

L’utilisation des différents types de revêtements externes des Vertébrés comme caractères morphologiques utilisables pour l’analyse phylogénétique implique la connaissance de leurs relations d’homologie. Cet article présente une comparaison des écailles, des plumes et des poils, au travers de données morphologiques, histologiques et développementales. Il clarifie ainsi les relations d’homologie existant entre ces différentes structures, affinées à l’aide de données génétiques.
Introduction
Un grand nombre de Vertébrés présente un revêtement externe : écaille, plume ou poil.
Le terme « écaille » recouvre à lui seul une grande variété de structures : écailles des poissons Actinoptérygiens, écailles des Chéloniens (le groupe des Tortues), écailles des Crocodiliens, des Lacertiliens (les Lézards) ou des Archosauriens (Dinosaures au sens classique + Oiseaux), ou encore écailles de certains Mammifères comme les tatous et les pangolins. D’autres groupes, en particulier de nombreux taxons fossiles, présentent également des structures superficielles rigides.
Utiliser, dans une analyse phylogénétique, le caractère morphologique « écaille », par exemple, exige de faire l’hypothèse a priori que toutes les structures désignées par ce terme sont homologues, c’est-à-dire qu’elles partagent la même structure et les mêmes relations avec les autres organes (homologie primaire ou de position). L’analyse phylogénétique permettra ensuite de vérifier si cette homologie structurale recouvre aussi une même origine embryologique, autrement dit que cette homologie de position est aussi une homologie de filiation (homologie secondaire).
Or l’homologie des différentes écailles des Vertébrés n’est pas vérifiée. Plus généralement, les différentes structures rigides qui recouvrent le corps des Vertébrés ont des organisations et des origines embryologiques différentes. Le présent article les passe en revue et revient sur leur évolution.
Les données paléontologiques
Les revêtements des poissons
Les premiers poissons du Siluro-dévonien, sans mâchoire, puis d’autres groupes de poissons gnathostomes aujourd’hui disparus, comme les Placodermes, avaient un corps sans squelette interne dur (donc probablement cartilagineux comme celui des Chondrichtyens ; requins et raies), mais recouvert d’écailles et, chez plusieurs groupes (Hétérostracés, Anaspides, Galéaspides), toute la région céphalique était recouverte d’une « armure » de plaques rigides.
Les poissons osseux (Actinoptérygiens et Sarcoptérygiens), qui présentent eux aussi un revêtement écailleux, apparaissent au Silurien et se diversifient au Dévonien. Vers la même époque, le Dévonien, apparaissent les premiers Chondrichtyens. La peau de ces poissons ne porte pas d’écailles macroscopiques et paraît lisse, mais elle est pourtant garnie de minuscules écailles, qui donnent à la peau des requins et raies actuels un toucher rugueux. Ces écailles diminuent les frottements de l’eau et augmentent l’efficacité de la nage.
Les revêtements des Amniotes
Au Carbonifère, apparaissent les premiers Amniotes, entièrement terrestres, et dont la peau limite les pertes d’eau par un revêtement plus imperméable que celui des Amphibiens et probablement écailleux comme celui des actuels Lézards, quoi que l’on ne dispose qu’exceptionnellement de restes de peau fossilisée.
Au Permien, se diversifient les reptiles Synapsides et les ancêtres des Mammifères, tandis que les Chéloniens et les Archosaures (Crocodiliens et Dinosauriens) apparaissent au Trias. Chez ces derniers, les écailles sont présentes et, chez les Tortues, les Crocodiliens ou dans plusieurs taxons de Dinosaures, se développent, en plus, des plaques protectrices beaucoup plus volumineuses.
Par ailleurs, on sait aujourd’hui que certains Dinosauriens du Crétacé présentaient des plumes semblables à celles des Oiseaux et que les premiers représentants des Oiseaux apparaissent au Jurassique. L’enracinement du clade des Oiseaux au sein du groupe des Dinosauriens est désormais assez largement accepté.
Les premiers Mammifères, issus des reptiles Synapsides du Permo-trias et probablement déjà recouverts de poils, remonteraient au Jurassique. Mais certains de leurs descendants actuels, comme le tatou ou le pangolin, sont recouverts d’écailles et de poils. S’agit-il là d’une réversion ou d’une convergence évolutive ?
Distinguer, entre ces diverses structures, les cas d’homologie des convergences évolutives demande d’en analyser d’abord l’histologie : une différence de structure histologique implique des développements embryonnaires différents, donc une relation d’analogie et non d’homologie.
