

Une équipe de chercheurs, grâce à un travail collaboratif impliquant 36 personnes travaillant dans 17 institutions, a reconstitué le portrait de la fleur de l’ancêtre des Angiospermes, grâce à un large jeu de données, et en utilisant des méthodes alternatives par rapport à la méthode de parcimonie. Cette reconstruction montre notamment que la fleur de cet ancêtre était bisexuée (ou hermaphrodite).
Référence : Sauquet, Hervé et al. “The ancestral flower of angiosperms and its early diversification.” Nature communications (2017).
La fleur des Angiospermes actuelles

a : fleur avec un périanthe indifférencié, composé de tépales. b : fleur avec un périanthe différencié en sépales et pétales.
Photo Lysimachia : Donald Hobern, CC BY 2.0
Les Angiospermes, ou plantes à fleurs, représentent le groupe de végétaux majoritaire dans les écosystèmes terrestres : ce sont les principaux producteurs primaires en milieu continental, et dans la plupart des milieux les êtres vivants dépendent de cette source de nourriture, de manière directe ou indirecte.
Les Angiospermes ont colonisé tous les milieux, des déserts chauds et froids aux prairies boréales, et certaines espèces sont même retournées secondairement en milieu aquatique. Les Angiospermes présentent donc une grande diversité en termes d’adaptations et de modes de vie, et cette diversité se retrouve aussi au niveau morphologique. Un des traits les plus variables concerne la fleur, c’est-à-dire une structure reproductrice généralement bisexuée (mâle et femelle) qui constitue une innovation évolutive pour ce groupe (Fig. 1). Les fleurs des Angiospermes présentent un grand nombre de tailles, d’organisations et de formes différentes (Fig. 4 à 6). Avant les résultats obtenus en phylogénie moléculaire, les systèmes de classification proposés pour les Angiospermes étaient essentiellement basés sur la structure de la fleur.
Les Angiospermes font partie du grand groupe des Spermatophytes (dont les représentants actuels possèdent des graines), qui comprend notamment les Conifères (ou Pinales ; voir Fig. 2).
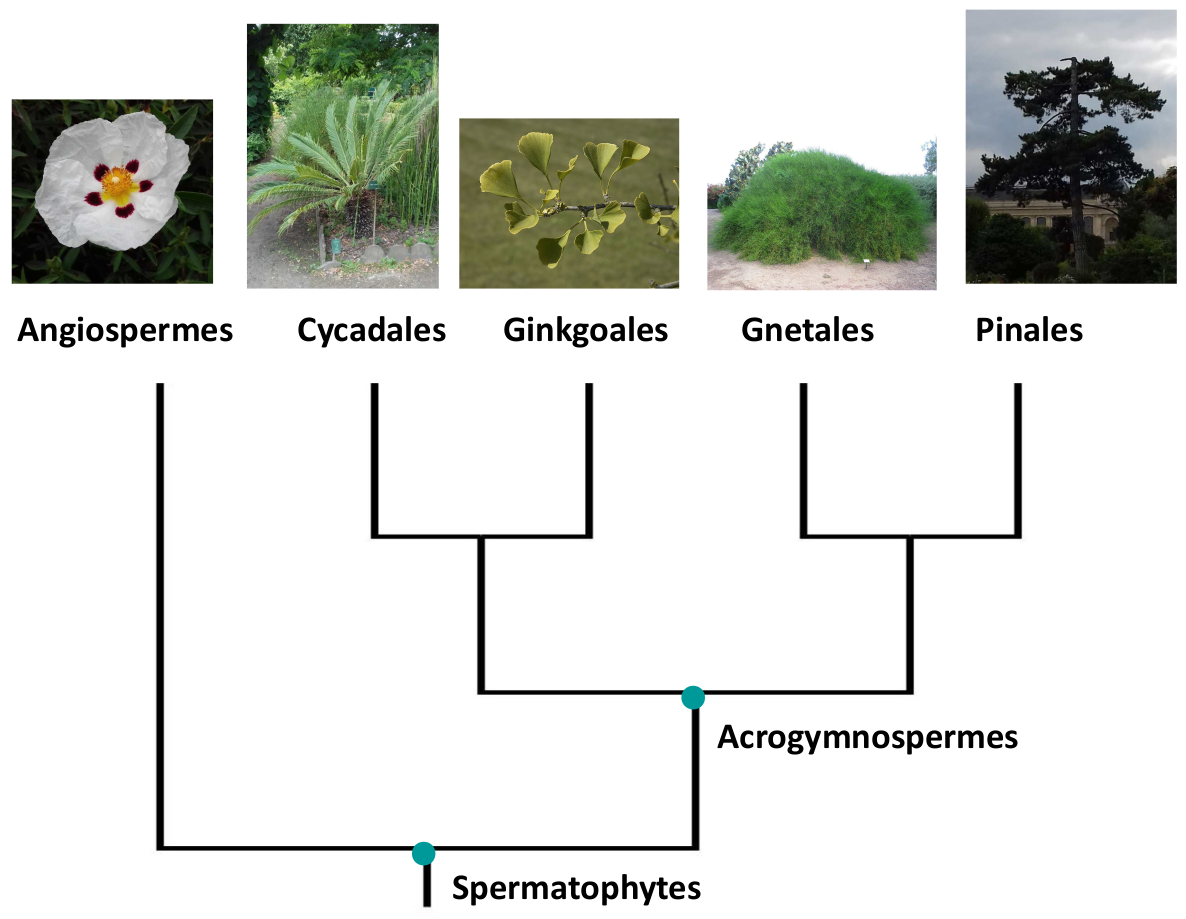
Légende : Représentés sur les photos, de gauche à droite : ciste, cycas, ginkgo, éphedra, pin. Il est à noter que les relations présentées ici reflètent l’opinion majoritaire au sein des spécialistes, mais ces relations restent à l’heure actuelle incertaines.
