Cro-Magnon de la grotte de Paglicci en Italie (Ref. 3) |
| hs-D1-2003-25.adn | H. sapiens | hypervariable 1 | 23 000 ans | Cro-Magnon de la grotte de Paglicci en Italie (Ref. 3) |
| hs-D1-sujetAS.adn | H. sapiens | hypervariable 1 | actuel | prélèvement sur un individu Européen (Ref. 4) |
| hs-mitochondrie-D1.adn | H. sapiens | hypervariable 1 | actuel | extrait de la séquence complète du génome mitochondrial (Ref. 5) |
| hn-D2-1999.adn | H. neanderthalensis | hypervariable 2 | 29 000 ans | spécimen de vallée de Néander en Allemagne, trouvé en 1856 (Ref. 6) |
| hn-D2-2000.adn | H. neanderthalensis | hypervariable 2 | 28-29 000 ans | spécimen de la grotte de Vindija en Croatie (Ref. 2) |
| hs-mitochondrie-D2.adn | H. sapiens | hypervariable 2 | actuel | extrait de la séquence complète du génome mitochondrial (Ref. 5) |
Il est à noter que les séquences hn-D1-1997.adn et hn-D2-1999.adn proviennent du même individu. Il s’agit de l’individu type de l’espèce Homo neanderthalensis. De même, les séquences hn-D1-2000.adn et hn-D2-2000.adn proviennent du même individu.
Utilisations possibles
Ces séquences peuvent être utilisées de différentes façons. Sans rentrer dans les détails, voici quelques grandes pistes d’utilisation :
- Alignement des 6 séquences de la région hypervariable 1 (D1) : mise en évidence de l’homologie entre H. neanderthalensis et H. sapiens.
- Alignement des 2 séquences D1 d’H. neanderthalensis, puis des 4 séquences D1 d’H. sapiens : mise en évidence de la très forte homologie au sein de chaque espèce. À noter la très forte homologie entre H. sapiens actuel et H. sapiens fossile (spécimens type Cro-Magnon).
- Alignement d’une séquence D1 d’H. neanderthalensis avec une séquence D1 d’H. sapiens : mise en évidence d’un plus grand nombre de différence entre les espèces qu’au sein de chaque espèce.
- Alignement des 3 séquences de la région hypervariable 2 (D2) : mise en évidence d’une forte homologie au sein des H. neanderthalensis, la séquence d’un homme actuel étant divergente.
Références
- GenBank : AF011222. Article : « Neandertal DNA sequences and the origin of modern humans », Krings et al., Cell 90 (1997), 19-30.
- GenBank : AF282971 et AF282972. Article : « A view of Neandertal genetic diversity », Krings et al., Nat. Genet. 26 (2000), 144-146
- GenBank : AY283028 et AY283027. Article : « Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans », Carameli et al., Proc. Natl. Acad. Sci. U.S.A. 100 (2003), 6593-6597. Article disponible en texte intégral gratuit.
- GenBank : MTHSAS. Séquence publiée en 1999.
- GenBank : HUMMTCG. Compilation de diverses séquences publiées de 1979 à 1991.
- GenBank : AF142095. Article : « DNA sequence of the mitochondrial hypervariable region II from the neandertal type specimen », Krings et al., Proc. Natl. Acad. Sci. U.S.A. 96 (1999), 5581-5585. Article disponible en texte intégral gratuit.
Complément : hypothèse sur la disparition d’H. neanderthalensis
Deux chercheurs des universités des Cambridge et Southampton viennent de publier un livre retraçant leurs conclusions sur les causes de la disparition de l’homme de Néandertal.
Pourquoi Néandertal n’a-t-il pas connu la même fortune évolutive que nous autres sapiens ? Quelles peuvent être les causes de son extinction, il y a environ 30 000 ans : une moindre capacité à faire face aux rigueurs d’un climat devenu glacial ? Une moins bonne compétitivité vis-à-vis d’Homo sapiens ?
Pour répondre à cette question Tjeerd van Andel de l’université de Cambridge a mené une formidable enquête mobilisant 31 collaborateurs sur pas moins de 400 sites à travers l’Europe pour mieux connaître ces hommes de Néandertal et premiers sapiens ayant vécu entre – 40 000 et – 20 000 ans : leur âge déterminé par datation absolue, leurs outils, le milieu dans lequel ils vivaient aussi bien pour sa flore que pour sa faune, les climats qu’ils rencontraient…
Les cartes produites sont si précises qu’elles permettent de suivre les migrations des hommes de Néandertal et des premiers sapiens dans leur milieu de vie avec leurs équipements et leurs outils.
Tous connaissaient le feu, et les sapiens du groupe des aurignaciens se nourrissaient des mêmes proies que les hommes de Néandertal les découpant avec un égal talent. Mais ils ne disposaient pas de tout le savoir-faire technique d’un autre sapiens, du groupe des gravettiens venus de l’est : des javelots permettant une chasse à bonne distance de l’animal au lieu de celle au corps à corps seule autorisée par les lances des hommes de Néandertal et des aurignaciens, des filets de pêche, des manteaux de fourrure cousus, des habitations encloses.
Alors Néandertaliens et sapiens aurignaciens, dépassés techniquement, auraient tous deux fini par succomber tandis que les sapiens gravettiens prospéraient et prenaient la relève.
Quelques liens pour approfondir ces notions
- L’ENS Lyon propose une excellente page sur l’utilisation des ADN anciens.
- Une discussion rapide sur les relations sapiens – neanderthalensis sur Planet-Terre [site ressources ENS-DESCO sciences de la Terre]
- Un ensemble de cours de paléoanthropologie, extrêmement bien fait, par le Prof. Dennis O'Neil de l’Université de Palomar, Californie, USA (en anglais).
- Deux articles majeurs sur les analyses d’ADN mitochondrial néandertalien sont disponibles gratuitement en texte intégral sur Internet (en anglais) :
- « Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans », Carameli et al., Proc. Natl. Acad. Sci. U.S.A. 100 (2003), 6593-6597.
- « DNA sequence of the mitochondrial hypervariable region II from the neandertal type specimen », Krings et al., Proc. Natl. Acad. Sci. U.S.A. 96 (1999), 5581-5585
- Enfin, plusieurs chapitres passionnants sur cette thématique sont présents dans l’ouvrage collectif « Aux origines de l’humanité – Tome 1 : de l’apparition de la vie à l’homme moderne », sous la direction de Yves Coppens et Pascal Picq, Fayard (2002).