Les données histologiques
L’analyse histologique des différents revêtements permet de distinguer deux types majeurs :
-
les éléments minéralisés, constitués d’une trame de collagène et de cristaux de phosphate de calcium (hydroxyapatite), semblables sur ce point aux os et aux dents 1
-
les éléments exclusivement organiques, mais rendus résistants par des processus de mort cellulaire et d’accumulation de protéines particulières dans les cellules mortes, les kératines (kératinisation).
Les structures apatitiques externes
Revêtement des « poissons cuirassés » fossiles
Les plaques qu’arboraient les « poissons cuirassés » du Siluro-dévonien ont été largement étudiées. Elles étaient constituées d’apatite, à l’instar des dents et des os, avec une structure pluristratifiée.
Chez les poissons cuirassés agnathes, le groupe des Hétérostracés par exemple, la tête était entièrement enserrée dans un petit nombre de grandes plaques, chacune constituée d’une base osseuse complexe appelée aspidine, surmontée d’une couche osseuse lacunaire portant des tubercules de dentine (figure ci-dessous). Il s’agissait donc d’un tissu d’origine entièrement mésodermique.
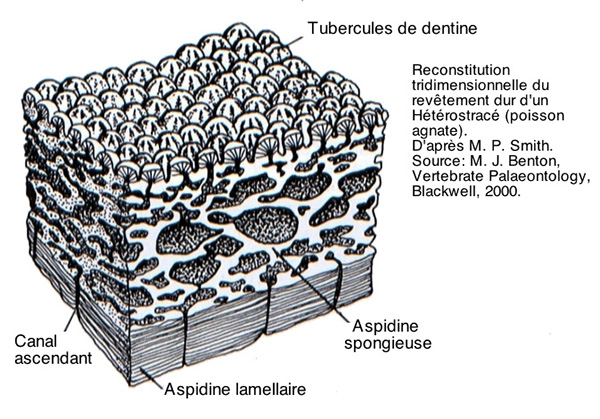
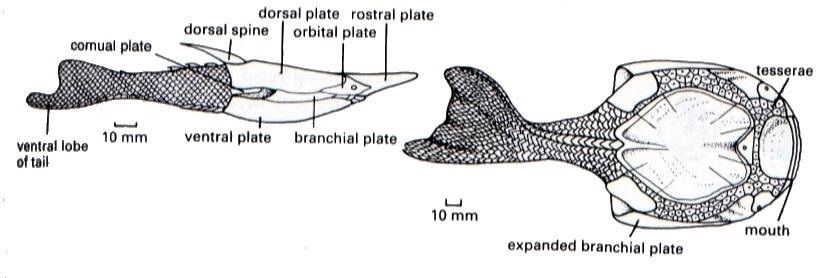
Pteraspis (à gauche) et Drepanaspis (à droite)
Au Dévonien, les Placodermes (un autre groupe de poissons, à mâchoire cette fois, donc inclus dans les Gnathostomes), ont eux aussi développé une région céphalique couverte de grandes plaques osseuses. Leurs mâchoires ne portaient pas de dents, mais des lames osseuses acérées. Le plus impressionnant, Dunkleosteus, devait occuper le sommet du réseau trophique des mers dévoniennes.

Notez les plaques pointues formant les « dents » et celles, plus petites, protégeant les orbites. Longueur de la tête: environ 1 m.
Écailles des Chondrichtyens
Chez les Chondrichtyens, la peau est percée de multiples petites écailles, appelées écailles placoïdes. Chacune d’entre elles s’enracine sur un bulbe de derme et comprend une cavité pulpaire entourée d’une épaisse couche de dentine, elle-même recouverte d’une couche d’émail. Cette structure est quasi-identique à celle d’une dent. Écaille placoïde et dent sont donc homologues morphologiquement.
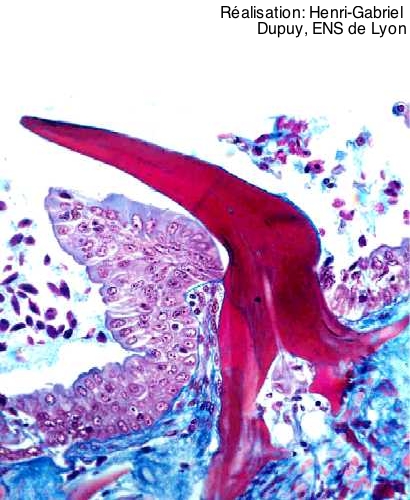
On distingue en rose vif la dentine, recouverte d’une fine couche d’émail (trait rose plus sombre). Elle enserre la cavité pulpaire et repose sur le derme (en bleu). L’écaille perce l’épiderme (en violet).