Photo Ginkgo : Wouter Hagens, domaine public
Photo Ephedra : Daderot, domaine public
On a longtemps classé les Angiospermes en Monocotylédones et Dicotylédones (classification basée sur le nombre de cotylédons dans la graine). Cependant des études ultérieures (APG I à IV) ont montré que le groupe des Dicotylédones n’était pas monophylétique, ce qui a conduit à le diviser en plusieurs groupes : les Amborellales, les Nymphaeales, les Austrobaileyales, les Magnoliidées et les Eudicotylédones (Fig. 3). Le groupe des Monocotylédones a quant à lui été conservé car il forme un clade (un groupe avec un ancêtre commun et tous ses descendants : Fig. 3).
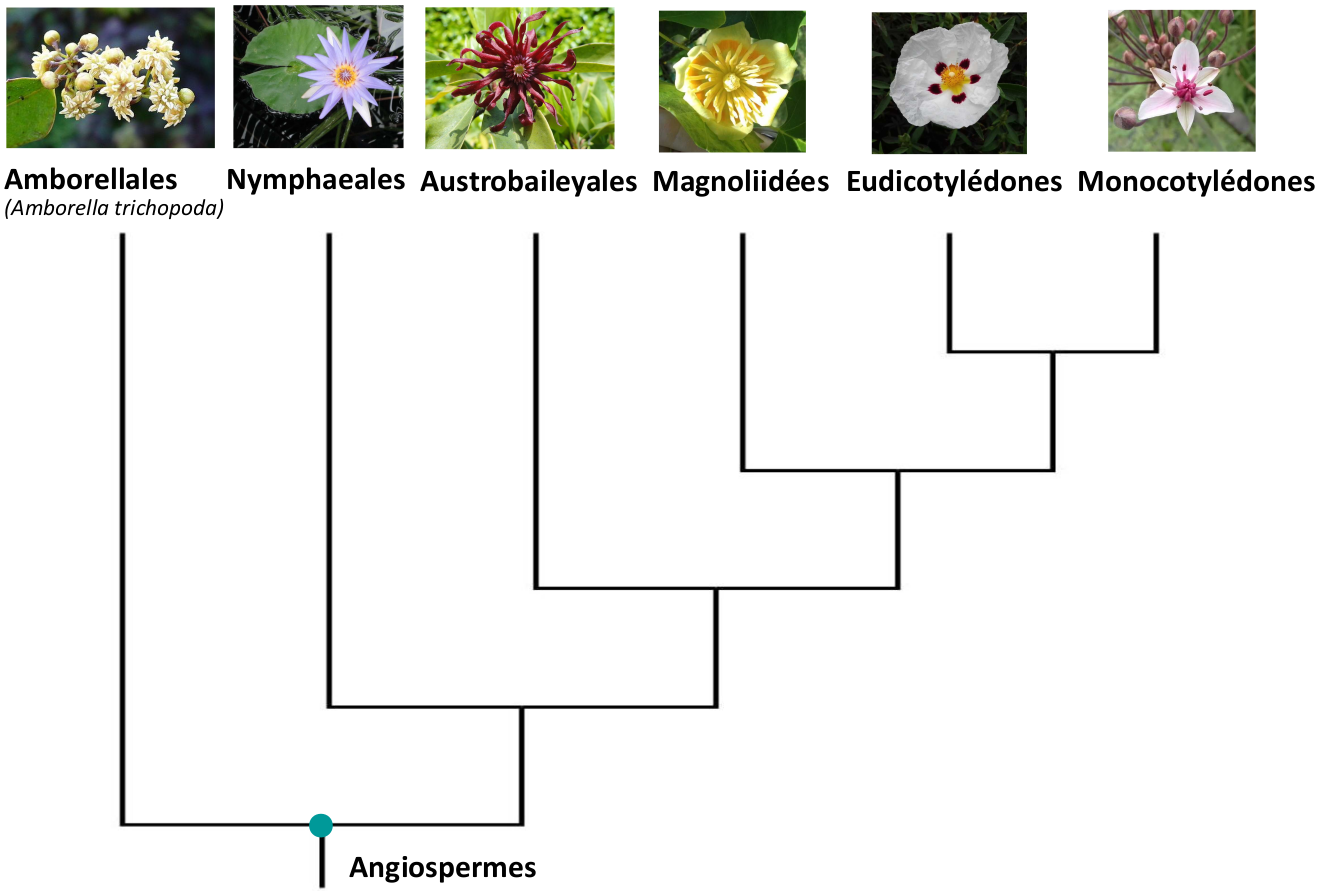
Représentés sur les photos, de gauche à droite : Amborella, nymphéa, badianier (espèce proche de l’anis étoilé, du genre Illicium), tulipier, ciste, jonc fleuri (Butomus). Il est à noter que les relations présentées ici reflètent l’opinion majoritaire au sein des spécialistes, mais ces relations restent à l’heure actuelle incertaines.
Photo Amborella : Scott Zona from USA, CC BY 2.0
Photo Illicium : Daderot, domaine public
Les Amborellales, les Nymphaeales et les Austrobaileyales, tout comme les Magnoliidées, comportent des familles ayant assez peu de représentants sous nos latitudes, mais on peut tout de même citer les nénuphars et les nymphéas (resp. genres Nuphar et Nymphaea, Nymphaeaceae), les badianiers (genre Illicium, dont une des espèces produit l’anis étoilé), le laurier sauce (Laurus nobilis, Lauraceae), les magnolias (Magnolia grandiflora, Magnoliaceae), le tulipier (Liriodendron tulipifera), parmi les espèces les plus connues, certaines ayant été introduites en Europe (Fig. 4).

De gauche à droite : Illicium floridanum, Nymphaea sp. et Liriodendron tulipifera.

En haut à gauche : jonc fleuri Butomus umbellatus, en bas à gauche : ophrys abeille Ophrys apifera, à droite : riz cultivé Oryza sativa.
Photo Oryza : Guy Sabattier, banque nationale SVT (utilisation autorisée dans un but pédagogique)
