

L'ATP est la molécule clef de l'énergétique cellulaire mais elle joue aussi d'autres rôles au sein de la cellule.
Cet article présente la structure et les trois grand types de fonction de cette molécule,
La molécule
L’ATP est un nucléotide, c’est-à-dire une molécule formée par un sucre à 5 atomes de carbone, le désoxyribose, une base azotée, l’adénine, et une chaîne de trois acides phosphoriques (voir fig. 1).
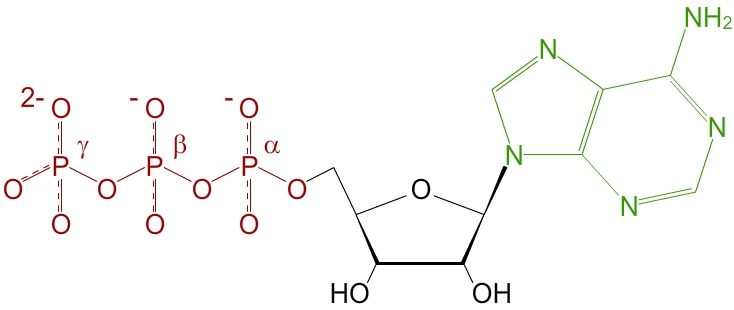
On distingue bien les trois parties de la molécule d'ATP : le sucre à 5 carbones en noir (du ribose), la base azotée en vert (de l'adénine) et les trois groupements phosphates, chargés en conditions physiologiques, en rouge.
L’ATP entre dans la fabrication des acides nucléiques
Si la molécule d’ATP est trouvée à l’état libre dans les cellules, elle sert également de matériau de construction pour la synthèse des acides nucléiques, la classe de macromolécules essentiellement en charge de l’information génétique (voir fig. 2).
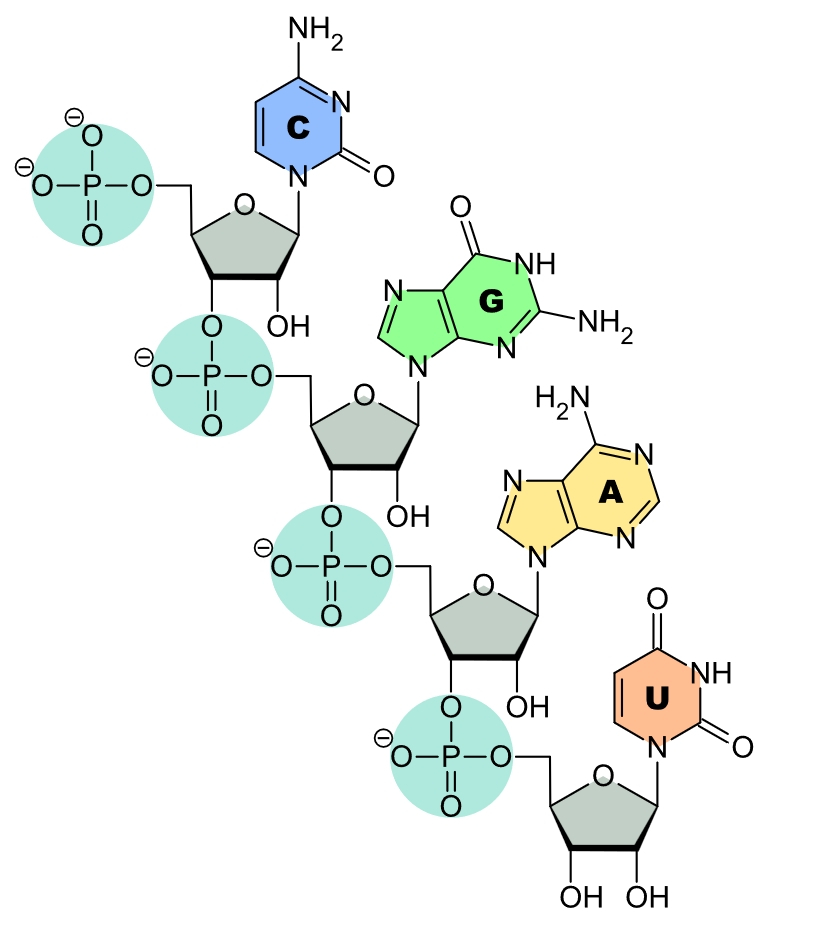
La molécule d’ARN est composée par un enchaînement de nucléotides. Les sucres (ribose, en gris) et phosphates (vert pâle) forment une chaîne monotone tandis que les base azotées présentent un ordre spécifique à la molécule, l’adénine est en jaune, l'uracyle en marron, la guanine en vert vif et la cytosine en bleu. L'ensemble adénine-ribose-phosphate (issu d'un nucléotide ATP) forme un demi barreau de la molécule bicaténaire d'ADN.