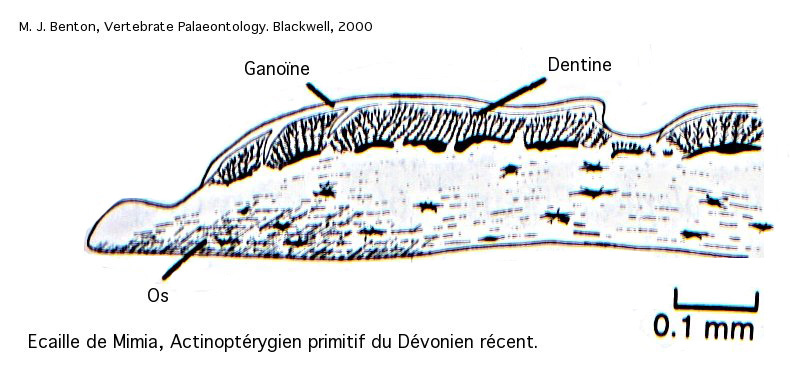
Écailles des Ostéichtyens
Les écailles des Ostéichtyens, les poissons osseux, diffèrent par leur structure de celles des Chondrichtyens. On distingue classiquement chez les Ostéichtyens deux types d’écailles : les écailles cosmoïdes et les écailles élasmoïdes.
Les écailles de type cosmoïde sont assez proches structurellement des plaques des poissons sans mâchoires, présentées plus haut : ce sont des éléments épais pluristratifiés, comprenant une lame osseuse basale surmontée d’une couche osseuse lacunaire, recouverte de dentine et parfois d’une lame d’un tissu semblable à l’émail par sa dureté. On trouve ce type d’écaille dans divers groupes d’Ostéichtyens, en particulier chez les poissons dévoniens comme Eusthenopteron, parent du Cœlacanthe, et chez les ancêtres des Dipneustes.
Chez les Actinoptérygiens les plus anciens, les écailles ressemblent encore à l’écaille cosmoïde, mais le tissu dur superficiel se distingue de l’émail par sa structure stratifiée. Ce tissu, appelé ganoïde, est caractéristique des Actinoptérygiens.
Chez les Actinoptérygiens plus récents, par contre, l’écaille est de type élasmoïde, c’est-à-dire beaucoup plus mince, partiellement incluse dans le derme. Elle est constituée, tout d’abord, d’une plaque basale de collagène, plus ou moins minéralisée, comprenant plusieurs couches de fibres de collagène (l’orientation des fibres changeant d’une couche à l’autre, cela produit une structure en contreplaqué). Ce tissu particulier est appelé élasmodine. Au-dessus, se trouve une seconde plaque, la couche externe, plus fine et mieux minéralisée, composée de fibres de collagène organisées en réseau. Le tout est recouvert, juste sous l’épiderme, d’une fine lame très minéralisée (la couche limitante), sans fibres de collagène.
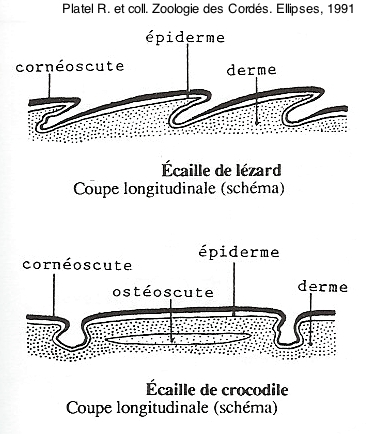
Protections dermiques osseuses des Tortues et des Crocodiliens
La carapace (la région dorsale) et le plastron (la région ventrale) des Tortues sont des structures complexes, qui comprennent une armature osseuse dermique et une couverture épidermique kératinisée. Cette enveloppe est typique de ce groupe et aurait été acquise relativement tôt dans son histoire, qui commence au Trias récent.
La carapace est constituée de plaques osseuses formées au sein du derme, attachées aux vertèbres et aux côtes. Ces plaques sont recouvertes d’un épiderme kératinisé, lui aussi divisé en plaques, mais ces dernières sont dépourvues de relation géométrique avec les plaques osseuses.
Les productions épidermiques kératinisées
Les Amniotes ont acquis, au cours de leur adaptation au milieu terrestre, un épiderme particulier, limité extérieurement par plusieurs couches de cellules mortes remplies de kératines qui restreignent considérablement le passage des gaz. Les cellules mortes les plus superficielles, les squames, se détachent et sont remplacées par les cellules sous-jacentes repoussées vers l’extérieur par la prolifération des cellules basales de l’épiderme. Ces cellules basales prolifèrent et certaines alimentent les couches sus-jacentes, où elles se différencient pour former plusieurs couches de cellules volumineuses, reliées entre elles par de nombreux desmosomes, et qui synthétisent des filaments de kératine (cellules épineuses)1.
