Polymorphisme du gène codant la calpaïne 10 et susceptibilité de développer un diabète de type 2.
Introduction
Les protéines de la famille des calpaïnes (abréviation CAPN) sont des protéases, enzymes capables de digérer des protéines. L’un des membres de cette famille multigénique semble lié au diabète de type 2 : il s’agit de la calpaïne 10 (CAPN-10). Ce n’est pas la première fois qu’une protéase est reliée au diabète : parmi les gènes soupçonnés de prédisposition au diabète, on retrouve déjà deux autres protéases (la carboxypeptidase E et la pro-hormone convertase 1).
Le gène capn-10 est un gène complexe, formé de 13 exons et 15 introns, porté sur le bras long du chromosome 2. Ce gène s’exprime, par épissage alternatif, en 8 protéines différentes (CAPN-10a, b, c, d, e, f, g, et h). Ce grand nombre d’isoformes, ainsi que l’expression ubiquitaire (c’est-à-dire dans tous les tissus) de CAPN-10, suggère un grand nombre de rôles cellulaires différents pour les produits de ce gène.
Le polymorphisme de capn-10
L’étude d’individus diabétiques et non diabétiques, pris dans une population de Mexicains et d’Américains, a montré l’existence de polymorphismes au niveau du gène capn-10, corrélés à l’apparition d’un diabète de type 2. De manière surprenante, ce polymorphisme se situe au niveau de parties non traduites du gène : des introns. Un polymorphisme important a en particulier été relevé au niveau de trois emplacements :
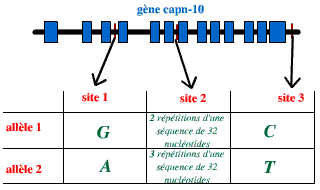 |
| Trois emplacements sont particulièrement polymorphes dans cette population. Pour chaque emplacement polymorphe, on note l'existence de deux allèles prédominants. |
Le risque de développer un diabète est 3 fois plus important chez les individus possédant une combinaison particulière d’allèles, par rapport aux autres individus étudiés. Cette combinaison est la suivante :
| Sur... |
Site 1 |
Site 2 |
Site 3 |
| l'un des deux chromosomes 2 |
allèle 1 (G) |
allèle 1 (2 répétitions) |
allèle 2 (T) |
| l'autre chromosome 2 |
allèle 1 (G) |
allèle 2 (3 répétitions) |
allèle 1 (C) |
Plus précisément, la présence d’un G au niveau du site 1 semble fortement liée à l’apparition d’un diabète de type 2. Ainsi, chez les individus homozygotes G//G, on note les effets suivants :
- baisse de la quantité d’ARN messager de capn-10 dans les cellules musculaires squelettiques ;
- plus grand degré de résistance à l’insuline.
De même, l’inhibition des protéines CAPN dans les cellules musculaires et les adipocytes conduit à une baisse de l’incorporation de glucose par ces cellules, et à une diminution de la synthèse de glycogène musculaire.
Conclusion : hypothèses pour capn-10
Le risque majeur d’apparition d’un diabète en liaison avec la calpaïne 10 est rencontré pour des combinaisons de mutations affectant l’expression de ce gène dans les tissus cibles de l’insuline et les tissus sécréteurs d’insuline. Ces combinaisons pourraient conduire à une résistance accrue à l’insuline, conduisant à une hyperglycémie, ce qui correspond au phénotype des diabétiques de type 2. L’homozygotie G//G au niveau du site 1 semble un facteur de prédisposition particulièrement important.
Attention toutefois ! Le diabète est une maladie avec un déterminisme génétique extrêmement complexe. Ainsi, si les individus G//G semblent bien prédisposés au diabète dans la population étudiée (mexico-américaine), ainsi que dans des populations finlandaises et allemandes, cela ne semble pas être le cas dans d’autres populations (anglaise, samoanes). Il s’agit bien là d’une prédisposition génétique : on peut supposer que l’interaction avec d’autres gènes est nécessaire, et encore à étudier…
Rappel : il s’agit d’un gène de prédisposition. Tous les individus G//G ne sont pas diabétiques.
Précisions : pour les puristes, voici les noms des trois sites de polymorphisme (dans l’ordre) : UCSNP-43, UCSNP-19 et UCSNP-63. Les explications de cette page sont pour l’essentiel rédigées d’après l’article de Y. Horikawa et al., « Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus », Nature genetics (2000) 26 : 163-175. Cette page s’appuie sur les connaissances publiées à la date du 10 décembre 2001 ; des nouvelles données peuvent avoir été publiées depuis.