En effet, les acides nucléiques sont des polymères de nucléotides, chaque nucléotide étant lui-même constitué d’un sucre (ribose pour l’ARN, désoxyribose pour l’ADN), d’un acide phosphorique et d’une base azotée parmi 5 possibles (adénine, guanine, thymine, cytosine et uracile). L’ATP, le nucléotide possédant une adénine comme base azotée, participe donc à la fabrication de cette classe de macromolécules, au même titre que le GTP, le TTP, le CTP et l’UTP. Notons qu’en toute rigueur, ATP désigne le nucléotide à base d’adénine comportant du ribose, celui comportant du désoxyribose devant être appelé dATP.
L’ATP, molécule clef de l’énergétique cellulaire
Les êtres vivants tirent leur énergie de l’oxydation des nutriments, et ceci est vrai même pour les plantes chlorophylliennes (qui utilisent l’énergie des photons pour fabriquer de la matière organique qui pourra être oxydée ultérieurement). Cependant, l’énergie libérée lors de cette oxydation n’est pas directement utilisable par les cellules. Elle est captée par un intermédiaire qui, dans l’immense majorité des cas, se révèle être l’ATP. En effet, de très nombreux évènements cellulaires ou réactions métaboliques énergétiquement défavorables peuvent se dérouler grâce à la rupture de la liaison phosphodiester riche en énergie d’une molécule d’ATP. On peut citer différents exemples comme :
-
le transport actif effectué par l’ATPase sodium potassium, qui assure le maintien des déséquilibres ioniques de part et d’autre des membranes cellulaires, en permettant le passage du sodium et du potassium tous deux contre leurs gradients électrochimiques respectifs ;
-
La synthèse d’acétylcholine (l’un des principaux neuromédiateurs) à partir d’acétate et de choline selon la réaction
acétate + choline + ATP → Acétylcholine + ADP + Pi.
L’ATP est donc le donneur immédiat d’énergie libre de très loin le plus important dans les systèmes biologiques. Ce rôle d’intermédiaire, couplé au fait que les stocks d’ATP ne sont pas très importants, fait que cette molécule est soumise à un renouvellement intense, ce qui nécessite une production permanente, rapide et importante.
Si on fait le calcul chez l’homme, on estime que la consommation énergétique moyenne d’un individu est d’environ 8 360 kjoules (2 000 kcal) par 24 heures. Cette énergie, contenue dans les molécules organiques (généralement glucides et lipides), doit servir à fabriquer de l’ATP avant d’être utilisée par les cellules. Le rendement de cette synthèse est nécessairement inférieur à 1, de l’ordre de 0,5, le reste étant perdu sous forme de chaleur. C’est donc environ 4 180 kjoules (1 000 kcal) qui seront stockés transitoirement dans les molécules d’ATP puis utilisés par les cellules.
Or, l’hydrolyse d’une mole d’ATP fournit 30,5 kjoules (7,3 kcal) dans les conditions standard (concentration des différents composés intervenants de 1 M) mais près de 50 kjoules (12 kcal) en conditions physiologiques ( ref. 1). Les 4 180 kjoules fournis aux cellules correspondent donc à l’hydrolyse de 83,6 moles d’ATP, soit environ 46 kg. Sachant que le stock d’ATP/ADP, pour tout l’organisme, est d’environ 0,1 mole (autour de 50 g), cela implique qu’il soit renouvelé 920 fois par jour, soit approximativement une fois toutes les 1,5 minutes. Cette valeur est une moyenne pour un organisme et peut être largement inférieure pour une cellule, en cas de très forte demande énergétique (typiquement une cellule musculaire lors d’un effort intense), pour se chiffrer en secondes.
Ce calcul est bien sûr basé sur de nombreuses approximations et le résultat n’est pas à prendre comme un chiffre exact, mais il donne un ordre de grandeur et permet de conclure qualitativement que l’ATP est une molécule possédant un turnover extrêmement rapide.
L’ATP intervient dans de nombreuses voies de régulation
L’ATP participe largement à la régulation, et ceci à travers plusieurs mécanismes.