Chez les Mammifères, les cellules de la couche suivante prennent un aspect granuleux dû à l’accumulation d’amas de kératohyaline, un assemblage de protéines (en particulier la fillagrine) impliqué dans le tassement de la cellule et l’apparition de liens entre les faisceaux de kératine. Ces cellules dégénèrent en progressant vers la surface de l’épiderme (disparition du noyau et du cytoplasme) pour ne laisser qu’une enveloppe cellulaire aplatie et fortement kératinisée.
Écailles et plumes des Sauropsides
Chez tous les reptiles Sauropsides (Tortues + Crocodiles + Lézards et apparentés + Serpents + Dinosauriens (dont Oiseaux)), la kératine est présente sous deux formes : la kératine alpha, flexible, assure la souplesse de la peau et est seule présente entre les écailles, alors que la forme bêta est plus rigide et se concentre au niveau des écailles.
Selon les groupes, les couches cellulaires à kératine bêta peuvent recouvrir ou remplacer les couches de cellules à kératine alpha. Dans tous les cas, les régions bordières de l’écaille ne présentent que la couche molle, assurant ainsi une relative indépendance des écailles entre elles.
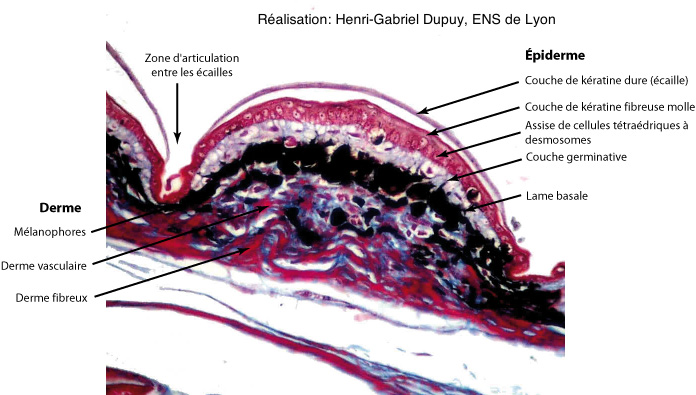
Pour certains groupes de reptiles Archosauriens, comme les Crocodiliens ou certains Dinosauriens, les plus grosses écailles du dos et de la queue recouvrent une plaque osseuse dermique (ostéoscute). Chez les Chéloniens (les Tortues), ces mêmes ostéoscutes, particulièrement développés, sont soudés entre eux et aux vertèbres et constituent la carapace et le plastron.
Les écailles présentes au niveau des pattes, chez les Oiseaux, ont une structure quasi-identique à celle des écailles des autres reptiles.
Les plumes des Oiseaux, quant à elles, sont aussi des phanères (productions tégumentaires issues de l’ectoderme) kératinisés, d’origine essentiellement épidermiques [8]. Elles se composent de cellules kératinisées formant un rachis central d’où partent des barbes elles-mêmes ramifiées en barbules. Le rachis s’insère dans la peau au sein d’un fourreau épidermique, le calamus, enfoncé dans le derme.
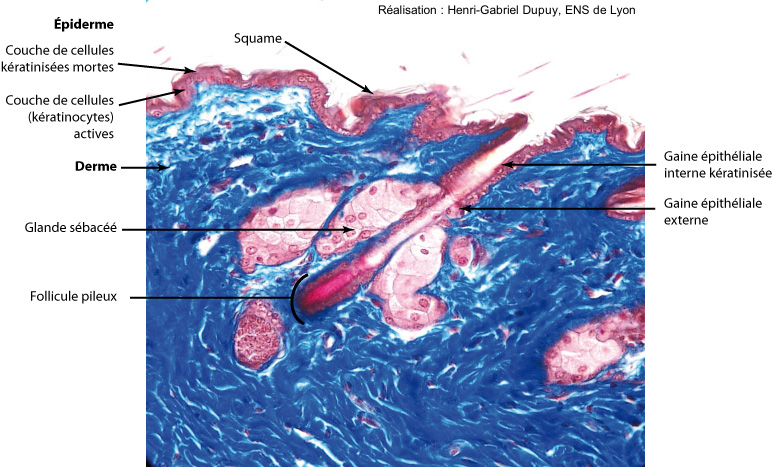
Poils des Mammifères
Les poils des Mammifères sont composés d’un tube kératinisé formé dans une poche épidermique invaginée dans le derme. C’est donc une structure presque entièrement épidermique. Le développement d’un poil résulte d’une interaction entre quelques cellules dermiques, constituant la papille dermique du follicule pileux, et la couche épidermique.