Les Monocotylédones comprennent environ 20 % des espèces actuelles d’Angiospermes, et sont caractérisées par la perte secondaire (ou réversion) du cambium (méristème permettant la formation de bois et de liber) et de l’un des deux cotylédons. Elles comprennent un grand nombre de familles qui nous sont familières, comme les Poaceae (famille des céréales comme le blé, le riz, le maïs…) ou les Orchidaceae, parmi tant d'autres (Fig. 5).

De gauche à droite : ciste à gomme Cistus ladanifer, liseron de mer Calystegia soldanella, violette tricolore Viola tricolor, vipérine commune Echium vulgare.
Photo Viola : Wisniowy, domaine public
Enfin les Eudicotylédones représentent environ 75 % des espèces actuelles d’Angiospermes, et constituent la majorité des espèces de la flore française : toutes les espèces d'arbres produisant des fleurs (chêne, hêtre et châtaignier chez les Fagaceae, charme chez les Betulaceae), les représentants des Asteraceae, des Rosacaeae, des Fabaceae, des Apiaceae… Les Eudicotylédones sont notamment caractérisées par la présence d'un pollen à 3 apertures (zones amincies de la paroi permettant les échanges et la formation du tube pollinique).
Quelles méthodes pour reconstituer la fleur ancestrale ?
La question de la structure des premières fleurs intéresse depuis longtemps les botanistes, et certains points comme le mode de pollinisation, ou le sexe des fleurs (unisexué ou bisexué) ont fait l'objet de longues discussions. S’il est établi que les Angiospermes se sont diversifiées pendant l'ère secondaire, les débuts de leur histoire évolutive sont encore mal connus (Sauquet & Magallón 2018). Les premiers pollens d'Angiospermes apparaissent au début du Crétacé (il y a environ 130 millions d’années), suivis par les premiers macro-fossiles (certains fossiles plus anciens pourraient appartenir à la lignée des Angiospermes, mais ils ne font pas l’unanimité parmi les spécialistes ; voir par exemple Herendeen et al., 2017). Cependant les phylogénies moléculaires donnent des dates plus anciennes, comprises entre 250 et 140 millions d'années pour l’ancêtre commun le plus récent de toutes les espèces actuelles.