Le premier correspond à la phosphorylation. Par définition une réaction de phosphorylation correspond à l’ajout d’un groupement phosphate sur une molécule, ce groupement phosphate provenant d’un ATP qui sert alors de donneur. Les enzymes catalysant ce type de réaction sont appelées des kinases. Précisons qu’il faut distinguer cette réaction de la phosphatation, qui correspond également à un ajout d’un groupement phosphate sur une molécule mais à partir d’un phosphate inorganique. Les enzymes catalysant ce deuxième type de réaction sont appelées phosphorylases (les phosphatases sont des enzymes qui enlèvent un groupe phosphate d’une molécule).
Les phosphorylations ont souvent un impact sur l’activité des molécules ainsi phosphorylées. Le cas le plus classique est une activation ou une inhibition de l’activité d’une enzyme. On peut citer pour exemple la glycogène synthase, enzyme impliquée dans la synthèse du glycogène, qui est inactive à l’état phosphorylé et active à l’état déphosphorylé. Mais il est amusant de constater que la glycogène synthase kinase, l’enzyme qui phosphoryle la glycogène synthase, est, elle, inactive à l’état déphosphorylé et active à l’état phosphorylé. Il n’y a donc pas de règle entre état phosphorylé/déphosphorylé et activation/inactivation.
Un autre exemple qui démontre parfaitement cela provient de l’enzyme Cdk1 impliquée dans le contrôle du cycle cellulaire, en particulier de la transition phase G2/mitose. L’activité de la Cdk1 (Cyclin Dependant Kinase 1, Kinase 1 dépendante d’une cycline) est régulée par son association avec la cycline B, mais également par des phosphorylations. Cette molécule possède 3 sites de phosphorylation, dont deux sont inhibiteurs (Thr 14 et Tyr 15) et un activateur (Thr 161). Pour que l’enzyme soit active, il faut donc qu’elle soit associée à la cycline B, mais également qu’elle soit à la fois phosphorylée sur la Thr 161 et déphosphorylée sur les Thr 14 et Tyr 15.
Enfin les phosphorylations peuvent également réguler des molécules autres que des enzymes. Ainsi la phosphorylation de la chaîne légère de la myosine par la MLCK (Myosine Light Chain Kinase, kinase de la chaîne légère de la myosine) permet l’interaction actine-myosine et la contraction musculaire dans le muscle lisse.
L’ATP peut également intervenir directement dans la régulation de l’activité de certaines enzymes en tant qu’effecteur allostérique. D’une manière générale, un effecteur allostérique est une molécule qui se lie à une enzyme sur un site spécifique, qui ne participe pas à la réaction chimique catalysée, mais qui va modifier l’affinité de l’enzyme pour les substrats et du coup modifier son activité. Par définition, un effecteur allostérique ne peut exister que pour des enzymes allostériques, dont possédant plus d’une seule sous-unité. L’ATP est un effecteur allostérique classique de nombreuses enzymes intervenant dans le métabolisme, ce qui peut être interprété comme le moyen d’adapter finement et extrêmement rapidement l’activité métabolique à l’état énergétique des cellules.
On peut prendre l’exemple de la PFK (PhosphoFructoKinase) qui catalyse la réaction Fructose 6 Phosphate + ATP → Fructose 1,6 Diphosphate + ADP. Elle fait partie de la glycolyse et correspond à l’étape d’engagement des glucides dans la voie de production d’énergie, on parle d’enzyme-clé. A ce titre elle est donc finement régulée. C’est une enzyme tétramèrique (constituée de 4 sous-unités). L’ATP est un effecteur allostérique de cette enzyme. Pourtant c’est aussi un substrat. Mais l’ATP-substrat et l’ATP-effecteur allostérique se fixent sur des sites différents. L’ATP agit comme un inhibiteur de cette réaction, l’interprétation étant la suivante : lorsque la charge énergétique de la cellule est forte (rapport ATP/ADP élevé) il n’est pas nécessaire de produire beaucoup d’ATP. L’inhibition de la première étape qui mène vers la production d’ATP par l’ATP lui-même (via la glycolyse – le cycle de Krebs – et la chaîne de transporteurs d’électrons) va donc entraîner une diminution de cette production. Inversement, lorsque la charge énergétique de la cellule est faible (rapport ATP/ADP faible) il est nécessaire de produire beaucoup d’ATP. Comme la concentration en ATP est faible, il va y avoir une levée d’inhibition, ce qui revient à une activation de cette réaction et, par suite, de la production d’ATP.
Références
-
Complexité des équilibres dans l’hydrolyse de l’ATP. Document de cours de l’université de Montréal.