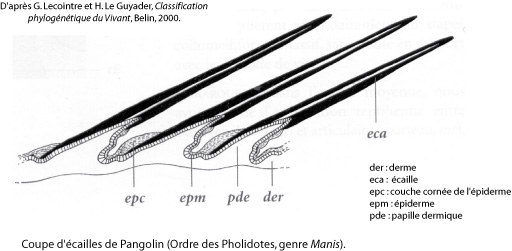
Quelques genres de Mammifères présentent, en plus des poils ou des griffes, des revêtements durs particuliers. C’est le cas des représentants de l’ordre des Pholidotes, les Pangolins (genre Manis), dont le corps est recouvert de larges écailles chevauchantes, kératinisées, et celui des Tatous (genre Dasypus, ordre des Edentés ou Xénarthres)1.

Mais la relative parenté phylogénétique des Pholidotes et des Xénarthres (indiquée par d’autres caractères morphologiques et moléculaires) ne recouvre pas d’homologie structurale de leur revêtement. L’armure des tatous est composée de plaques osseuses recouvertes par l’épiderme kératinisé (comme la carapace des tortues), alors que les écailles du pangolin ne sont constituées que de kératine et diffèrent peu, par leur structure, d’autres éléments très kératinisés dérivés des poils, comme les épines du hérisson ou du porc-épic [4] (figure ci-dessous).
Si l’on s’en tient aux seules caractéristiques de la peau, une différence majeure, séparant plus clairement Synapsides et Sauropsides, est la structure de l’épiderme.
Chez les Sauropsides, la limitation des pertes en eau a été obtenue par le développement d’une couche épidermique particulière, contenant de la kératine bêta et recouvrant la couche à kératine alpha, plus molle. C’est cette kératine bêta qui constitue la couche dure de l’écaille. Par contre, chez les Synapsides, dans le rameau des Mammifères, cette même adaptation a été obtenue par l’épaississement de la couche à kératine alpha et son renforcement par des protéines riches en acides aminés soufrés et en histidines (HSP, Histidine-rich proteins) [12].
Cet inventaire des structures épidermiques souligne combien le développement d’os dermiques est fréquent parmi les Amniotes, puisqu’il concerne de nombreux groupes : les Tortues, les Crocodiles, plusieurs groupes de Dinosaures ou les Tatous. De très nombreux caractères morphologiques et moléculaires séparant nettement, par ailleurs, ces groupes dans les phylogénies ; on peut affirmer que ces ossifications dermiques sont des convergences.
Les données du développement
Le développement des écailles des Chondrichtyens et des Ostéichtyens
On a dit que l’écaille placoïde des Chondrichtyens était histologiquement semblable à une dent. L’étude de ces écailles, chez les Sélaciens en particulier, a démontré que leurs similitudes morphologiques avec les dents recouvraient aussi des similitudes de développement.
L’émail est un tissu d’origine ectodermique, formé par l’épiderme, alors que la dentine est synthétisée par des cellules du derme. L’écaille placoïde, comme la dent, est donc un tissu épithélio-conjonctif, ou ecto-mésodermique. Certains auteurs [11] proposent de remplacer le terme d’écailles placoïdes par celui d’odontodes (de odontos, la dent) pour souligner cette similitude.
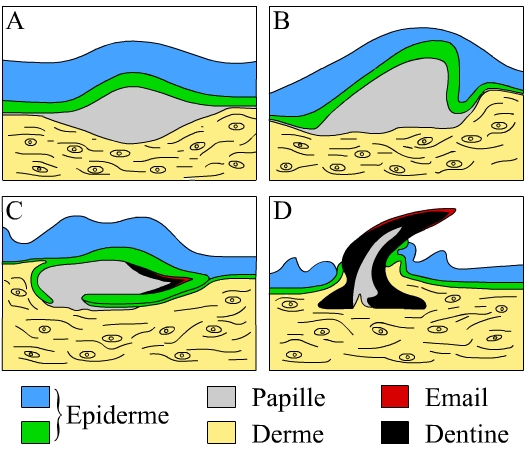
A et B Formation de la papille dermique.
C Dépôt des tissus calcifiés: l’émail et la dentine.
D L’écaille transperce la couche épidermique.
Écailles placoïdes et dents sont ainsi des organes homologues au sens phylogénétique, c’est-à-dire de même origine embryologique. Les dents dériveraient d’écailles placoïdes internalisées dans la cavité buccale. D’un point de vue évolutif, on peut supposer que l’odontode représente l’état ancestral que les Chondrichtyens seraient les seuls Vertébrés à avoir conservé, parallèlement à l’acquisition des dents.
L’origine embryologique des écailles des Ostéichtyens a également été analysée, surtout depuis que le poisson-zèbre (zebrafish) est devenu, avec le nématode (Cænorhabditis elegans), la drosophile (Drosophila melanogaster) ou la souris (Mus domesticus), l’un des animaux modèles des chercheurs en biologie du développement.