Différentes approches permettent de reconstituer le portrait-robot de cet ancêtre commun. La première est l’étude des fossiles. Une limite de cette approche est que les premiers fossiles de fleurs datent d’au moins 10 millions d’années après l’apparition du groupe. Ces fleurs fossiles peuvent donc présenter des morphologies très différentes de celles de leur ancêtre commun. De plus, les relations phylogénétiques entre groupes fossiles ne sont parfois pas résolues et l’on ne connaît pas le groupe frère des Angiospermes parmi les Spermatophytes fossiles, ce qui empêche la reconstitution du portrait-robot de l’ancêtre commun.
Une approche alternative consiste à étudier les états de caractères présents chez les espèces actuelles. Si on connaît les relations phylogénétiques entre plusieurs espèces (grâce par exemple à des données moléculaires), il est possible de retracer l’évolution de caractères morphologiques sur l’arbre phylogénétique en utilisant le principe de parcimonie. Cela permet de déduire l’état des caractères chez l’ancêtre commun à tout le taxon. Par exemple, au sein du groupe des Primates, toutes les espèces ont un pouce opposable et des ongles plats (contrairement aux autres Mammifères dont ils sont proches). On peut donc en déduire que ces caractères étaient présents chez l’ancêtre commun à tous les Primates. Dans le cas des Angiospermes le problème est plus délicat : les fleurs des espèces ayant divergé le plus précocement présentent de grandes variations morphologiques, ce qui complique la reconstruction. Par exemple Amborella trichopoda (l’unique représentant des Amborellales, groupe frère de toutes les autres plantes à fleurs) possède des fleurs unisexuées, tandis que les Nymphaeales ont majoritairement des fleurs bisexuées : la parcimonie ne peut donner autre chose qu'une reconstruction ambiguë pour la base de l'arbre (au niveau des pièces fertiles des fleurs). Ce trait n'est pas le seul pour lequel la reconstruction en parcimonie est équivoque. Le problème ici concerne à la fois la variation de la morphologie florale à la base de l'arbre, et les limites de la méthode de parcimonie.
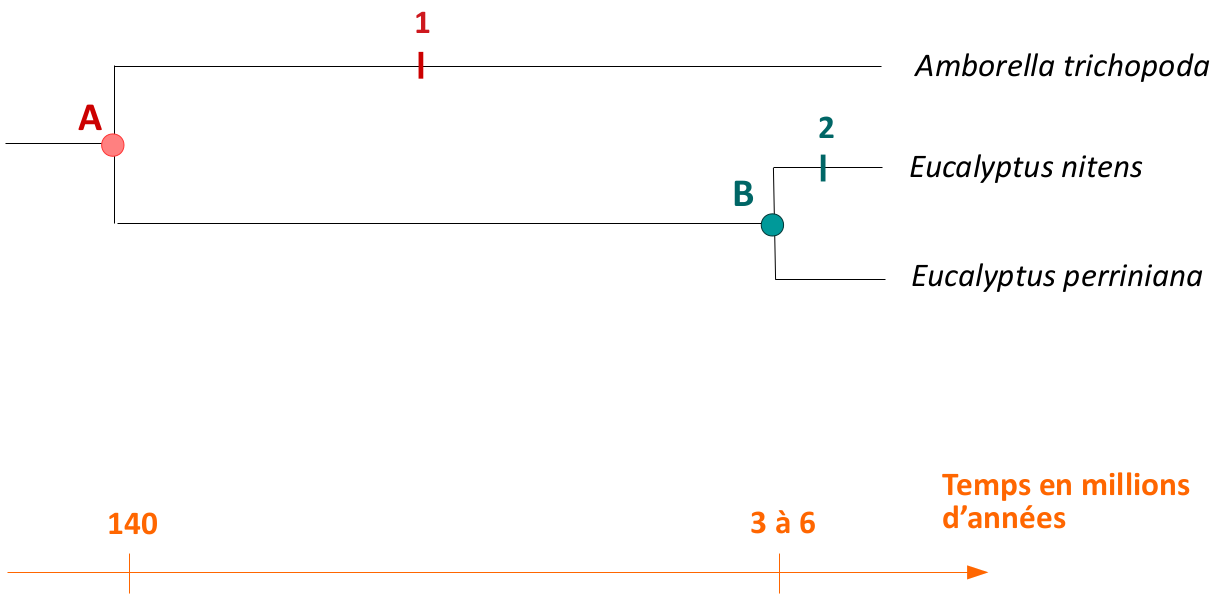
L’ancêtre commun des Angiospermes est noté A, l’ancêtre commun des deux espèces d’eucalyptus est noté B. La transformation sur la branche menant de l’ancêtre A à Amborella trichopoda est notée 1, la transformation menant de l’ancêtre B à Eucalyptus nitens est notée 2. L’âge correspondant à l’ancêtre A est un âge minimal.