L’écaille cosmoïde semble proche, par sa structure, de l’odontode des Chondrichtyens, si l’on admet, comme plusieurs auteurs [11], que la ganoïde n’est qu’une variante de l’émail, donc un tissu d’origine ectodermique. Écaille placoïde et écaille cosmoïde seraient bien, alors, des organes homologues 1.
En ce qui concerne l’écaille élasmoïde, sa structure histologique diffère en apparence de celle de l’écaille placoïde ou des dents. Elle ne présente pas, en particulier, de base osseuse. Son développement est aussi légèrement différent de celui des écailles placoïdes. Les écailles élasmoïdes des Ostéichtyens se développent selon une séquence bien établie : dans une étape assez tardive du développement, des cellules de type fibroblastes envahissent la partie supérieure du derme. À certains endroits de la peau seulement, selon un motif en damier, ces cellules se multiplient et se différencient pour former une « papille dermique », de plusieurs couches de cellules. Ce schéma rappelle ce que l’on observe chez les Chondrichtyens, mais, chez ces derniers, la position de ces papilles dermiques paraît aléatoire et non pré-organisée comme elle l’est chez les Ostéichtyens.
Entre les deux couches supérieures de cellules de la papille, juste sous l’épiderme, apparaît d’abord la couche collagénique externe. Un peu plus tard, les cellules les plus profondes de la papille se différencient et commencent à synthétiser l’élasmodine. La couche limite minéralisée apparaît en dernier. L’ensemble de la papille et de l’écaille en formation s’étend latéralement et s’incline progressivement tandis que l’épiderme sus-jacent se plisse : l’écaille s’enracine donc dans le derme, du côté antérieur, et finit par traverser l’épiderme, du côté postérieur.
Selon Sire et al. [11], les écailles élasmoïdes seraient bien homologues des odontodes et des écailles cosmoïdes. Ces auteurs proposent que la couche limitante soit l’homologue de l’émail, tandis que les deux couches collagéniques seraient des homologues légèrement différenciés de la dentine. L’évolution de l’écaille cosmoïde présente chez les Ostéichtyens les plus anciens (et conservée dans le groupe des Actinoptérygiens) aurait conduit, chez les Téléostéens, à l’écaille élasmoïde, par disparition de la base osseuse, remplacée fonctionnellement par l’élasmodine, dérivée de la dentine.
Ainsi, pour ces auteurs, les écailles des Chondrichtyens, comme des Ostéichtyens, et les dents de tous les Vertébrés seraient toutes des structures mixtes ecto-mésodermiques, formées à la limite de l’épiderme et du derme, et dont le développement nécessiterait des interactions entre ces deux tissus. Toutes dériveraient d’une structure ancestrale commune, l’odontode, semblable aux écailles placoïdes des Chondrichtyens. On aurait donc les relations d’homologie suivantes entre les tissus des écailles et des dents :
| Écaille placoïde | Dent | Écaille cosmoïde | Écaille élasmoïde |
|---|---|---|---|
| Émail | Émail | Ganoïne | Couche limite |
| Dentine | Dentine | Dentine | Couche collagénique externe |
| Élasmodine |
Démontrer l’exactitude de ces relations d’homologie entre ces structures nécessiterait cependant encore de vérifier que les mêmes voies génétiques, donc des gènes homologues, sont bien impliquées dans le développement de toutes ces « écailles ».
Le développement des structures kératinisées
Chez la plupart des Sauropsides qui ne présentent pas de spécialisations ou d’ornementations particulières, les écailles ne sont que des protubérances de l’épiderme qui s’invaginent vers l’extérieur au cours du développement, prennent dans certains cas une forme arquée, par prolifération dissymétrique des cellules, et développent une couche kératinisée dure. Cette zone écailleuse se renouvelle beaucoup plus lentement que le reste de l’épiderme.
Le poil des Mammifères, comme la plume des Oiseaux, apparaît initialement sur la peau sous la forme d’une prolifération localisée de cellules de l’épiderme qui définit une région épaissie, la placode [12]. Cette placode distingue le développement des poils et des plumes de celui des écailles reptiliennes. Elle représente un « centre spécialisé de prolifération et de différenciation de cellules épidermiques et dermiques » [7]. C’est une caractéristique des annexes tégumentaires, terme qui regroupe donc ‘plumes des Oiseaux’ et ‘poils des Mammifères’. Existe-t-il néanmoins des relations entre écailles reptiliennes et plumes d’une part, écailles reptiliennes et poils d’autre part ?
Les plumes des oiseaux se forment à partir d’une placode qui s’allonge, d’abord, en un germe faisant saillie à la surface de la peau, avant de s’enfoncer dans le derme pour donner un follicule comportant un revêtement épidermique et une pulpe centrale dermique. Les cellules épidermiques internes du revêtement se différencient pour former le rachis, les barbes et les barbules, qui se développent en hélice autour du derme, tandis que les cellules externes forment le fourreau du calamus. À mesure qu’elles sortent du fourreau, les barbes s’étalent et donnent à la plume sa structure plane.