Pour répondre à ces difficultés, il est possible d'utiliser des méthodes qui prennent en compte les variations de longueur entre les différentes branches de l'arbre, à savoir les méthodes probabilistes. Le principe est de considérer que la modification d’un état de caractère sur une branche de l’arbre est d’autant plus probable que cette branche est longue (voir Fig.7). Par exemple, la branche menant de l’ancêtre commun des Angiospermes (noté A sur la figure 7) à Amborella est longue d’au moins 140 millions d’années (Magallón et al., 2015) alors que celle menant d’Eucalyptus nitens à son dernier ancêtre commun avec une espèce proche, E. perriniana (ancêtre noté B sur la figure 7), est longue de 3 à 6 millions d’années seulement (Thornhill et al., 2015). La transformation 1 est plus probable que la transformation 2 (le temps écoulé étant plus long), autrement dit il est donc beaucoup plus probable qu’Amborella présente des traits différents de ceux de l’ancêtre A qu’Eucalyptus nitens présente des caractères différents de l’ancêtre B (Fig.7). En parcimonie, les deux modifications ont la même probabilité, tandis que les méthodes probabilistes permettent de considérer que la transformation 1 est plus probable que la transformation 2.
L'étude présentée ici (Sauquet et al., 2017) utilise ce type de méthode probabiliste dans le but de fournir un portrait robot de la fleur de l'ancêtre commun des Angiospermes actuelles. Pour cette étude un arbre de 792 espèces a été utilisé, celles-ci étant choisies de manière à représenter un bon échantillonnage de la diversité des Angiospermes (98% des ordres sont présents, ainsi que 86% des familles). De nombreux caractères concernant les fleurs ont été renseignés, ce qui a permis de proposer des reconstructions de la fleur à différents nœuds, et notamment au niveau de l'ancêtre commun à toutes les Angiospermes.
Les résultats présentés ici correspondent aux résultats obtenus avec les méthodes probabilistes ; ils sont en accord avec les résultats obtenus en parcimonie, mais présentent moins de cas où la reconstruction est ambiguë.
La reconstitution de la fleur de l’ancêtre commun des Angiospermes

Il est essentiel de noter que certains caractères (comme la couleur, la forme ou la taille relative des pièces) n’ont pas été reconstruits, et sont ici représentés de façon arbitraire.
La reconstruction indique que la fleur de l'ancêtre commun des Angiospermes actuelles était bisexuée, présentait une symétrie radiale (fleur actinomorphe), avec vraisemblablement au moins 4 verticilles de 3 tépales pour le périanthe. Les étamines (au moins une dizaine) étaient disposées en plusieurs verticilles (au moins 4 verticilles de 3 étamines, soit la même organisation que le périanthe). Le gynécée était composé de plusieurs carpelles libres (Fig. 8 et 10). La figure 8 ci-dessous montre une représentation possible de cette fleur.
La combinaison de caractères au niveau de la fleur de l'ancêtre commun des Angiospermes est unique, elle ne se retrouve ni chez les fossiles, ni chez les espèces actuelles.
L’évolution du sexe

Amborella trichopoda, unique représentante du groupe des Amborellales, est une plante dioïque : ses fleurs sont unisexuées et les fleurs mâles et femelles sont portées par des plants différents. Les fleurs mâles comportent 9 à 11 tépales verts à jaunes et 11 à 22 étamines organisées en spirale. Les fleurs femelles présentent de 7 à 8 tépales verts à jaunes, 1 à 2 étamines stériles et un gynécée formé par 5 carpelles séparés. La fleur d’Amborella trichopoda est donc très différente de celle de l’ancêtre commun des Angiospermes (figure 8).
Si la parcimonie ne permettait pas de trancher sur la question des pièces fertiles de cette fleur, les reconstructions basées sur des modèles probabilistes d’évolution indiquent que la fleur de l’ancêtre commun des Angiospermes était très probablement bisexuée, avec des étamines et des carpelles au sein de la même structure. Ce résultat est intéressant du point de vue botanique car la question divise depuis longtemps les spécialistes, mais il illustre également un point intéressant en biologie évolutive : Amborella est certes le groupe frère de toutes les Angiospermes actuelles, mais ce n’est en aucun cas l’image de l’ancêtre commun des plantes à fleurs (Figure 9).