Ainsi, le développement des plumes diffère de celui des écailles des autres reptiles : le germe est tubulaire et les barbes se développent d’abord en tube avant de se déployer latéralement. Il ne s’agirait donc pas d’une écaille reptilienne allongée, redressée et découpée en barbes comme le supposaient les premiers modèles, mais du résultat d’une série d’innovations embryologiques.
Cette interprétation s’appuie sur les récentes découvertes fossiles : en effet, parmi les « dinosaures emplumés » récemment décrits, certains ne présentent qu’un type de plume (plume de duvet, rémiges), d’autres plusieurs types différents, comme les Oiseaux, et surtout certains dinosaures présentent à l’état adulte des structures équivalentes à certaines étapes du développement embryologique actuel de la plume.
Chaque étape embryologique du développement de la plume correspond à une innovation évolutive spécifique d’un groupe monophylétique de Dinosauriens : autrement dit, on peut désormais replacer sur l’arbre phylogénétique des Dinosauriens, en tant qu’innovation évolutive, l’apparition de chaque étape embryologique aboutissant à une plume [8]. Ces dinosaures, comme les Oiseaux aujourd’hui, arboraient donc deux revêtements différents, les écailles, restreintes aux membres inférieurs chez les Oiseaux, et les plumes.
Dans le cas du poil des Mammifères, les cellules épidermiques de la placode s’invaginent dans le derme pour former le follicule pileux. Les écailles des reptiles actuels et les poils des Mammifères diffèrent donc sensiblement dans leur structure, comme dans leur développement : le développement des écailles ne montre pas de formation d’une placode ni d’un follicule, le poil comme la peau des mammifères ne possède pas de kératine bêta (ce qui pourrait être une perte de caractère dans la branche des reptiles Synapsides contenant les Mammifères).
Contrairement au cas de la plume, on ne dispose pas d’un registre fossile suffisant pour éclaircir les relations évolutives entre écailles et poils. On ignore également si le poil est une innovation propre au groupe des Mammifères ou s’il était déjà apparu chez les Synapsides avant l’individualisation des Mammifères. Néanmoins, une hypothèse présente le poil comme une modification d’un type d’écaille reptilienne [12] : certains reptiles arborent des écailles spécialisées portant, au niveau de la région charnière reliant deux écailles, une épine sensorielle. Des modifications du développement conduisant à une invagination de cette épine et à la perte de l’écaille protectrice pourraient expliquer l’apparition du poil mammalien.
Ainsi, plumes et poils proviennent tous deux d’une invagination épidermique donnant un follicule dans le derme, mais ces deux structures sont apparues indépendamment et elles ne proviennent pas d’une modification directe des écailles reptiliennes.
Écaille des poissons et revêtements tégumentaires des Amniotes apparaissent finalement comme des formations distinctes, tant structurellement qu’embryologiquement. Elles ne sont donc en rien des caractères homologues. Cela signifie aussi que l’on ne peut considérer la peau des amphibiens comme le résultat d’une « perte du caractère ‘écaille’ conservé par ailleurs chez les Amniotes ». Les amphibiens ont effectivement perdu la capacité à développer des écailles dérivées de l’odontode ancestral pour ne garder que la peau dépourvue de couche kératinisée de leurs ancêtres aquatiques. Ils ont, par ailleurs, développé des glandes muqueuses, qui limitent le dessèchement de l’épiderme. L’acquisition de ces glandes muqueuses chez les Amphibiens et de la couche cornée de l’épiderme ( stratum corneum) chez les Amniotes sont deux innovations évolutives, sans relation de descendance avec les écailles des poissons.
Quelques données génétiques
Les similitudes des premières étapes du développement des plumes et des poils, avec l’initiation de la placode, laissent penser que certains gènes, intervenant dans la structuration générale de l’épiderme et les interactions derme-épiderme, ont pu être recrutés dans ces deux développements.
Par contre, les voies de développement des écailles des poissons (productions dérivées du derme) et des écailles reptiliennes, des poils et des plumes (structures dérivées de l’épiderme) étaient supposées distinctes. Cependant, une découverte récente a remis en cause cette supposition.
Kondo et al. [9][10] ont identifié un gène du développement qui s’avère impliqué à la fois dans la formation des écailles des poissons et des poils des mammifères. Ces chercheurs ont étudié chez le Médaka, un petit Téléostéen proche du poisson-zèbre, une mutation qui se traduit par la disparition quasi-complète des écailles. Or le gène impliqué, appelé rs-3, est connu également chez la souris, où sa mutation entraîne l’absence de poils. Ce gène interviendrait dans les séquences les plus précoces du développement des structures tégumentaires : il code un récepteur transmembranaire (EDAR, récepteur de l’ectodysplasine-A).
Selon Kondo et al. [10], poils et écailles partagent la particularité d’être disposés selon un arrangement ordonné sur le corps. EDAR serait donc une molécule conservée, impliquée dans la définition de cette disposition organisée des structures de l’épiderme. L’intervention d’un même gène dans ces deux groupes d’animaux n’implique pas, cependant, une relation d’homologie entre écaille des poissons et poils. Il s’agit d’une convergence évolutive où le même gène, qui intervient très en amont dans le développement des structures tégumentaires, s’est trouvé recruté dans les deux processus.
Conclusion
Les revêtements externes des Vertébrés sont relativement diversifiés et il est donc nécessaire de clarifier leurs relations en tant que caractère morphologique utilisable pour l’analyse phylogénétique. Comme on l’a vu, les données morphologiques, histologiques et développementales éclairent ces relations :
-
Les structures dermiques osseuses (carapaces, plaques) sont fréquentes chez les Vertébrés et peuvent apparaître dans des groupes par ailleurs bien distincts phylogénétiquement (les dinosaures Ankylosauridés, les Ostracodermes et les Tatous, par exemple). Elles ne représentent donc, souvent, que des convergences évolutives.
-
Écailles placoïdes et dents sont des organes homologues au sens phylogénétique ; elles partagent une origine embryologique commune.
-
Les écailles des Ostéichtyens seraient, elles aussi, homologues des écailles placoïdes et en dériveraient.
-
Par contre, les structures kératinisées des Tétrapodes, d’origine ectodermique, n’ont pas de lien direct avec les écailles ecto-mésodermiques des poissons et n’en sont que des analogues morphologiques.
-
Parmi les Tétrapodes, les productions tégumentaires autres que les écailles reptiliennes, à savoir poils et plumes, ne sont pas non plus homologues entre elles. Quoiqu’elles comprennent les mêmes catégories de tissu, leurs structures et leurs modalités de développement diffèrent. De plus, le registre fossile en fait deux caractères exclusifs : aucun fossile et aucun organisme n’a présenté simultanément des poils et des plumes. Poils et plumes représentent donc deux acquisitions indépendantes. Le poil serait apparu dans un groupe de reptiles Synapsides à l’origine des Mammifères, peut-être dès le Permien, alors que la plume est une acquisition d’un groupe de Dinosaures théropodes, survenue au cours du Jurassique.
-
Par ailleurs, l’hypothèse d’homologie entre la plume et l’écaille reptilienne est contestée aujourd’hui par la combinaison des dernières découvertes fossiles et de l’étude du développement embryonnaire de la plume. Cependant, le poil pourrait être issu d’une transformation évolutive de certaines écailles reptiliennes spécialisées. Quoi qu’il en soit, même si elles ne sont pas phylogénétiquement homologues, plumes, poils et écailles kératinisées découlent chacune d’une modification des interactions entre cellules dermiques et épidermiques au cours du développement.
Bibliographie
- Alberts B. et coll., Biologie moléculaire de la cellule. Flammarion Médecine-Sciences, 1995.
- Platel R., Zoologie des Cordés. Ellipses, 1991.
- Lautrou A., Anatomie dentaire. Masson, 1997.
- Lecointre G. & Le Guyader H., Classification phylogénétique du vivant. Belin, 2000.
- Benton M. J., Vertebrate Palaeontology 2nd ed., Blackwell Science, 2000.
- Encyclopaedia Universalis, article Poissons et Actinopterygiens.
- Alibardi L. (2004) Dermo-epidermal interactions in reptilian scales : Speculations on the evolution of scales, feathers, and hairs. Journal of Experimental Zoology Part B : Molecular and Developmental Evolution 302B(4), 365-383.
- Prum R. et Brush A. (2005), Les plumes des dinosaures. Pour la Science, dossier spécial Le monde des Dinosaures, Juillet/Septembre 2005.
- Sharpe P. T. (2001) Fish scale development : Hair today, teeth and scales yesterday ? Current Biology 11(18), R751-R752.
- Kondo et al. (2001), The medaka rs-3 locus required for scale development encodes ectodysplasin-A receptor, Current Biology 11, 1202-1206.
- Sire J. Y. and Akimenko M. A. (2004) Scale development in fish : a review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio) International Journal od Developmental Biology 48(2-3), 233-247.
- Wu et al. (2004) Evo-Devo of Amniote integuments and appendages. Int. J. Dev. Biol. 48 : 249-270.
Crédits