La lignée conduisant à Amborella s’est séparée du reste des Angiospermes il y a environ 140 millions d’années, ce qui permet d’envisager un certain nombre de transformations au niveau des caractères morphologiques dans cette lignée. Les groupes taxonomiques divergeant à la base des arbres phylogénétiques sont souvent mal interprétés comme « proches » de l’ancêtre commun : ils ne partagent pas les caractères dérivés apparus plus tard dans la phylogénie, ce qui est aussi le cas de l’ancêtre commun, mais ils peuvent posséder un certain nombre de caractères dérivés qui leur sont propres (autapomorphies) et qui les différencient de l’ancêtre commun. Amborella, et de manière générale les espèces situées à l’extrémité des branches correspondant aux nœuds profonds de l'arbre comme les nénuphars ou les magnolias ont ainsi un certain nombre de caractères dérivés qui les différencient de l’ancêtre commun des Angiospermes, les fleurs unisexuées étant un exemple chez Amborella. De la même manière les amphibiens actuels ne sont pas une image fidèle de l’ancêtre commun des Tétrapodes, ils ont certes en commun avec cet ancêtre un certain nombre de traits ancestraux pour les Amniotes (peau nue, reproduction en milieu aquatique…) mais les amphibiens actuels ont aussi un certain nombre de caractères dérivés que ne possédait pas l’ancêtre commun des Tétrapodes (dents particulières avec un pédicelle par exemple).
Nous avons vu précédemment que la fleur de l’ancêtre commun des Angiospermes, telle que reconstruite ici, ne correspondait à aucune espèce actuelle ou fossile. Cette morphologie a donc évolué, et les fleurs de quelques nœuds profonds ont également été reconstituées, avec la même approche. Les résultats obtenus indiquent que les différentes trajectoires évolutives initiales correspondent toutes à une simplification (réduction du nombre d’organes) par rapport à la fleur de l’ancêtre commun (Fig. 10). L’évolution est souvent vue comme un phénomène entraînant une complexification croissante des organismes, mais nous avons de nombreux exemples illustrant le cas contraire chez les êtres vivants.
La fleur de l’ancêtre des Monocotylédones
Par rapport à la fleur ancestrale, la plupart des Angiospermes présentent une simplification des verticilles. Chez les Monocotylédones par exemple, deux verticilles du périanthe et probablement deux verticilles d’étamines ont été perdus (Fig. 10). La fleur reconstruite pour l’ancêtre commun des Monocotylédones correspond à une fleur trimère, dotée de deux verticilles de trois tépales, deux verticilles de trois étamines, et trois carpelles soudés (Fig. 10). Cette morphologie est très commune chez les Monocotylédones actuelles, et de nombreuses espèces présentent cette organisation, d’autres morphologies pouvant facilement dériver de celle-ci (avec par exemple des pertes ou l’acquisition de la symétrie bilatérale comme chez les Orchidaceae).
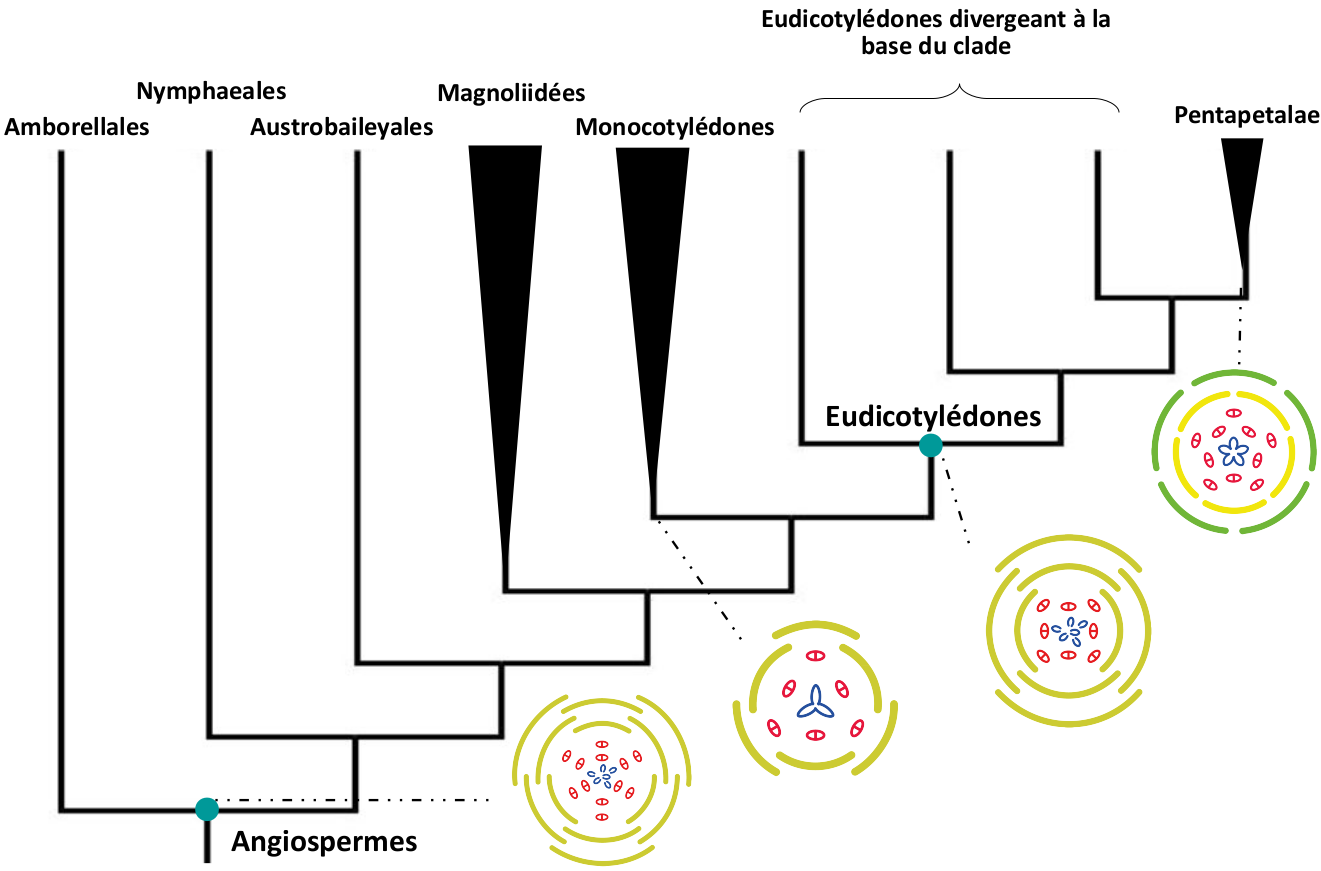
Un périanthe jaune moutarde représente des tépales, tandis que pour les Pentapetalae les sépales sont en vert et les pétales en jaune. Les étamines sont représentées en rouge et les carpelles en bleu.
La fleur de l’ancêtre des Eudicotylédones
La fleur de l’ancêtre commun des Eudicotylédones est plus complexe à reconstituer car les résultats varient entre les méthodes. La méthode la plus fiable nous montre une fleur encore assez complexe, possédant de nombreux points communs avec la fleur de l’ancêtre des Angiospermes, mais ayant perdu des pièces florales dans les verticilles du périanthe et de l’androcée (Fig. 10). Cette fleur était vraisemblablement dimère (même si une fleur trimère est envisageable, dans certaines reconstructions), avec quatre verticilles de deux tépales, probablement deux verticilles d’étamines, et quatre à six carpelles libres (Fig. 10). L’incertitude rencontrée ici est liée à la très grande variabilité au niveau de la morphologie florale que l’on trouve dans les ordres qui divergent à la base du clade des Eudicotylédones, qui complique le travail de reconstruction.
Chez les Eudicotylédones l’organisation de la fleur (nombre de verticilles et nombre de pièces par verticille) devient moins variable dans le grand clade des Pentapetalae. Si la fusion des pièces et la symétrie des fleurs restent variables, les caractères énoncés ci-dessous sont beaucoup plus constants que pour les groupes divergeant à la base du clade des Eudicotylédones. Les Pentapetalae sont caractérisées par une fleur pentamère, comme leur nom l’indique, mais également par une différenciation du périanthe en sépales et pétales. La fleur reconstruite ici possède deux verticilles de 5 étamines libres, et 4 ou 5 carpelles soudés (Fig. 10). Cette morphologie est assez typique des Pentapetalae, même si de nombreuses variations ultérieures ont produit des morphologies florales plus spécialisées (corolle bilabiée des Lamiaceae, cyathe des Euphorbiaceae, corolle papilionacée des Fabaceae : voir Fig. 11 ci-dessous).

Corolle papilionacée des Fabaceae (ici Lathyrus heterophyllus), corolle bilabiée présente chez un certain nombre de Lamiales, dont les Lamiaceae (ici Glechoma hederacea), cyathe du genre Euphorbia (ici Euphorbia amygdaloides) ; la cyathe est généralement interprétée comme une inflorescence.
Photo Lathyrus : HermannSchachner, CC0
Photo Glechoma : AnRo0002, CC0
Photo Euphorbia : Dinkum, CC0
Comment expliquer la simplification de la morphologie florale des Angiospermes ? Cette question est bien trop large pour fournir une réponse détaillée ici, mais on peut penser que l’ancêtre commun des Angiospermes est lui-même apparenté à des végétaux dotés de structures reproductrices potentiellement complexes. Il est possible que la diminution progressive du nombre d’organes ait été avantageuse dans le passé, et le soit encore actuellement. Il serait intéressant pour cela d’avoir des données sur la pollinisation des espèces fossiles, étant donné que les caractères floraux peuvent présenter un avantage significatif au niveau de la pollinisation. Ce type d’information est malheureusement difficilement accessible. La morphologie florale pourrait être impliquée dans la diversification de certaines familles : le capitule des Asteraceae et la corolle bilabiée des Lamiaceae par exemple, pourraient constituer des innovations clés à l’origine de la grande diversification de ces deux taxons. Il est possible qu’un tel scénario impliquant des caractères particulièrement favorables se soit également produit plus précocement dans l’histoire des Angiospermes.
En conclusion cette étude a permis de proposer une reconstruction pour un certain nombre de caractéristiques de la fleur de l’ancêtre commun des Angiospermes actuelles (apparu il y a 140 à 250 millions d’années). Cette reconstitution n’est probablement pas l’image exacte de cette fleur car des incertitudes subsistent, mais ce travail permet d’enrichir les discussions sur l’évolution précoce des Angiospermes en apportant des résultats nouveaux, notamment sur le sexe, le nombre et la disposition des pièces florales. Les méthodes probabilistes ont permis de faire de nouvelles propositions sur des points où la parcimonie était insuffisante, et les études futures basées sur ce type d’approche permettront d’enrichir et de compléter le modèle développé ici.
Les Angiospermes représentent 90 % des espèces de plantes actuelles, et il est probable que leur mode de reproduction, impliquant la structure particulière qu’est la fleur, ait joué un rôle dans le succès évolutif du groupe. Une étude plus approfondie du registre fossile et une meilleure intégration des données pourrait permettre d’une part de mieux comprendre comment est apparue la fleur, et d'autre part, d’étudier la diversification et l’expansion d’un groupe qui a changé de manière considérable la composition des écosystèmes terrestres.
Références
Article original
Sauquet, H., von Balthazar, M., Magallón, S., Doyle, J. A., Endress, P. K., Bailes, E. J., Barroso de Morais E., Bull-Hereñu, K., Carrive, L., Chartier, M., Chomicki, G., Coiro, M., Cornette, R., El Ottra, J. H. L., Epicoco, C., Foster, C. S. P., Jabbour, F., Haevermans, A., Haevermans, T., Hernández, R., Little, S. A., Löfstrand, S., Luna, J. A., Massoni, J., Nadot, S., Pamperl, S., Prieu, C., Reyes, E., Dos Santos, P., Schoonderwoerd, K. M., Sontag, S., Soulebeau, A., Staedler, Y., Tschan, G. F., Wing-Sze Leung, A., & Schönenberger, J. 2017. The ancestral flower of angiosperms and its early diversification. Nature Commun. 8:16047. doi: 10.1038/ncomms16047
Références générales
Herendeen, P. S. [et al. 2017], Friis, E. M., Pedersen, K. R., & Crane, P. R. 2017. Palaeobotanical redux: Revisiting the age of the angiosperms. Nature Plants 3:17015. doi:10.1038/nplants.2017.15
Magallón, S. [et al. 2015], Gómez-Acevedo, S., Sánchez-Reyes, L. L., & Hernández-Hernández, T. 2015. A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 207: 437-453. doi: 10.1111/nph.13264
Sauquet, H. and Magallón, S. (2018), Key questions and challenges in angiosperm macroevolution. New Phytol. doi: 10.1111/nph.15104
Thornhill, A. H. [et al. 2015], Ho, S. Y. W., Külheim, C., & Crisp, M. D. 2015. Interpreting the modern distribution of Myrtaceae using a dated molecular phylogeny. Molec. Phyl. Evol. 93: 29-43.
Classifications APG
Angiosperm Phylogeny Group [= A.P.G.]. 1998. An ordinal classification for the families of flowering plants. Ann. Missouri Bot. Gard. 85: 531-553.
Angiosperm Phylogeny Group [= A.P.G.] II. 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linnean Soc. 141: 399-436.
Angiosperm Phylogeny Group [= A.P.G.] III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linnean Soc. 161: 105-121.
Angiosperm Phylogeny Group [= A.P.G.] IV. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linnean Soc. 181: 1-20.
Logiciels
Arbres phylogénétiques : FigTree - Tree Figure Drawing Tool - Version 1.4.2 - 2006-2014, Andrew Rambaut - Institute of Evolutionary Biology, University of Edinburgh. - http://tree.bio.ed.ac.uk/



